
续随子ElPDAT1基因生物信息学及表达特性分析
2019-06-01 02:23任国鹏葛丽萍孙超超
山西农业科学 2019年5期
任国鹏,葛丽萍,孙超超,唐 露
(山西农业大学林学院,山西 太谷 030801)
续随子(Euphorbia lathyris L.)属于大戟科大戟属1~2年生草本植物[1],原产于欧洲[2],引入我国栽培已久。续随子在我国主要被用作药用植物[3],作为油料作物却不多见[4]。CALVIN 等[5]研究发现,续随子种子油脂与理想柴油替代品的分子组成类似,可以代替石油作为新能源,是一种具有开发前景的新型能源油料作物,因此,其也被称为“石油植物”[6]。近年来,中国农科院油料作物研究所、中科院植物研究所等科研部门分别进行了续随子种质资源的收集、核型分析、新品种选育及生物柴油制备的研究[7-9]。另外,续随子种子中油脂含量很高,总油脂含量约45%~60%[10]。危文亮等[11]通过对不同的续随子品种的研究发现,续随子种子中油的脂肪酸成分以 C16和 C18脂肪酸为主,主要为棕榈酸(16∶0)、油酸(18∶1)、亚油酸(18∶2)、亚麻酸(18∶3),该 4 种脂肪酸占总脂肪酸含量的96%以上,且在不同试验材料中的油酸占总脂肪酸含量的比例均超过83%。因此,了解续随子在我国作为能源植物发展的潜力,分析其种子的油脂合成过程以及研究调控该油脂合成过程中相关基因具有重要的意义。
油料植物通常在种子中会积累大量的三酰甘油(TAG),这些油脂既是工业用油和食用油的主要来源,也是种子生长发育所必须的能量来源[12]。TAG是油脂的主要储存形式,在内质网上[13]经过多种酶催化甘油和脂肪酸而形成[14]。经二酰甘油酰基转移酶(DGAT)催化二酰甘油而形成三酰甘油的过程被称为Kennedy 途径[15],这是TAG 合成的主要途径。除此之外,TAG 合成还存在着另一条途径,不依赖于脂酰-CoA 的合成途径(即PDAT 途径)。在PDAT 酶的催化下,磷脂酰胆碱(PC)sn-2 位的酰基链能够转移到二酰甘油上,最终形成TAG 和溶血卵磷脂(LPC)[16-17]。这2 条途径是植物种子油脂累积的重要途径,所以,PDAT 在TAG 合成过程中也起着重要的调控作用。PDAT 途径最早在酿酒酵母中发现,且证明PDAT 基因促进了酵母中TAG 的合成[18]。当酵母DGAT 基因发生突变时,该酵母TAG含量与野生型相比降低了50%,而DGAT 和PDAT双突变的酵母中TAG 的含量仅为野生型的1%[19]。以酵母PDAT 基因为探针在拟南芥中获得2 个PDAT 基因,并通过研究证实了该基因在油脂合成中的重要作用[20]。目前,对于续随子ElPDAT1 基因的功能和表达特性的相关研究还未见报道。
本研究以续随子为试材,对PDAT1 基因进行了生物信息学分析,并通过qRT-PCR 分析该基因的表达特性,旨在了解PDAT1 基因在续随子种子油脂合成中的调控机制,为进一步分析该基因的生物学功能及其他油料作物的油脂改良与利用提供参考。
1 材料和方法
1.1 试验材料
试验所用的续随子材料栽培于山西农业大学林学院苗圃,所用种子采自榆次栽培植株。分别采集续随子根、茎、叶、花及种子发育的3 个时期S1(花后 15 d),S2(花后 30 d),S3(花后 45 d)的种子保存于-80 ℃冰箱,用于RNA 的提取。
1.2 ElPDAT1蛋白的鉴定
以拟南芥的AtPDAT1 蛋白为检索序列,对续随子的转录组数据进行BLAST 比对,分析获得ElPDAT1 蛋白,并从转录组数据库中筛选出功能注释为PDAT1 的基因,最终得到续随子ElPDAT1 基因cDNA 全长序列。
1.3 续随子ElPDAT1基因的序列特征及蛋白功能分析
续随子ElPDAT1 蛋白的分子量、脂质系数、平均亲水性指数等理化性质通过在线软件ProParam(https://web.expasy.org/protparam/)进行分析;采用软件 TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)和 SignalP 4.1(http://www.cbs.dtu.dk/services/SignalP/)对续随子ElPDAT1 的跨膜结构域及信号肽进行预测;利用NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测续随子ElPDAT1 蛋白的功能结构域;通过在线软件SOPMA(https://npsaprabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和 SWISS-MODEL(https://swissmodel.expasy.org/)对续随子ElPDAT1 蛋白进行二级结构及三级结构的预测和建模;运用MEGA 7.0 多序列比对软件对续随子和已知功能的植物种类的PDAT1 蛋白序列构建系统进化树。
1.4 续随子ElPDAT1基因的表达特性分析
根据分析获得的续随子ElPDAT1 基因的cDNA 序列,用在线软件Primer 5 设计实时荧光定量PCR 的引物。以ElActin[21]基因作为内参,进行PCR扩增。引物序列如表1所示。
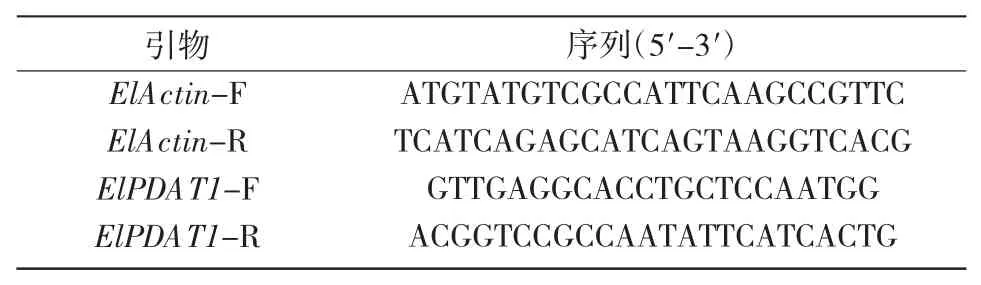
表1 ElActin 和ElPDAT1 基因的荧光定量PCR 引物序列
利用全式金TRNzol RNA 提取试剂提取续随子的根、茎、叶、花及种子不同发育时期的RNA。采用abm 公司的 5×All-In-One MasterMix(with AccuRT Genomic DNA Removal Kit)反转录试剂盒反转录获得cDNA。实时荧光定量PCR 通过CFX96TMOptics Module(BIO-RAD)qRT-PCR 分析仪进行扩增。使用SYBR/FAM only 作为荧光染料,用abm 公司的EvaGreen2×qPCR 试剂盒,以续随子ElActin 作为内参基因,进行实时荧光定量PCR 检测。每个样品分别进行3 次生物学重复和3 次技术重复。反应体系(10 μL) 为:EvaGreen2×qPCR MasterMix 5 μL,Forward Primer 0.3 μL,Reverse Primer 0.3 μL,Template DNA 0.5 μL,Nuclease-free H2O 3.9 μL。实时荧光定量 PCR 反应程序为:95 ℃ 10 min,95 ℃ 15 s,60 ℃ 1 min,循环数为 40 个。
1.5 数据分析
试验采用Excel 和SPSS 软件处理数据,并绘制表达量柱状图。
2 结果与分析
2.1 续随子ElPDAT1基因的序列特征分析
续随子ElPDAT1 基因的cDNA 全长2 968 bp,编码区长度2 037 bp,共编码678 个氨基酸(图1)。对PDAT1 蛋白理化性质分析(表2)表明,该蛋白相对分子质量为75.38 ku,为不稳定亲水性蛋白。

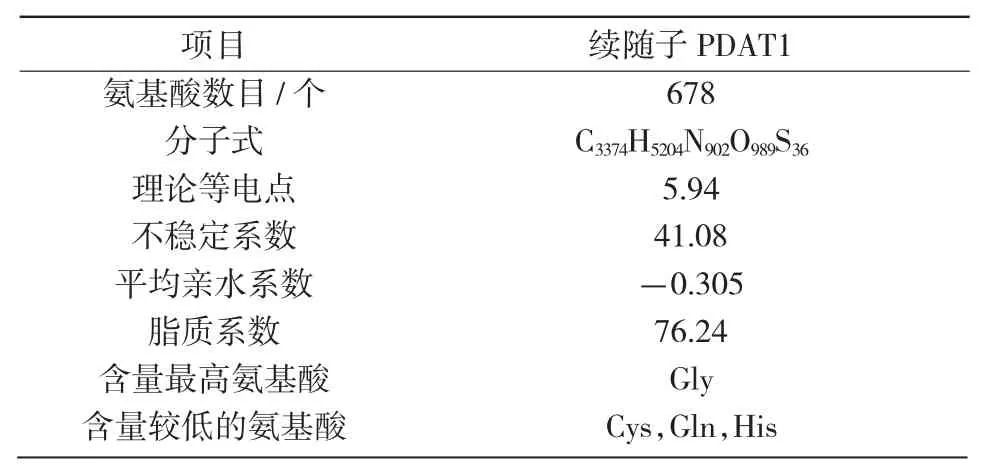
表2 续随子ElPDAT1 蛋白理化性质分析
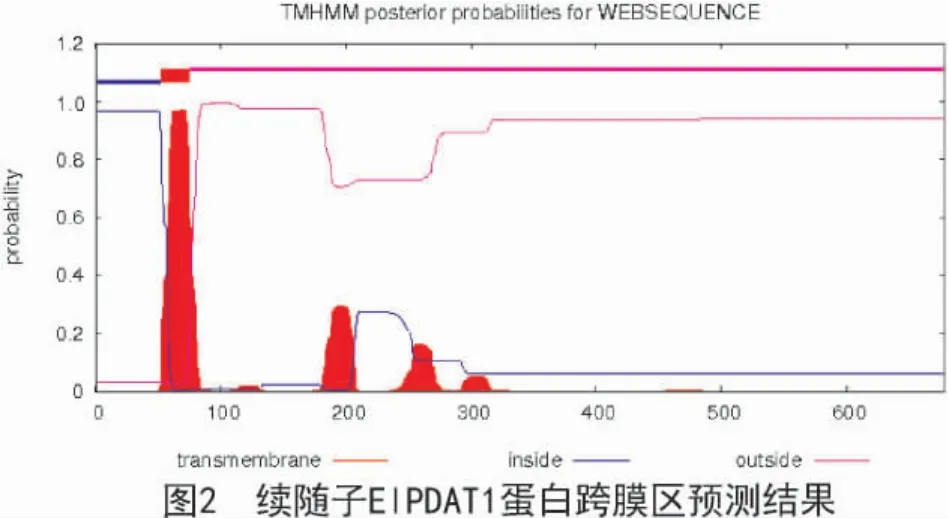
使用TMHMM 在线软件预测蛋白质跨膜区和跨膜方向(图2),续随子ElPDAT1 蛋白存在一个典型的跨膜螺旋区,由膜内到膜外,位于53~75 区域,跨膜现象明显。
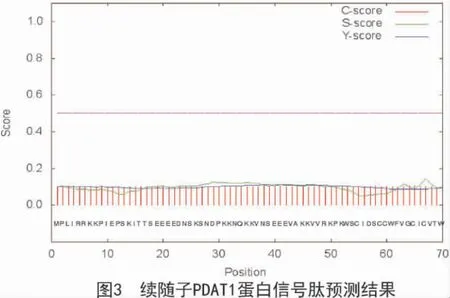
利用Signal P 4.1 软件对续随子ElPDAT1 蛋白信号肽预测结果(图3)表明,该蛋白不存在信号肽及切割位点。
通过NCBI-CDD 对续随子ElPDAT1 蛋白进行功能结构域预测,结果表明,续随子ElPDAT1 基因所编码的蛋白质属于PLN02517 superfamily,含有典型的 PLN02517 保守结构域(图4)。PLN02517 为磷脂酰胆碱酰基转移酶超家族,因此,可以预测ElPDAT1 蛋白具有酰基转移功能,该结果与其他已知的物种PDAT1 蛋白预测的功能一致。

利用在线软件SOPMA 预测续随子ElPDAT1蛋白二级结构,结果表明,ElPDAT1 蛋白的二级结构主要由4 种元件组成:无规则卷曲(43.95%)、α-螺旋(34.81%)、延伸链(14.60%)、β-转角(6.64%)。其中,无规则卷曲和α-螺旋是该蛋白的主要元件。利用SWISS-MODEL 在线软件对续随子ElPDAT1蛋白三级结构预测(图5),与二级结构预测结果一致。

2.2 续随子ElPDAT1蛋白的系统进化树分析

利用MEGA 7.0 多序列比对软件对续随子及蓖麻(Ricinus communis)、橡胶树(Hevea brasiliensis)、麻疯树(Jatropha curcas)、木薯(Manihot esculenta)、陆地棉(Gossypium hirsutum)、花生(Arachis hypogaea)、大豆(Glycine max)等同源性较高物种的PDAT1 蛋白的氨基酸序列进行比对分析,并构建系统进化树。结果发现,续随子与蓖麻、木薯、麻疯树的亲缘关系较近(图6),而它们均属于大戟科,并且它们之间的序列相似度高达84%以上,与陆地棉、大豆等不同科植物的亲缘关系较远。
2.3 续随子ElPDAT1基因在不同器官中的表达特性分析
为了研究ElPDAT1 基因在续随子不同器官及种子发育不同时期的表达量与种子含油量的关系,本研究选取续随子根、茎、叶、花及种子发育的3 个时期S1,S2,S3,对ElPDAT1 基因的表达特性进行分析。结果显示,ElPDAT1 基因在续随子根、茎、叶、花及种子的不同发育时期均有表达,在根和茎中表达量最低,但在种子中的表达量明显高于其他器官。在种子的不同时期,表达量先升高后降低,在种子的S2 时期表达量最高。以叶的表达量作为对照,S2 时期的表达量为叶的5.82 倍(图7)。

3 结论与讨论
植物种子中的油脂主要以TAG 的形式存在,TAG 是通过复杂的生理生化过程在植物体中进行合成和积累。DGAT 是Kennedy 途径中的限速酶和关键酶,对种子的油脂合成起到重要调控作用[22-23]。但是在TAG 不依赖于酰基-CoA 的合成途径中,PDAT 则被认为是TAG 合成的关键酶。因此,为了更好地理解ElPDAT1 在续随子油脂合成中的调控作用,本研究对获得的续随子ElPDAT1 基因进行了详细的生物信息学分析,结果显示,ElPDAT1 基因的cDNA 序列全长为2 968 bp,共编码678 个氨基酸;理化性质预测结果均符合PDAT1 基因的特征,属于不稳定的亲水性蛋白;结构域预测表明,ElPDAT1 蛋白含有PLN02517 保守结构域,并具有PDAT 酶活性。不同物种间进化树分析发现,ElPDAT1 蛋白与蓖麻、木薯、麻疯树的亲缘关系较近,它们的同源性最高达到84%,这与植物分类学中将它们归为一个科的结果一致。
在植物种子中,油脂主要是以TAG 的形式而储存,相对于Kennedy 途径,PDAT 途径的研究相对较少,但有研究表明,将外源PDAT1 基因转入酵母突变体中,使该突变体的TAG 合成能力得到恢复[24-25]。拟南芥中采用RNA 干扰拟南芥AtPDAT1 后,种子中油脂含量减少了70%~80%,且花粉管不能正常萌发,胚发育受阻[26]。其他研究发现,当DGAT1 基因缺失后,拟南芥种子中经过PDAT1 途径合成和积累的TAG 含量高达野生型的70%,且该基因催化产生大量不饱和脂肪酸[27]。可见,PDAT 基因对TAG的合成及植物的生长发育有着不可或缺的作用。因此,为更多地了解续随子中ElPDAT1 介导的TAG合成途径,本试验研究了ElPDAT1 基因在续随子不同器官及种子不同时期的表达量,结果表明,ElPDAT1 基因在续随子的各个器官中均有表达,在根和茎的表达量非常低,但在种子中的表达量比其他器官高出很多。在种子发育初期即S1 时期表达量较高,在发育中期即S2 时期达到最高,而后降低。这与之前对续随子中油脂成分积累规律的研究结果相符,证明ElPDAT1 基因在续随子种子TAG的合成过程中起到关键的作用。
续随子是重要的油料植物,其油脂含量很高,因此,围绕续随子油脂合成关键基因展开研究,是研究续随子的主要途径之一。ElPDAT1 作为续随子种子油脂合成的另一条途径的关键基因,本研究通过生物信息学及qPCR 表达模式分析,证明了该基因与续随子种子油脂合成的关系,为阐明ElPDAT1基因的功能和续随子及其他油料作物油脂改良提供了依据。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
疯狂英语·新读写(2020年3期)2020-06-06
中学生数理化·七年级数学人教版(2019年10期)2019-11-25
中国粮油学报(2019年4期)2019-07-12
中学生数理化·高一版(2018年9期)2018-10-09
湖南教育·C版(2018年3期)2018-06-05
中国粮油学报(2018年12期)2018-01-26
中国老年学杂志(2015年9期)2015-01-31
医学综述(2011年12期)2011-12-09
