
玉米叶片水分利用效率的保守性
2019-05-13 02:06:48周怀林周广胜
生态学报 2019年6期
周怀林,周广胜
1 南京信息工程大学应用气象学院,南京 2100442 中国气象科学研究院,北京 1000813 南京信息工程大学气象灾害预警协同创新中心,南京 2100444 中国气象局固城农业气象野外科学试验基地,保定 072656
植物的碳同化和水分蒸腾过程通过气孔而紧密耦合[1],一般用水分利用效率(Water Use Efficiency,WUE 即单位水分消耗量所同化的碳量)量化这一耦合关系的强弱[2-3]。水分利用效率是植物个体或生态系统水分利用过程的重要特征参数,可表征不同时空尺度的植物碳-水耦合关系,对植物响应气候变化和气候适应性研究具有重要意义[4- 7]。在叶片尺度上,水分利用效率通常定义为净光合速率与蒸腾速率的比值(Pn/Tr)[8],以表示植物叶片对内、外环境变化的综合响应。由于叶片的水分利用涉及CO2和水分的吸收、转化、运输和利用等一系列生物-物理-化学过程,导致水分利用效率受到众多因素的影响。目前,影响水分利用效率的因素可划分为环境因素和生物因素两类,环境因素可以分为大气环境因素和土壤因素,其中大气环境因素涵盖了CO2浓度、太阳辐射、温度等因素,土壤因素包涵了土壤水分、土壤机械组成、土壤理化性质和土壤微生物等因素;而生物因素主要包括物种类别、光合途径、生育期、叶龄、气孔结构特征等[9- 11]。对影响水分利用效率的环境因素研究发现,CO2浓度升高会导致Pn增强,Tr降低,水分利用效率显著提高[8,12- 14];风速对水分利用效率的影响类似于CO2浓度[15],而温度则相反[16];适度的土壤水分亏缺能提高水分利用效率[17],但土壤水分的有效阈值范围仍不清楚。水分利用效率的生物影响因素研究表明,C4植物的水分利用效率要普遍高于C3植物[18];水分利用效率随着生长进程的推进而升高[19];物种间水分利用效率对气候的适应也存在很大差异[20-21];比叶面积被认为与水分利用效率存在一定的相关性,但随物种及其所处环境而异[22]。值得注意的是,环境因素和生物因素对水分利用效率并不是单独起作用的,它们之间往往存在明显的交互作用。例如,土壤水分降低会刺激新气孔的产生,使得叶片上的气孔密度增加,但气孔的体积和孔径缩小,导致气孔导度降低,并且Tr的下降幅度大于Pn,从而造成水分利用效率升高;同时,水分利用效率的升高反过来会减少根对土壤水分的吸收,进而减缓土壤水分胁迫[9,23- 25]。
已有研究对于理解植物叶片水分利用规律及其特性具有非常重要的意义,这些研究通常是测定植株顶部第1片或者其他完全展开叶来分析植株水分利用特征,目前关于植物不同叶位叶片的水分利用效率及其相互关系仍鲜见报道。本研究试图以玉米作为研究对象,利用2013—2014年玉米不同灌溉方案模拟试验资料,分析玉米顶部第1片完全展开叶水分利用效率在生育期内的变化规律及其与主要生理生态因素的关系,并探讨不同叶位叶片的水分利用效率及其相互关系。为此,本研究提出两个基本假设:(1) 水分利用效率在不同叶位之间存在相对稳定性特征;(2) 不同叶位之间的水分利用效率相对稳定性由Pn和Tr的协调变化决定。
1 材料与方法
1.1 试验站点概况
试验于2013—2014年玉米生长季(6月下旬—10月上旬),在中国气象局固城农业气象野外科学试验基地(115°40′E,39°08′N)的大型可控式水分试验场开展。该站位于华北平原北部高产农业区,气候类型属于暖温带大陆性季风气候,年平均气温为13.6℃,降水量494.0 mm(70%以上集中在夏季)。该站海拔15.2 m,地势平坦,土层深厚,土壤类型以砂壤土为主。土壤有机碳含量约为13.67 g/kg,全氮0.87 g/kg,有效磷25.76 mg/kg,有效钾118.55 mg/kg,pH值8.19[26]。0—50 cm土壤平均田间持水量为23.4%,凋萎系数7.10%,土壤容重1.23 g/cm3。试验场配有电动可移动式遮雨棚,发生降雨时启用,以隔绝自然降水。试验场内每个试验小区面积为8 m2(4 m×2 m),小区之间有3 m深的混凝土隔离墙以防止小区间水分的水平交换。
1.2 试验设计与田间管理
2013—2014年试验玉米品种、播种时间、水分处理的开始时间和水分处理梯度设计等如表1所示。2013年水分处理开始前,各小区适当灌溉,土壤湿度始终保持在适宜水平(约70%田间持水量)。2014年水分控制之前,为了保证出苗率,土壤湿度维持在50%左右的田间持水量。两年试验均采取一次性灌水后不再进行灌溉的水分控制方案,并在全生育期内利用大型电动遮雨棚遮挡自然降水,水分处理梯度依据近30年7月下旬平均降水量(80 mm,2013年)和7月平均降水量(150 mm,2014年)进行设置。试验还设置了一个自然降水对照处理,试验开始后在全生育期内不用遮雨棚遮挡自然降水(表1),且所有处理均设有3个重复。两年试验的施肥量(磷酸二铵)均为240 g/小区。2013年试验小区的玉米行距和行距分别设置为50,30 cm,种植密度为52株/小区,收获时间为10月8号;由于试验取样较为频繁,2014年将株距调整为25 cm,行距保持不变,种植密度为64株/小区,收获时间为10月9号。

表1 试验设计基本信息表
T:2013年水分处理,Water Treatment in 2013; W:2014年水分处理,Water Treatment in 2014
1.3 观测项目与方法
1.3.1土壤相对含水量
试验处理开始后,于每个试验小区内选取1个取样点(两行玉米之间),利用1 m土钻分层取样(10 cm/层),取样深度为50 cm,即0—10、10—20、20—30、30—40、40—50 cm共5层土壤样品。采用烘干称重法测定每层土壤重量含水量W。
W=(m2-m3)/(m3-m1)
式中,W土壤重量含水量(g/g);m1为土壤铝盒重(g);m2为土壤铝盒和湿土的总重量(g);m3为土壤铝盒和干土的总重量(g)。
SRWC=(W/FWC)×100%
式中,SRWC为相对土壤湿度;FWC为田间持水量(g/g)。
1.3.2光合速率和气体交换参数
利用Li- 6400便携式光合系统分析仪(Li-COR Inc.,Lincoln,USA)进行叶片气体交换参数测定。每个水分处理选取3株长势一致、健康的玉米植株进行叶片光合作用的测定:在开花期之前选取第1、3片完全展开叶(从植株顶部数起),开花期之后则选取顶端第1片完全展开片(变态叶除外)、穗位叶和下部老叶(约为第9—10片叶),测定部位为叶片的中上部,且避开中央叶脉。空气流速设为500 μmol/s,CO2浓度、光合有效辐射和温度与外界大气环境一致,观测时段为晴朗天气的9:00—12:00。观测间隔为10—14d,2013进行了5次观测,依次为DAS 33(七叶末期)、DAS 42(拔节期)、DAS 52(拔节-抽雄期)、DAS 59(开花期)、DAS 85(乳熟期);2014年进行了7次观测,分别为DAS 25(七叶—拔节期)、DAS 37(拔节期)、DAS 44(拔节末期)、DAS 56(吐丝期)、DAS 72(灌浆期)、DAS 84(乳熟-成熟期)、DAS 95(成熟期)。记录的参数包括净光合速率Pn(μmol m-2s-1)、Gs(mol m-2s-1)、蒸腾速率Tr(mmol m-2s-1)等。水分利用效率(WUE)由Pn和Tr计算而来,即WUE=Pn/Tr。
1.3.3叶片生物量与叶面积
叶片生物量采用烘干法测定,即将叶片从植株上剪下后迅速进行鲜重称量,然后装入牛皮纸袋中,置于105℃烘箱中杀青1小时,再将烘箱温度调至80℃继续烘干至恒重,最后称量叶片干重。叶面积测量采用面积法测定[27],用直尺量取叶片的长度L(cm)和叶片最宽处的宽度W(cm),然后利用经验公式计算单叶面积:S=0.75×L×W。比叶面积(Specific Leaf Area,SLA)由叶片面积与其干重的比值计算而来。
1.4 数据处理
本研究的数据整理通过Excel 2016完成,统计分析和绘图采用R软件(R 3.4.2,R Core Team,2018)。第1片完全展开叶的生理生态特征与水分利用效率的相关性分析采用常用的Pearson显著性检验方法(psych包)。在玉米开花期之前,对第1片和第3片完全展开叶的两个叶位层次进行比较;而玉米开花期之后则对第1片完全展开叶、穗位叶和下部老叶的3个叶位层次进行比较分析。所以,不同叶位的水分利用效率差异分析分为开花期前和开花期后两个阶段,水分处理、观测时间和叶位对水分利用效率的影响采用三因素(three-way ANOVA)方差分析(car包),并用最小显著法(Duncan′s)进行多重比较。
2 结果分析
2.1 第1片完全展开叶光合生理生态特征对土壤水分的响应2.1.1 气体交换参数和水分利用效率
在2013年试验中,T1处理的土壤水分在玉米全生育期内维持在较高水平(0—50 cm平均相对湿度80%±7.8%),而T2—T6处理在试验处理后持续降低。如图1所示,T1处理的净光合速率(Pn)在DAS 42(拔节期)达到最大值37.21 μmol m-2s-1,此后则随着生育期而降低,至DAS 85(乳熟期)为19.10 μmol m-2s-1,下降了48.67%;而T2—T6处理则在DAS 33(七叶末期)开始逐渐降低,并且呈现出土壤水分胁迫越强下降幅度越大的规律。T1—T4水分处理的蒸腾速率(Tr)整体随着水分处理的延续不断降低,T5—T6则存在明显波动。气孔导度(Gs)的变化趋势与Pn相似,不同的是各处理之间的差异愈加明显。叶片水分利用效率的整体变化趋势在DAS 33至DAS 85期间(七叶末期-乳熟期),表现为先升高后降低再升高,水分较好的处理T1—T3的变化幅度要明显小于水分状况相对较差的T4—T6处理,并且其值也低。
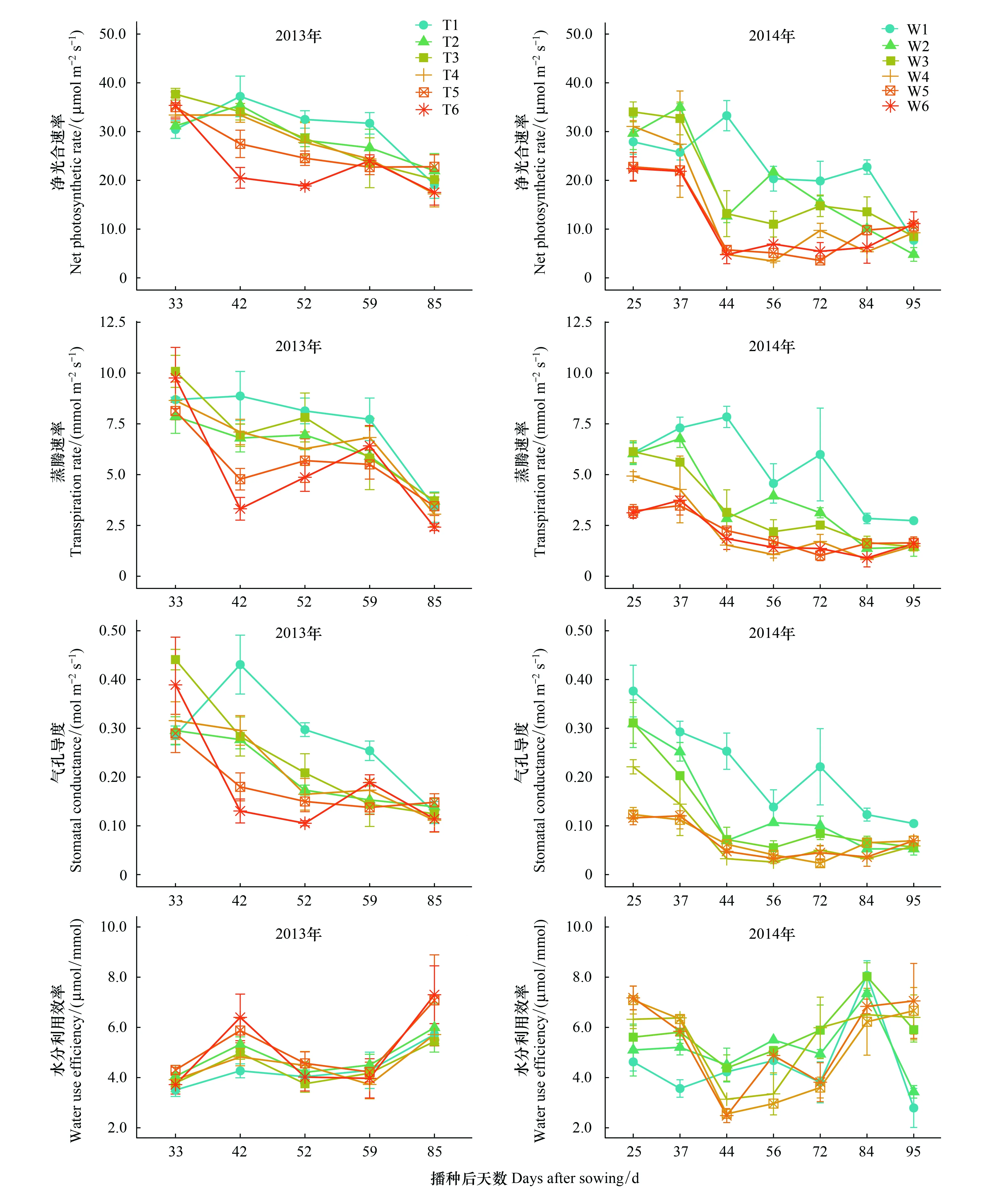
图1 第1片完全展开叶的气体交换参数和水分利用效率的变化趋势Fig.1 The gas exchange parameters and water use efficiency (WUE) for the first fully opened leaf and their changes with timeDAS:播种后天数,Days after sowing; T1—T6为2013试验的6个水分处理,W1—W6为2014试验的6个水分处理; 图中数值平均值±标准误
在2014年试验中,W1为自然降水处理,其Pn在生育期早期有所上升,并在DAS 44(拔节末期)达到最大值,然后随着生育期而降低,其变化趋势与2013年T1处理类似,并且在成熟期达到最小值。W2—W6处理在进入拔节期后急速下降,之后维持在较低水平,并且水分处理高的处理要明显高于水分低的处理(图1)。Tr的变化趋势与Pn相似。不同水分处理的Gs在生育期内则表现出逐渐降低的趋势,所不同的是W1在DAS 72(灌浆期)有一个明显反弹,而W2—W6处理则是先急速下降然后平缓波动。水分利用效率的变化大致呈现出“U”型分布,在生育期早期,低水分处理的水分利用效率大于高水分处理,而在DAS 37(拔节期) 结束后则相反,直到DAS 84(乳熟—成熟期),最后在DAS 95(成熟期)再次发生反转(图1)。
2.1.2生理生态性状
2013年各处理的叶片含水量(Leaf water content,LWC)随着生育期发展而逐渐降低(图2),处理之间的差异在生育期初期和末期差异较大,而在生育期中期相对较小。叶面积(Leaf area,LA)和叶片干物质含量(Leaf dry mass,LDM)的变化趋势相一致,皆在拔节期末期-抽雄初期达到最大值,T1—T6之间的差异比较明显,总体表现为灌水量高的处理大于灌水量低的处理。比叶面积(Specific leaf area,SLA)由叶面积和叶片干物质含量共同决定,表现出持续下降的趋势。2014年试验表明,W1—W3处理的叶片水分含量变化趋势与2013年类似,但是W4—W6处理则是先减少后增加再减少,在DAS 84(乳熟-成熟期)出现一个明显的反弹峰值。叶面积和叶片干物质含量的变化趋势十分相似,呈现出明显的单峰分布特征,并在拔节末期达到最大值。比叶面积呈现出缓慢上升的趋势,且灌水量越多比叶面积越小。

图2 第1片完全展开叶的生理生态特征变化趋势Fig.2 Physiological and ecological characteristics changes of the first fully opened leaf
2.1.3气体交换参数与功能性状的关系
由表2中可知,2013年玉米植株的第1片完全展开叶的水分利用效率与光合参数(Pn、Tr、Gs)和功能性状参数(比叶面积、叶片水分含量)存在显著的负相关(P<0.05),同时Pn与其他光合参数和功能性状参数都具有显著正相关关系。Tr与Pn、Gs极显著正相关,其次是比叶面积和叶片水分含量。叶面积主要与叶片干物质含量相关性最强,其次是叶片水分含量和比叶面积,光合参数中只与Pn的相关性达到显著水平。比叶面积除了与水分利用效率和叶片干物质含量显著负相关外,与Tr、Gs、Pn和叶面积表现出显著正相关关系。
2014年的试验发现,水分利用效率与Pn显著正相关,而与Tr、叶面积和叶片干物质含量则表现出显著负相关关系(P<0.05)。Pn与除比叶面积以外的其他参数的相关性类似与2013年,为显著正相关,但与比叶面积显著负相关。Tr、Gs与其他参数的相关性与Pn大体一致。叶面积与叶片干物质含量的正相关性极强,其次是叶片含水量,而与比叶面积呈现出显著负相关关系。叶片干物质含量与比叶面积的负相关性要强于叶面积,而比叶面积与叶片含水量的正相关性很弱。

表2 第1片完全展开叶生理生态特征与水分利用效率(WUE)的相关分析
WUE:水分利用效率,Water use efficiency;Pn:净光合速率,Net photosynthetic rate;Tr:蒸腾速率,Transpiration rate;Gs:气孔导度,Stomatal conductance; LA:叶面积,Leaf area; LDM:叶干重,Leaf dry mass; SLA:比叶面积,Specific leaf area; LWC:叶片含水率,Leaf water content.* 表示P<0.05,** 表示P<0.001; ***表示P<0.0001
2.2 不同叶位叶片水分利用效率对土壤水分的响应
2013年试验发现,在DAS 59(开花期)之前T1—T6处理的第1片叶和第3片叶的水分利用效率在同一时期相同处理内没有显著差异,且第1片叶的水分利用效率总体要高于第3片叶,而水分处理之间仅在DAS 42(拔节期)观测期间表现出显著差异(P<0.05)。随着水分梯度的减少,水分利用效率表现出增加趋势。在进入DAS 59(开花期)后,相同水分处理不同叶位叶片之间的水分利用效率也无明显差异,同时T1—T6处理的水分利用效率大小无明显差别(图3)。
如图3所示,在2014年试验中除DAS 44(拔节末期)之外,其余时期的水分利用效率皆表现出明显的水分处理差异,但是同时期相同水分处理的不同叶位水分利用效率仍无明显差异。在DAS 72(灌浆期)之前,较高水分处理W1—W3的表现出相对较低的水分利用效率,而较低水分处理W4—W6则相反。在DAS 44(拔节末期)开始,较高水分处理的水分利用效率却表现出与前期不一样的规律,展示出较高的水分利用效率,直到DAS 84(乳熟—成熟期)。在DAS 95(成熟期),高水分处理表现出低水分利用效率,而低水分处理表现出高水分利用效率。从玉米整个生育期来看,叶片水分利用效率在不同生育期存在波动,生育期后期的水分利用效率总体上要高于生育前期。
三因素方差分析表明水分处理对于叶片水分利用效率具有显著影响。2013年水分处理对水分利用效率的影响主要在开花期之前,在开花期以后各水分处理间的水分利用效率无明显差异;而2014年水分处理对整个生育期的水分利用效率将达到或已达到显著水平(P<0.05)(表3)。叶位对水分利用效率的影响仅在2013年开花期之前达到显著水平,说明相同外界环境条件下,玉米植株不同叶位之间的水分利用效率不受其所在的植株部位影响。观测日期(即不同生育期)对水分利用效率的影响在试验期间表现出明显的差异。对水分处理、叶位和生育期三大因素之间的交互作用分析发现,仅水分处理和观测日期的交互作用达到显著水平。
2.3 不同叶位叶片光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)的相对变化特征
在2013年的试验中,DAS 59(抽雄—开花期)之前,第1片完全展开叶的光合速率(Pn)和蒸腾速率(Tr)都要高于第3片完全展开叶,并在DAS 42(拔节期)和DAS 52(拔节—抽雄期)达到显著水平(P<0.05),相对变化率为-37.72% — -9.62%(表4)。在DAS 59之后,玉米已进入生殖生长阶段,穗位叶的Pn和Tr则大于第1片完全展开叶,但未达到显著水平,而植株下部老叶的光合速率和蒸腾速率显著小于第1片完全展开叶(P<0.05)。试验结果表明,在不同叶位叶片光合速率和蒸腾速率呈现出协同变化的特征,同一时期不同叶位的水分利用效率具有保守性。
如表5所示,在2014年的试验中,在DAS 56(抽雄—开花期)之前,第1片完全展开叶的Pn和Tr基本上高于第3片完全展开叶,相对变化率的变化方向一致、变化幅度在-25.51% — -9.44%之间。在DAS 56(抽雄—开花期)之后,穗位叶的Pn和Tr起初大于第1片完全展开叶,随后则一直小于第1片完全展开叶,而植株下部老叶的Pn和Tr则一直显著小于第1片完全展开叶(P<0.05)。叶片水分利用效率在同一时期相对第1片叶的相对变化率在-20.27%—6.77%之间,未达到显著水平,表明水分利用效率具有叶位和叶龄的保守特性。
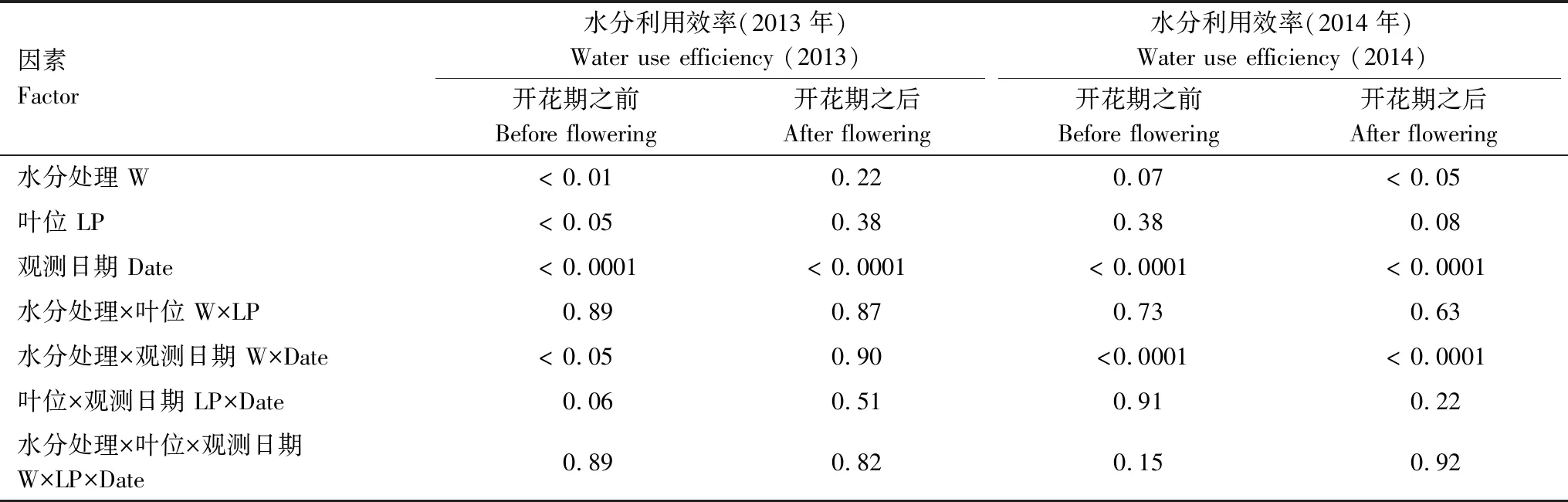
表3 叶片水分利用效率(WUE)的三因素方差分析
W:水分处理,Water treatment; LP:叶位,Leaf positions; 表中数值表示方差显著性检验的概率(P值)
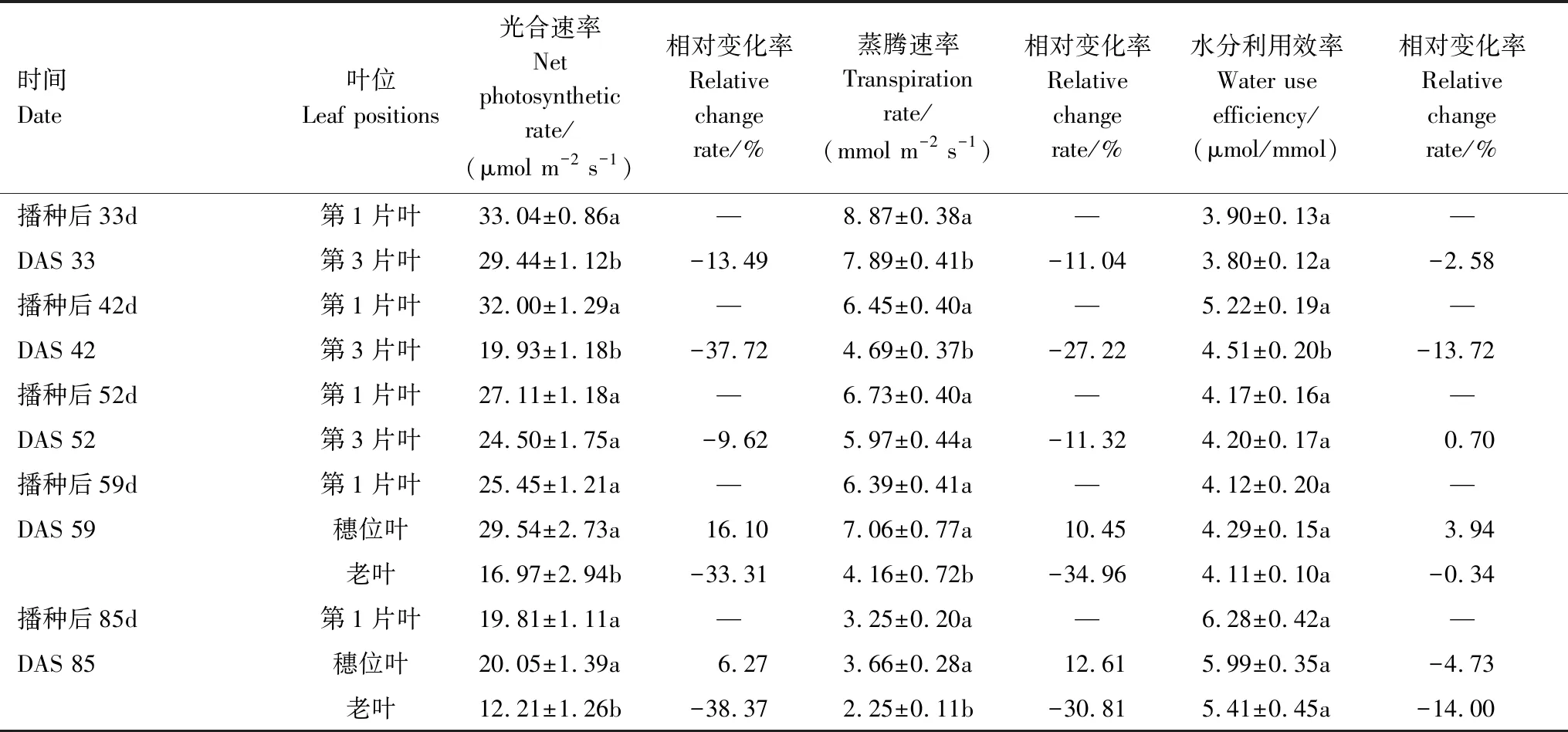
表4 不同叶位光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)的相对变化(2013年)
DAS:播种后天数,Days after sowing; 表中数值为平均值±标准误; 不同小写字母代表差异性显著(P< 0.05); Relative change rate,相对变化率,表示其他叶位的光合速率(或蒸腾速率,水分利用效率)相对第1片完全展开叶的变化百分率
3 讨论
叶片是植物重要的光合器官,也是对环境变化响应非常敏感的器官。植物叶片水分利用效率的动态变化按时间长短可划分为日、季节和年变化。一般来讲,植物叶片水分利用效率在上午时段要明显高于下午时段。研究发现,在水分胁迫条件下玉米叶片水分利用效率日变化均呈现出上午和下午高、中午低的趋势[28],并出现典型的“午休”现象[29]。利用稳定碳同位素对木本植物水分利用效率的季节和年变化研究表明,生长初期一般高于生长后期[30]。对草本植物(玉米)的研究也发现,随着同一叶片叶龄的增加,叶片水分利用效率逐渐降低[31]。然而,相同叶位的叶片水分利用效率在生育期内的变化趋势鲜见报道,特别是在土壤水分持续减少的过程中。本研究表明,植株顶部第1片叶片水分利用效率在拔节期和乳熟期具有两个峰值,存在明显的周期变化规律(图1)。拔节期和乳熟期分别是植株形态生长和籽粒形成的关键时期,此时较高水分利用效率体现了植株旺盛的生长需求,是植物水分利用策略的重要体现。
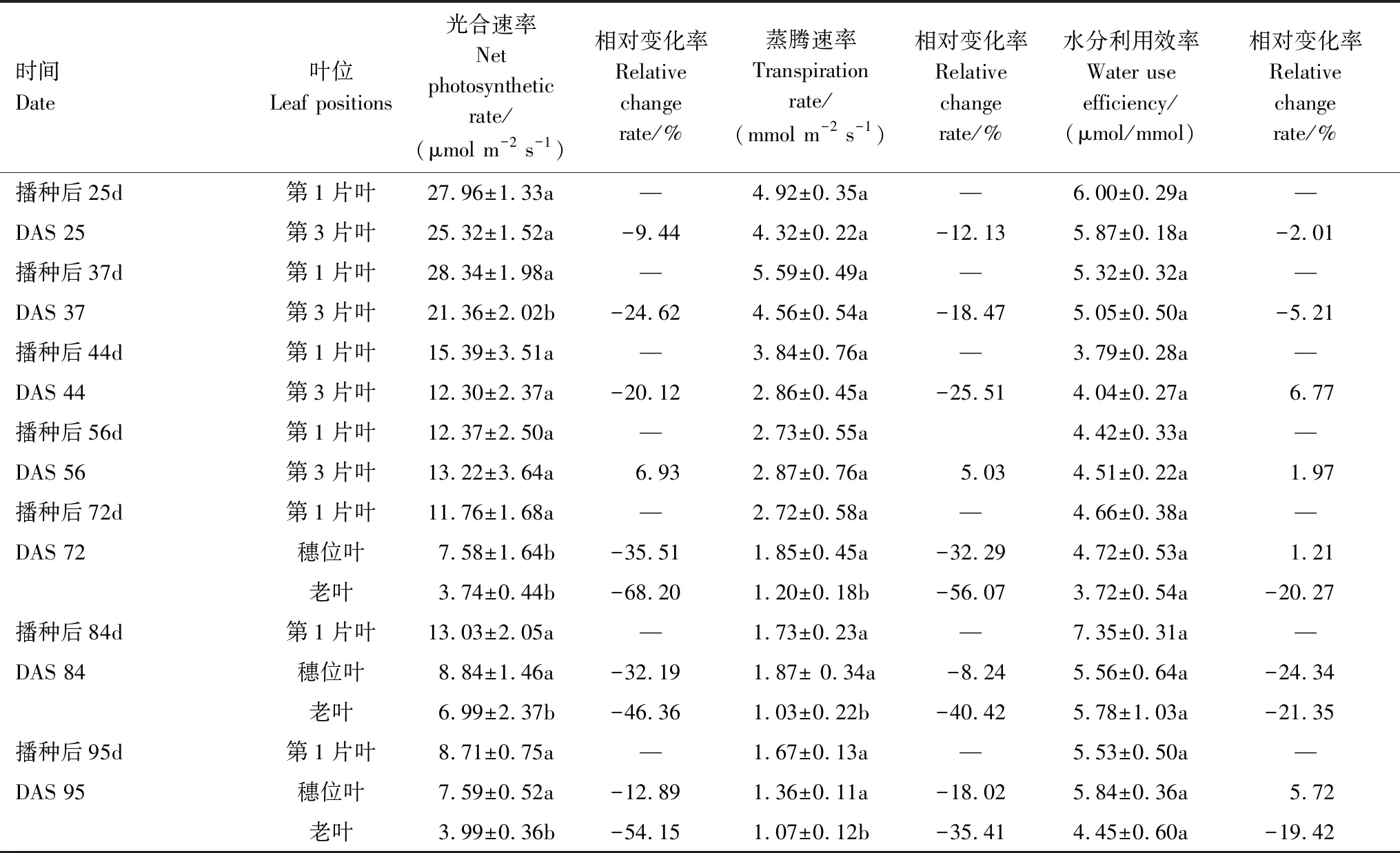
表5 不同叶位光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)的相对变化(2014年)
叶片在垂直方向上的分布意味着植物体上不同位置的叶片也存在叶龄差异,即叶片的叶龄从植株顶部到底端依次增加。在叶片完整的生命史中,光合速率和蒸腾速率首先会随叶龄增加而上升,达到最大值后或保持一段时间,然后再下降,而其最大值通常出现在叶面积达到最大之前[32]。有研究发现,在无胁迫条件下棉花植株的叶片光合速率或蒸腾速率在植株不同位置上几乎一致[33],即不同位置叶片的水分利用效率没有明显差异。本研究也表明,同一时期在相同的外界条件下,不同叶位叶片的水分利用效率不存在显著差异,即玉米叶片水分利用效率具有时间上的空间稳定性和叶龄保守性,并且光合速率和蒸腾速率的协同变化是造成这一现象的主要原因。利用稳定同位素法分析白羊草(C4植物)在不同干旱胁迫下的水分利用效率发现,新、老叶片的Δ13C无显著差异,即间接性地表明新叶和老叶的水分利用效率也无明显差异[34],这与本研究结果相一致。
近年来,大量研究表明适度的土壤水分亏缺能在一定程度上提高植物水分利用效率[5,35- 36],而且重度水分亏缺反而会导致植物水分利用效率的下降,本研究结果基本与之相符,但是土壤水分的供水能力和植物水分利用效率的定量关系仍不清楚,需要进一步深入研究。水分利用效率的研究层次大致可以划分为4个:叶片、植株、群落和生态系统,其调控因素也因研究尺度而异。目前存在的问题是不同尺度的水分利用研究存在明显的不一致性,在不同尺度或区域之间的可比较性差[37- 38]。利用碳-氧同位素对高原草地的水分利用效率分析发现,叶片水分利用效率是生态系统水分利用效率的2—3倍[39]。针叶林和阔叶林的水分利用效率在叶片尺度上存在差异,而在生态系统水平上并没有差异[40]。本研究发现不同叶位叶片的水分利用效率在干旱过程中具有时间上的空间稳定性,这为水分利用效率的尺度化研究提供了新的思路。
4 结论
利用2013—2014年连续两年玉米不同灌溉方案模拟实验资料,对不同叶位叶片的水分利用效率变化特征及其影响因素进行了探讨,得出的主要结论为:
(1)玉米第1片完全展开叶的水分利用效率在生育期内的拔节期和灌浆期具有两个明显的峰值,总体上生育期后期的水分利用效率要高于生育期前期。
(2)叶片水分利用效率与叶片生理生态特征密切相关。
(3)玉米不同叶位叶片的水分利用效率具有时间上的空间稳定性以及年龄保守性,其原因是光合速率和蒸腾速率在不同叶位之间协调变化引起的。
致谢:中国科学院植物研究所许振柱、张峰和宋健老师对试验过程给予帮助,麻雪艳、石耀辉、王秋玲、王敏政、冯晓钰、王帆在试验观测、数据采集和整理过程中给予帮助,特此致谢。
猜你喜欢
农业与技术(2023年11期)2023-06-19 02:28:18
中国农学通报(2022年29期)2022-11-25 07:28:12
干旱地区农业研究(2022年4期)2022-07-28 05:37:08
四川蚕业(2022年1期)2022-06-06 02:03:42
节水灌溉(2021年5期)2021-05-26 06:49:38
中国农业科技导报(2020年12期)2020-03-15 10:50:08
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
甘肃林业科技(2016年2期)2016-11-16 09:15:39
西南农业学报(2016年5期)2016-05-17 05:42:39
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
