
江苏省2017年禽流感流行病学调查及分析
2019-05-05 09:32韩凯凯刘青涛刘宇卓赵冬敏黄欣梅章丽娇姜亮亮宇善广崔国玉
江西农业学报 2019年4期
杨 婧,袁 朗,韩凯凯,刘青涛,刘宇卓,赵冬敏,黄欣梅,章丽娇,姜 波,姜亮亮,宇善广,崔国玉,李 银*
(1.江苏省农业科学院 兽医研究所/农业部 兽用生物制品工程技术重点实验室,江苏 南京 210014;2.山东省烟台市牟平区畜牧兽医工作站,山东 烟台 264100;3.山东省烟台市牟平区大窑兽医站,山东 烟台 264100)
禽流感(AI)是由A型禽流感病毒引发的高度接触性传染病[1]。根据禽流感的表面血凝素(HA)和神经氨酸酶(NA)抗原性的不同而分型,HA有18种亚型,NA有11种亚型[2]。根据致病性不同又可分为高致病性禽流感、低致病性禽流感和无致病性禽流感[3]。H9N2亚型禽流感病毒为低致病性禽流感病毒,呈世界性分布,并可以传播到人类和低等哺乳类动物。自1998年以来,已有多起人感染H9N2 AIV的报道[4]。此外, H9N2 AIV存在重组现象,可为形成跨种传播的新型流感病毒提供条件[5]。
研究表明,养殖场的饲养环境与鸡群感染疾病息息相关,活禽市场亦是重要的禽流感病毒感染地[6],也是与消费者日常生活密切相关的场所。因此对养殖场和活禽市场中的禽流感进行流行病学调查具有重要的公共卫生意义。本研究于2017年对江苏省南京市、常州市、无锡市、徐州市、连云港市、盐城市等地区的活禽市场、养殖场进行了禽流感流行病学调查与分析,以期为深入了解该地区禽流感感染状况和防控措施的制定提供技术支持。
HA基因是A型流感病毒致病性的主要影响因子[7],能识别两种类型的受体,分别为α-2,3-(禽流感受体)和α-2,6-半乳糖(人流感受体)连接的唾液酸。PB2蛋白含有1个色氨酸富集区[8],识别并结合宿主前mRNA 5′端的帽子结构[9];利用PB1蛋白的核酸酶活性对宿主mRNA进行裂解[10],以此作为病毒mRNA的转录引物。
禽流感病毒的基因易发生重组和漂移,为了了解2017年我国江苏地区H9N2亚型禽流感病毒的变异情况,探讨其HA、PB2基因是否发生了重组,特选取2017年所分离的4个H9N2亚型禽流感病毒分离株,对其HA、PB2基因进行了测序,分析了它们的遗传演化规律,研究了H9N2禽流感病毒的变异情况。
1 材料与方法
1.1 实验材料
自2017年1月至2017年12月,在江苏省南京市、常州市、无锡市、徐州市、连云港市、盐城市等地区的活禽市场和养殖场采集鸡、鸭、鹅拭子。
0.45 μm微孔滤膜过滤器, Millipore Ireland Ltd.产品;生理盐水,自配, pH 6.8~7.0;1%鸡红细胞溶液,实验室自制,现用现配;9~11日龄SPF鸡胚,购自南京天邦生物科技有限公司;反转录酶(M-MLV)、RNase Out、10 mmol/L dNTP、5×Buffer、10×PCR Buffer、2.5 mmol/L dNTP、25 mmol/L MgCl2、聚合酶(ExTaq)等,均购自TaKaRa公司;琼脂糖(BIOWEST AGAROSE G-10),购自Gene Company LTD; Axygen RNA/DNA提取试剂盒和胶回收试剂盒,均购自康宁生命科学有限公司; PCR仪,购自TaKaRa公司;电泳仪,购自南京普阳科学仪器研究所;全自动凝胶成像分析仪(JS-680B),购自培清有限公司;高压锅(SY-450),购自徐州圣业医疗器械有限责任公司;青霉素、链霉素、丁胺卡那霉素、制霉菌素、棉签,均购自南京医药百信药房有限责任公司。
其他材料和仪器:固体石蜡、打孔器、注射器、剪刀、镊子、离心机、超净工作台、-70 ℃冰箱、-20 ℃冰箱等。
1.2 实验方法
1.2.1 采集流调拭子样品 采集样品时,将棉签拭子插入泄殖腔和口咽部,轻轻旋转拭子2~3圈,使其充分接触采样部位。口咽拭子较容易采集,同时具有较高效价。采集粪便样品时应选择新鲜、较湿润的粪便。将采集后的棉签拭子保存在含有四抗的灭菌后pH在7.2~7.5的生理盐水中,其中四抗的浓度分别为青霉素2000 IU/mL、链霉素2 mg/mL、丁胺卡那霉素1000 IU/mL、制霉菌素1000 IU/mL。
1.2.2 处理拭子样品 将采集后的拭子样品置于-20 ℃冰箱中反复冻融3次。以8000×g离心5 min,取上清,保存在-20 ℃冰箱备用。
取用0.45 μm滤器过滤后的拭子样品上清0.2 mL,每胚尿囊腔接种9~11日龄SPF鸡胚2枚,弃去24 h内死亡的鸡胚,收取24~120 h内死亡的和120 h之后未死亡鸡胚的尿囊液,进行血凝试验。
1.2.3 血凝试验(HA试验) 在96孔微量反应板上自左向右每孔各加25 μL PBS缓冲液;在左侧第一孔中加入25 μL病毒尿囊液,混合均匀,吸取25 μL至第二孔,如此依次倍比稀释至第11孔时,弃掉25 μL,第12孔为红细胞PBS阴性对照;从右向左依次向每孔加入25 μL 1%鸡红细胞悬液,然后将96孔微量反应板在振荡器上微振几下,在37 ℃温箱中静置20 min后开始观察试验结果。
1.2.4 病毒RNA的提取 将血凝阳性样品尿囊液在无菌条件下取出200 μL,置于经高压灭菌的1.5 mL离心管中,加入200 μL的V-L试剂后,在涡旋振荡仪上混匀,静置5 min;加入75 μL V-N试剂,在涡旋振荡仪上混匀,以12000×g离心5 min;取上清,加入新的2 mL离心管中,再加入300 μL异丙醇(含无水乙醇),上下倒置混匀,转移至新的含有吸附柱的2 mL离心管中,以3000×g离心1 min;弃滤液,加入500 μL Buffer W1(含无水乙醇),以12000×g离心1 min;弃滤液,加入800 μL Buffer W2(含无水乙醇),以12000×g离心1 min;弃滤液,以12000×g离心1 min;将吸附柱置于新的1.5 mL离心管中,加入15 μL的TE试剂后,静置1 min,以12000×g离心1 min。此即为提取的总RNA。
1.2.5 病毒RNA的反转录RT和H5-H7-H9禽流感病毒三重PCR扩增 反转录体系为:RNA 12.0 μL、10 mmol/L dNTP 2.0 μL、5×Buffer 4.0 μL、Unit 12 1.0 μL、M-MLV 1.0 μL、RNase Out 0.5 μL。反转录程序为:65 ℃ 5 min,42 ℃ 60 min,98 ℃ 5 min。
H5-H7-H9禽流感病毒三重PCR扩增体系为: cDNA 2.0 μL、2.5 mmol/L dNTP 2.0 μL、10×Buffer 2.5 μL、ddH2O 13.75 μL、25 mmol/L MgCl21.5 μL、ExTaq0.25 μL、H5上游引物P10.5 μL、H5下游引物P20.5 μL、H7上游引物P30.5 μL、H7下游引物P40.5 μL、H9上游引物P50.5 μL、H9下游引物P60.5 μL。PCR扩增程序为:94 ℃ 5 min;94 ℃ 50 s,56 ℃ 50 s,72 ℃ 1 min,共30个循环;72 ℃ 10 min。
1.2.6 电泳和核酸胶纯化及测序 配制1%琼脂糖凝胶,进行电泳。调节电泳仪为稳压模式,在80 V下电泳30 min。将阳性条带切下后,用Axygen胶回收试剂盒纯化阳性条带,并送去测序。
1.2.7 H9阳性样品用SPF鸡胚纯化后测序 选择4株血凝价较高、宿主来源和分离地点不同的H9阳性样品,分别命名为: A/Chicken/Jiangsu/S01/2017(简称S01)、A/Duck/Jiangsu/S02/2017(简称S02)、A/Chicken/Jiangsu/S03/2017(简称S03)、A/Goose/Jiangsu/S04/2017(简称S04)。用有限稀释克隆法接种9~11日龄SPF鸡胚,获取较单一的H9样品,按照1.2.4~1.2.6中的方法进行HA、PB2片段的测序。
1.2.8HA、PB2基因的序列拼接和遗传演化分析 用DNAStar软件中的Seqman拼接所得HA、PB2基因序列,用MEGA 5.0分别对其进行遗传演化分析。从GenBank中获得的代表性H9N2禽流感病毒的HA、PB2基因序列作为分析比较的参考序列(见表1)。

表1 HA、PB2基因遗传演化分析的参考序列
2 结果与分析
2.1 采集流调拭子样品
自2017年1月至2017年12月,每个月份均在江苏省南京市、常州市、无锡市、徐州市、连云港市、盐城市等地区的活禽市场、养殖场,采集鸡、鸭、鹅拭子,共计2760份。
2.2 拭子样品接胚后数据分析
各个月份的拭子样品接胚后,收取尿囊液对其进行血凝试验。除了2017年1月未检测到血凝阳性样品外,其余10个月份均有血凝阳性样品。由表2可知:2017年5月的病毒阳性率最高,为36.73%;2017年9月次之,为26.92%;2017年4月居第三,为19.61%;2017年1月最低,为0%。2017年全年病毒阳性率差分的95%的置信区间为6.4124%~20.9058%。

表2 2017年1~12月流调样品数据分析结果
2.3 H5-H7-H9禽流感病毒三重RT-PCR检测血凝价阳性尿囊液数据分析
采用H5-H7-H9禽流感病毒三重RT-PCR方法对上述血凝价为阳性的鸡胚尿囊液进行鉴定,结果(表3)表明:2017年4月的H9禽流感病毒阳性率最高,为11.76%;2017年9月次之,为11.54%;2017年5月居第三位,为8.16%;2017年11月居第四位,为3.85%;其余月份均未分离到H9禽流感病毒。2017年1月至2017年12月间,H5禽流感病毒、H7禽流感病毒总的阳性率均为0%;H9禽流感病毒总的阳性率为3.26%,差分的95%置信区间为0.0780%~6.4980%。

表3 H5-H7-H9禽流感病毒三重RT-PCR检测结果
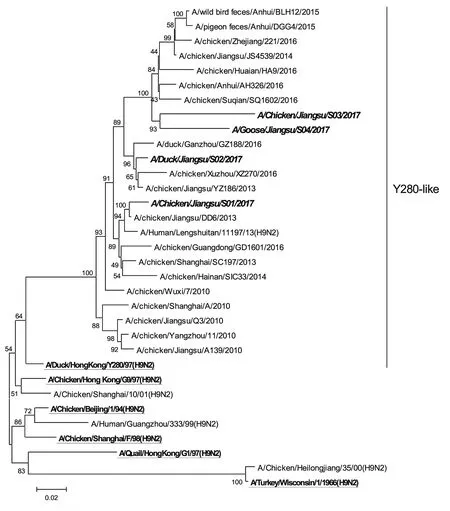
图1 4个H9N2病毒分离株HA基因的进化树
2.4 HA基因的遗传分析
用MEGA 5.0对参考序列和本试验所得的4个H9N2病毒分离株的HA基因进行遗传演化分析,得到的进化树如图1所示。
由进化树分析结果可知:这4个分离株的HA基因均属于Y280-like分支, S01、S02、S03、S04与Y280核苷酸的同源性分别为91.7%、93.0%、87.2%、89.7%,与Y280氨基酸的同源性分别为93.6%、95.1%、89.0%、91.7%。这4个分离株之间HA基因核苷酸的同源性是87.2%~92.8%,氨基酸的同源性是88.5%~94.3%,其中S03与S04之间的亲缘关系最近,两者核苷酸的同源性是92.8%,氨基酸的同源性是94.3%。
分析此4个分离株的氨基酸序列发现: HA裂解位点附近的氨基酸均为-R(K)SSR/GL-,无多个连续的碱性氨基酸插入,具有典型的低致病性禽流感病毒HA基因的特征。分析HA蛋白的受体结合位点发现:226位氨基酸均为亮氨酸(L)。
2.5 PB2基因的遗传分析
用MEGA 5.0对参考序列和本试验所得的4个H9N2病毒分离株的PB2基因进行遗传演化分析,得到的进化树见图2。
从进化树分析结果可知:这4个分离株的PB2基因均属于G9-like分支, S01、S02、S03、S04与G9核苷酸的同源性分别为90.5%、90.3%、88.1%、89.6%,与G9氨基酸的同源性分别为92.8%、92.6%、89.3%、90.7%。这4个分离株之间PB2基因核苷酸的同源性是97.0%~99.5%,氨基酸的同源性是98.3%~99.7%,其中S01与S02之间的亲缘关系最近,两者核苷酸的同源性是99.5%,氨基酸的同源性是99.7%。
分析这4株分离株的氨基酸序列发现:PB2基因第627位氨基酸均为谷氨酸(E),第701位氨基酸均为天冬氨酸(D),第448位氨基酸均为天冬酰胺(N),均未发生E627K的突变。

图2 4个H9N2病毒分离株PB2基因的进化树
3 讨论
本研究采集流调拭子样品的结果表明,2017年江苏省各地区禽流感病毒的分离率有两个高峰期,分别是4~6月、9~11月,这可能与气温环境相关,过热过冷均不适宜该病毒的繁殖。而H9亚型禽流感病毒也是在4月分离率最高,在9月次之,这与气温环境的相关性非常明显,也与病毒分离的研究结果一致。2017年未在江苏省的活禽市场和养殖场中检测出H5、H7亚型禽流感病毒,表明该省H5、H7亚型禽流感病毒的感染率较低。 对流感病毒HA蛋白的研究结果表明:第183、190和226位氨基酸是关键的受体结合位点,与病毒的受体结合特性和宿主特性相关,尤其是226位,当为谷氨酸(Q)时,其与α-2,3-半乳糖连接的唾液酸(禽流感受体)结合,当为亮氨酸(L)时,其与α-2,6-半乳糖连接的唾液酸(人流感受体)结合[11]。在本研究中,4株H9N2禽流感病毒的226位氨基酸均为亮氨酸(L),呈现出典型的人流感受体结合特性。Hatta等[12]研究发现, PB2蛋白第627位氨基酸从谷氨酸(E)到赖氨酸(K)的变异对禽流感病毒跨越种间屏障感染哺乳动物起决定性作用。本研究发现4株分离株PB2蛋白第627位氨基酸均为谷氨酸(E),没有发生E627K的突变,表明其对哺乳动物不呈高致病力。Sun等[13]对2003~2008年中国北方不同地区的22株禽源和猪源H9N2毒株进行了分析,发现同一时期不同地区病毒株之间同源性较高,但较早期病毒均有不同程度的变异和进化。本研究的进化树分析结果显示,当分离毒株的年代越为接近时,其同源性越高,与病毒分离地的相关性不大。由此表明, H9N2发生变异与时间呈现一定的相关性[14]。综上所述,本研究中的4株H9N2禽流感病毒具有结合人体的由α-2,6-半乳糖连接的唾液酸的能力,但尚未具有突破种间屏障感染哺乳动物的能力,对公共卫生安全存在一定的潜在威胁。
基于以上结果和分析,建议继续加强对江苏省各地区活禽市场、养殖场的禽流感流行病学调查。应尽早取缔活禽市场,加强养殖场综合卫生防疫管理,以确保公共卫生安全。本研究中禽流感的高发季为春秋时节,由此提示公众在春秋时节需要做好禽流感的防御工作,尽量不接触患病禽类。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
医学概论(2022年3期)2022-04-24
科学家(2022年3期)2022-04-11
首都食品与医药(2022年3期)2022-02-21
中国现代医药杂志(2019年9期)2019-10-31
教师·中(2018年4期)2018-06-02
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
家禽科学(2015年2期)2015-10-21
水禽世界(2014年3期)2014-09-27
