
沉默CTTN基因表达抑制非小细胞肺癌A549细胞侵袭的研究
2019-05-23 05:34:32刘迪罗猛通讯作者
医药前沿 2019年11期
刘迪 罗猛 (通讯作者)
(贵州医科大学 贵州 贵阳 550002)
浸润和转移是恶性肿瘤的显著特征,目前尚未发现行之有效的治疗策略来抑制肿瘤的转移,因此世界范围内对肿瘤侵袭转移的研究方兴未艾。最近研究发现,侵袭性伪足(InvadoPodia)与肿瘤的侵袭转移有着密切关系[1]。位于染色体11q13区上的癌基因CTTN所编码的皮层机动蛋白Cortactin (cortical actin bindingProtein)最初被发现是作为致癌的酪氨酸蛋白激酶v-src的底物,与微丝激动蛋白和致癌性酪氨酸激酶v-Src结合,从而影响细胞的侵袭性以及肿瘤转移灶的形成[2]。有研究发现,CTTN基因能够促进食管癌细胞系的运动和侵袭[3]。非小细胞肺癌(Non-Small Cell Lung Cancer,NSCLC)是造成肺癌相关死亡的主要原因。本研究旨在通过siRNA基因沉默技术检测CTTN基因表达下调后对NSCLC中具有较强侵袭性的A549细胞株侵袭能力的影响。
1.材料和方法
1.1 细胞培养
肺腺癌A549细胞(美国ATCC细胞库)以含有10%胎牛血清(浙江天杭生物科技股份有限公司)的DMEM高糖培养基(美国Hyclone公司)作为完全培养基,在37℃,5%CO2浓度条件下常规培养。待其处于对数生长期时进行后续试验。
1.2 细胞转染及免疫印迹
将A549细胞铺于六孔板,每孔2ml完全培养基。待细胞融合度达到60%左右时采用liPofectamine2000(美国Invitrogen公司)脂质体转染法分别将阴性siRNA和CTTN siRNA(广州市锐博生物科技有限公司)转染A549细胞(siRNA∶liPo 2000=5μl∶5μl),48h后使用RIPA裂解液(北京索莱宝科技有限公司)裂解细胞提取蛋白,SDS-聚丙烯酰氨凝胶电泳进行蛋白分离。冰水浴中100V,100min转膜。脱脂奶粉封闭1h后,兔抗Cortactin蛋白一抗(稀释比例1∶3500,美国Abcam公司)室温孵育1h,山羊抗兔二抗(稀释比例1∶5000,北京索莱宝科技有限公司)室温孵育1h,加入ECL化学发光液后于美国AlPha公司Fluorchem Q型蛋白成像系统进行曝光,采用Imag J软件分析条带灰度值。
1.3 激光共聚焦显微镜检测
1ml细胞悬液中计数3000个细胞加到直径15mm的共聚焦小皿,待细胞其充分贴壁后,4%多聚甲醛固定,0.2%Triton-±s100破膜,兔抗cortactin-体(稀释比1:1000)室温孵育1h,羊抗兔dylight488荧光二抗(1:2000,美国Abbkine公司)再次室温孵育1h,5μg/ml Tritc-鬼笔环肽(上海翊圣生物科技有限公司)室温避光孵育30min后使用德国ZEISS公司LSM 700型激光共聚焦显微镜随机选取5个视野,统计出现侵袭性伪足的细胞数和总细胞数并计算百分率。
1.4 Transwell小室模型
实验前一天将Matrigel基质胶(美国BD公司)置于4℃冰箱中融化过夜。实验时用预冷的枪头抽取已用无血清培养基稀释至1mg/ml的Matrigel基质胶80μl,铺于孔径为8μm的Transwell小室(美国Corning公司)的膜上。基底膜水化后,从1×106/ml浓度的无血清细胞悬液中抽取200μl加至Transwell上室。24孔板下室加入700µl完全培养基。24h后,弃去孔中培养液,甲醇固定20分钟,吉姆萨染色20min,PBS洗涤2遍后用棉签轻轻擦掉上层未迁移细胞。200倍显微镜下随即五个视野观察细胞,记数。
2.统计学分析
数据采用SPSS20.0统计软件进行统计学分析,计数资料采用率(%)表示,进行χ2检验,计量资料采用(±s)表示,进行t检验,P<0.05为差异具有统计学意义。
3.结果
3.1 siRNA沉默效率检测
如图1所示,western blot检测发现,转染CTTN基因的特异性siRNA(si-CTTN)后Cortactin蛋白表达显著降低,差异具有统计学意义。表明CTTN基因的沉默可以显著抑制Cortactin蛋白的表达。
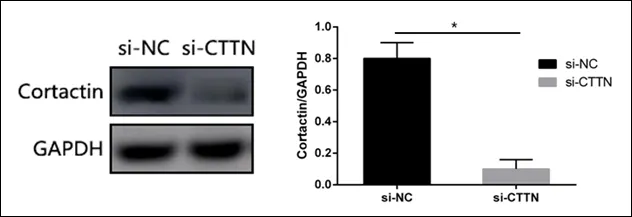
图1 CTTN基因沉默效率检测
3.2 CTTN的沉默抑制A549细胞株侵袭性伪足的形成
共聚焦显微镜下,呈现红色荧光的TRITC-鬼笔环肽标记的F-actin和呈现绿色荧光的Dyligt488标记的cortactin共定位于胞核周围所呈现出的黄色荧光,即为侵袭性伪足。
如图2所示,镜下观察发现,与si-NC组的71.46±8.9%侵袭性伪足出现率相比,CTTN沉默(si-CTTN)组A549细胞中侵袭性伪足出现率显著降低至22.37±4.8%(P<0.05)。

图2 CTTN基因沉默对A549细胞侵袭性伪足的影响 (bar=10μm)
3.3 CTTN的沉默降低A549细胞株侵袭能力
如图3所示,Transwell细胞侵袭模型中,倒置显微镜观察发现,与si-NC组的220±27个侵袭细胞相比,CTTN的沉默导致侵袭细胞数显著降低至51±10个(P<0.05)。

图3 CTTN基因沉默对A549细胞侵袭能力的影响(×200)
4.讨论
无论是在我国还是在全球范围内,NSCLC都已成为致死率最高的恶性肿瘤[4]。30%早期阶段的NSCLC患者可通过外科手术就能治愈肺癌。然而遗憾的是,NSCLC患者被发现时往往已处于肿瘤晚期,肺癌的远端转移导致患者已无法通过手术切除进行治疗。肿瘤的转移复发也严重影响患者的预后,导致生存期显著缩短[5]。因此,如何抑制恶性肿瘤的转移,从微生物学角度考虑如何抑制肿瘤细胞的运动侵袭,是目前肿瘤学研究的重中之重。
细胞伪足是肿瘤细胞侵袭转移的主要“工具”,尤其是侵袭性伪足在细胞运动过程中扮演着非常重要的角色。肌动蛋白是细胞伪足的主要组成部分,也是导致肿瘤细胞侵袭转移的主要参与蛋白[6]。另有文献报道,活体荧光标记肌动蛋白证实,微丝骨架在侵袭性肿瘤细胞伪足前沿的延展和形态构建中发挥重要作用[7]。CTTN基因编码的皮层蛋白(Cortactin)是一种微丝肌动蛋白结合蛋白,它可以与肌动蛋白微丝的侧面结合,并直接参与细胞骨架的成型[8]。目前的研究已经发现,cortactin是连接细胞骨架重建信号通路中的关键分子,在调控细胞的运动性、侵袭性、以及肿瘤转移灶形成中发挥关键作用[9]。目前已被证实Cortactin广泛分布诸如层状伪足、侵袭性伪足和细胞膜皱褶等细胞外围[10]。同时发现,CTTN基因编码的cortactin参与了众多以肌动蛋白为基础的过程。cortactin在肿瘤分化、侵袭和转移等病理情况下表达异常升高。
在本研究中,我们采用siRNA将CTTN基因进行特异性沉默,并用western blot技术检测发现,Cortactin蛋白的表达被显著抑制,证实了CTTN基因确实编码Cortactin蛋白,并验证了脂质体转染这种方法学的可靠性。在此基础上,我们使用激光共聚焦显微镜和Transwell细胞侵袭小室模型发现,CTTN基因的沉默导致出现侵袭性伪足的细胞数显著少于对照组,并且穿过Matrigel基质胶侵袭至Transwell下室的细胞数显著少于对照组,差异均差异具有统计学意义。但是值得注意的是,本研究只是通过体外验证CTTN基因参与调控细胞运动。动物模型及临床出现肿瘤转移的患者中CTTN基因及其编码的Cortactin蛋白的检测将是对本研究提供更强的说服力和可信度。
猜你喜欢
材料工程(2024年3期)2024-03-29 16:36:08
中学生数理化·八年级物理人教版(2020年11期)2020-12-14 06:58:18
中国畜牧杂志(2020年1期)2020-01-16 04:09:54
小学科学(学生版)(2019年11期)2019-12-09 09:06:30
安徽农学通报(2019年3期)2019-03-18 01:18:06
儿童故事画报·发现号趣味百科(2016年7期)2017-02-08 09:10:14
中国疼痛医学杂志(2017年5期)2017-01-10 14:22:26
中国医科大学学报(2015年10期)2015-03-01 02:09:59
发明与创新(2015年25期)2015-02-27 10:39:26
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:05
