
蒜头果中3-酮酯酰-CoA合酶基因克隆与表达分析
2019-04-29 01:28李云琴陈中华原晓龙
中国油脂 2019年3期
李云琴,陈中华,原晓龙,王 毅
(云南省林业科学院云南省森林植物培育与开发利用重点实验室, 国家林业局云南珍稀濒特森林植物保护和繁育重点实验室,昆明 650201)
神经酸(Nervonic acid,NA)具有促进大脑发育、改善记忆、调节血脂等功效,对心血管疾病及免疫性疾病的治疗效果显著[1-3],是大脑中神经细胞及神经纤维的核心天然成分,神经酸还可以作为香料原料加以利用[4]。虽然神经酸用途广泛,对人体健康有益,但难以通过人体自身合成,需要从外界摄取。神经酸最早在鲨鱼油中获取[5],但鲨鱼资源稀少且成本高昂,加之国际社会禁止大量捕杀鲨鱼,神经酸的来源受到限制,造成资源紧缺,市场供不应求。通过化学方法[6]和生物化学[2]合成神经酸,终因副产物多、工艺路线长、得率低而失败。后来,研究发现一些植物中富含神经酸,如蒜头果(Malaniaoleifera)及盾叶木(Macarangaadenantha)种仁,鸡爪槭(Acerpalmatum)果实及遏蓝菜(Thlaspiarvense)种子[7]、元宝枫(Acertruncatum)种子[8]等。而我国特有的铁青树科(Olacaceae)蒜头果属单种属植物蒜头果,为目前发现的自然界中神经酸含量最高的植物[9]。研究显示,蒜头果种仁含油率高达64.5%[10],其果仁油脂中神经酸含量高达60%以上。因此,从蒜头果果仁中提取神经酸将为可持续开发利用植物神经酸开创一条新路[11]。
神经酸为超长链单不饱和脂肪酸(VLCMFA)[12]。VLCMFA以油酸为底物,通过超长链脂肪酸链延长循环进行碳链的延长,每循环1次延长2个碳原子,相继形成鳕油酸(20∶1Δ9c)、芥酸(22∶1Δ13c)、神经酸(24∶1Δ15c)[3]。这个延长过程受3-酮酯酰-CoA合酶、3-酮酯酰-CoA还原酶、3-羟酯酰-CoA脱水酶和羟酯酰-CoA还原酶形成的酶复合体的组合催化和调控,其中3-酮酯酰-CoA 合酶(KCS)目前被认为是关键催化步骤[13-15],在整个超长链脂肪酸延长酶复合体中起到了极其重要的作用,因此也作为限速酶被广泛应用[16-18]。目前对KCS的研究主要在油菜(Brassicanapus)[18]、大豆(Glycinemax)[19-20]、油桐(Verniciafordii)[21]、滇牡丹(Paeoniadelavayi)[22]等传统油料植物中,而对蒜头果的KCS相关研究未见报道,特别是超长链KCS在蒜头果中神经酸生物合成的作用并不清楚。因此,本研究利用转录组测序技术获得蒜头果果实中超长链KCS基因序列,同时,利用RT-PCR克隆获得该基因,并进行生物信息学分析及表达分析,为最终揭示蒜头果神经酸生物合成奠定基础。
1 材料与方法
1.1 实验材料
实验所需蒜头果树生长于文山州广南县旧莫乡岩腊村,采集花谢后1、2、3、4个月的蒜头果果实为材料,并采集花谢2个月后蒜头果树叶作为对照。样品采集后即刻放入液氮中保存。
总RNA小量提取试剂盒购自美国Qiagen公司,pESY-T3克隆试剂盒与感受态细胞购自北京全式金生物技术有限公司,LA-Taq酶、反转录酶M-MLV购自大连宝生物工程有限公司。电泳仪等。
1.2 实验方法
1.2.1 转录组分析
将花谢后3个月的蒜头果果实送到华大基因通过Illumina Hiseq2000平台进行转录组测序,对序列进行组装拼接后,通过对转录组Unigene进行本地Blast,寻找蒜头果果实转录组中的KCS基因,同时,将转录组数据中的Unigene注释到NR、NT、Swiss-Prot、KEGG、COG、GO数据库,检索注释结果中的KCS基因,并对获得的Unigene进行分析。
1.2.2 提取RNA
提取蒜头果果实RNA,按照RNA提取试剂盒说明书进行提取。分别提取每个样品的RNA后,用1%的琼脂胶进行电泳检测RNA完整性。并分别取1 μg RNA用于反转录得到cDNA。将各样品cDNA保存在-20℃冰箱备用。
1.2.3 蒜头果KCS基因克隆
通过对蒜头果转录组数据分析,获得表达量高、并拥有完整开放阅读框的KCS基因序列。以获得的KCS基因序列,设计含有起始密码子的特异引物(MoKCSF:5′-ATGGCTCAACCAAAGCTTGT-3′)和含有终止密码子的特异引物(MoKCSR:5′-TTAGATGGCCGAGACTTGCG-3′)。以蒜头果果实cDNA 为模板,用HiFi高保真聚合酶进行PCR扩增,电泳检测后将目的片段连接到pGMT载体上,并转入DH5α感受态细胞中,通过菌落PCR筛选出阳性克隆,送上海生工测序公司进行测序,最终通过序列比对确定获得KCS全长基因片段,并永久保存在-80℃冰箱。
1.2.4 蒜头果KCS蛋白生物信息学分析
用NCBI在线软件ORFFinder对KCS基因进行蛋白翻译后,并用在线软件ProtParam对KCS蛋白理化性质进行分析,KCS保守结构域分析由在线蛋白功能结构域分析软件完成,其氨基酸同源序列比对则用DNAMAN 软件完成,最终确定KCS的保守结构域。并将蒜头果KCS蛋白与美国国家生物技术信息中心(NCBI)已知其他植物的超长链KCS蛋白进行多重比对后,采用MEGA7中自带的ClusterW进行蛋白序列多重比对后,采用Neighbor-joining算法(自检举1 000 次),绘制并进行进化树分析。
1.2.5 蒜头果KCS基因不同组织表达分析
分别取蒜头果不同组织的RNA 2 μg,按照RevertAidTM First Strand cDNA Synthesis Kit试剂盒说明合成cDNA第一条链。以不同组织的cDNA为模板,用特异引物TMoKCSF(5′-ATCTTTGGTGTTCTTGATCG-3′)和TMoKCSR(5′-ATTGAATAAG CTACAATTGA-3′)进行荧光定量PCR分析,并以elongation factor 1-alpha基因作为内参基因,具体反应体系如下:分别向PCR管中加入2×SYBR Green master mix(含ROX) 12.5 μL; 引物TMo-KCSF和TMoKCSR各0.5 μL,cDNA 1 μL,最终用 PCR water nucle-free补足25 μL。PCR反应条件为:预变性温度95℃,时间10 min,然后,变性温度95℃,时间15 s;退火和延伸温度为 60℃,时间为30 s,共40个循环。反应结束后,收集信息进行Ct值分析。
2 结果与分析
2.1 转录组分析
通过Illumina Hiseq2000平台测序对蒜头果果实转录组测序,总计产出6 299 384 146 bp数据。组装结果得到165 110个Unigene。对所获得的Unigene进行本地Blast以及在NR、NT、Swiss-Prot、KEGG、COG、GO数据库比对分析, 最终获得13个可能的KCS基因片段(见表1)。

表1 转录组分析得到的KCS基因信息
由表1可知,根据已知其他植物KCS基因大小在1 kb以上,从转录组数据中最终得到大于1 kb的KCS基因片段有2个。通过蛋白序列分析以及表达量分析最后确定c55526_g1作为后续克隆分析的对象。
2.2 RNA提取与基因克隆
将蒜头果不同时期的果实和叶片样品,经过液氮磨碎后,用RNA提取试剂盒提取各样品RNA,并进行电泳检测查看其完整性,电泳结果见图1。
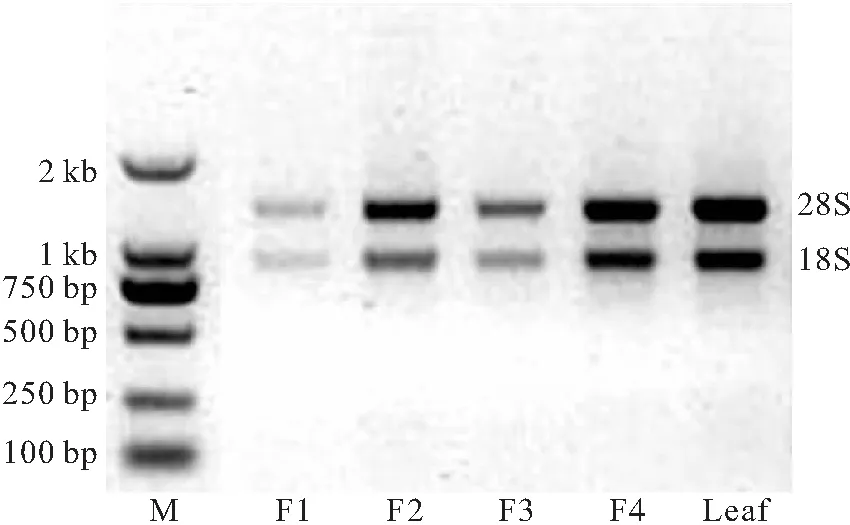
注:F1、F2、F3、F4分别为花谢1、2、3、4个月后蒜头果果实; Leaf为花谢2个月后蒜头果树叶。
图1 提取的RNA电泳结果
由图1可看出,所有样品均有完整的28S和18S两个条带,说明RNA完整性比较好。条带明亮度不一样说明提取的各个样品RNA浓度不一样。用分光光度计测量各样品RNA在260、280 nm下的吸光度。检测显示各样品OD260/OD280值均在1.8~2.1之间,适合于后续反转录实验。
基于转录组数据中获得的KCS基因片段序列的特异引物(MoKCSF和MoKCSR),以MoKCSF和MoKCSR为引物,以蒜头果花谢2个月后的果实cDNA为模板,进行高保真PCR扩增,将扩增出来的目的片段连接到克隆载体中,并进行测序比对验证。最终获得 1 539 bp的蒜头果KCS基因全长cDNA,并命名为MoKCS1。
2.3 蒜头果KCS蛋白序列分析
用NCBI在线软件ORFFinder对KCS基因进行蛋白翻译后,并用在线软件ProtParam对KCS蛋白理化性质进行分析,结果显示KCS基因共编码512个氨基酸,其相对分子质量为57 730.49,等电点为9.39。利用NCBI在线结构域分析软件(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)以及DNAMAN软件对与KCS亲缘关系近的其他植物KCS蛋白序列进行比对,见图2。由图2可以看出,该蛋白与已报道的KCS含有KCS家族的3个功能保守结构域“FGNTSSSS”“GSGFKCNSAVW”和“GMGCSA”,表明该蛋白属于KCS 家族蛋白。蒜头果KCS与榴莲(Duriozibethinus)KCS的蛋白序列同源性为80.66%,与芝麻(Sesamumindicum)KCS同源性为79.49%,与山黄麻(Tremaorientalis) KCS同源性为78.72%。
2.4 系统进化树分析
将推导出来的KCS蛋白序列在美国国家生物技术信息中心(NCBI)进行比对,用MEGA7中自带的ClusterW进行蛋白序列多重比对后,采用Neighbor-joining算法(自检举1 000 次),绘制出蒜头果KCS与其他植物的KCS蛋白的进化树,见图3。从图3可以看出,蒜头果的KCS蛋白在系统进化树上自成一支,与目前发现的KCS蛋白的亲缘关系都相对较远。
2.5 荧光定量PCR检测KCS基因不同组织表达
为了解KCS基因在蒜头果不同个体之间的表达差异,以特异引物TMoKCSF和TMoKCSR,用荧光定量PCR检测KCS基因在不同样品中的表达情况,结果见图4。由图4可以看出,KCS基因在不同个体中以花谢3个月后蒜头果果实表达量最高,花谢2个月后蒜头果果实次之,而在花谢1个月及4个月后的蒜头果果实和蒜头果叶中表达都非常微弱。

MoKCSMAQPKLVKPLIAP---SASPRLPDFKQGVKLKYVKLGYHYLVTHAMYLFLPPLAVIAAVQDzKCSMSESYPSTPLIPK---STSYKLPDFKQSVKLKYVKLGYHYLITHGMYIFLTPLAVVIAAQSiKCSMSETTASTPLIPP---SSSRKLPDFKQSVKLKYVKLGYHYLITHGMYLFLSPLVVVIAAQToKCSMSDSNPKTPLVPPQSSSSSRKLPDFKQSVKLKYVKLGYHYLITHGMYLFLSPLLIVIAAQ∗.∗∗.∗.∗.∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.∗∗∗∗.∗∗∗∗..∗∗MoKCSLSTFSLQDVHDLLGQLRYNLISVILCSSTLVFLSTLYFLTRPRPVYLVDFSCFKPDDDDKDzKCSLSTFSLQDLHDLWDHLRFNLISVILCSALLVFFSTLYFLTRPRPVFLVDFSCYKPDDA-RSiKCSVSTFSLQDLYVLWDHLRFNLISVIVCSALLVFLSTVYFLTRPRPVYLVNFSCYKPNDD-RToKCSVSTFSLQDVHDLWDHLRFNLISVILCSTLLVFLLTLYFLTRPRPVYLVDFSCYKPEDA-R.∗∗∗∗∗∗∗.∗.∗∗.∗∗∗∗∗∗.∗∗.∗∗∗∗.∗∗∗∗∗∗∗∗∗.∗∗∗∗∗.∗∗∗.MoKCSKCSWQRFMKCSESRGTFTEENIEFQRKIMARSGIGESTYLPPAVMKIPPNSSMAEAREEADzKCSKCTRQIFMERSRLTGTFTEENLEFQRKILERSGLGESTYLPEAVLRVPPNPCMAEARKEASiKCSKCTRQIFMERSRLTGTFTEENLEFQRKILERSGLGESTYLPEAVLRVPPNPCMAEARKEAToKCSKCTRQIFMDRSRLTGTFTEENLEFQRKILERSGLGESTYLPEAVLRVPPNPCMAEARKEA∗∗.∗∗∗∗∗∗∗∗∗∗∗.∗∗∗∗∗∗.∗∗∗.∗∗∗∗∗∗∗∗∗...∗∗∗.∗∗∗∗∗∗∗MoKCSKMIIFGVLDRLFEKTSVKPKDISILIVNCSLFNPVPSLSAMVVNHYKLRGNTRTYNLGGMDzKCSETVMFGALDQLFEKTCLKPKDIGILIVNCSLFNPTPSLSAMVINHYNLRGNIISYNLGGMSiKCSETVMFGAIDELLAKTSLKPKDIGILIVNCSLFNPTPSLSAMVINHYKLRGNIISYNLGGMToKCSELVMFGALDQLFEKTSLKPKDIGILIVNCSLFNPTPSLSAMIVNHYKLRGNIISYNLGGM..∗∗.∗∗∗∗..∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗..∗∗∗∗∗∗∗.∗∗∗∗∗∗MoKCSGCSAGLISIDLAKDLLRVHPNSYALVVSMESITMNWYFGNERSMIVPNCLFRMGGAAVLLDzKCSGCSAGLISIDLAKDLLLAHPNSYALVISMENITLNWYFGNERSMLVSNCLFRMGGAAILLSiKCSGCSAGLISIDLAKDLLQVHPNTYALVISMENITLNWYFGNERSMLVSNCLFRMGGAAILLToKCSGCSAGLISIDLAKDLLQVHPNSYALVISMENITLNWYFGNDRSMLVSNCLFRMGGAAILL∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.∗∗∗∗.∗∗∗∗∗.∗∗∗∗∗∗.∗∗∗.∗∗∗∗∗∗∗∗∗∗∗.∗∗MoKCSSNKMSDRWHSKYKLVHTVRTHKGSDDKCYTCVTQREDSIGKIGISLSKDLMAVAGDALKADzKCSSNKRSDGWRSKYRLVQTVRTHKGADDKCFTCVTQKEDSTGRIGVSLSKDLMAVAGDALKTSiKCSSNKRSDYWRSKYRLVHTVRTHKGSDDKCFSCVTQMEDSKGKIGVALSKDLMAVAGDALKTToKCSSNKRSDRWRSKYQLVHTVRTNKGADDKCFSCVTQKEDDMGKIGVSLSKDLMAVAGDALKT∗∗∗∗∗∗.∗∗∗.∗∗.∗∗∗∗.∗∗.∗∗∗∗..∗∗∗∗∗∗∗.∗∗..∗∗∗∗∗∗∗∗∗∗∗∗∗∗.MoKCSNITTLGPLVLPMSEQLLFFATLVGRKFFKVKVKPYIPDFKLAFEHFCIHAGGRAVLDELQDzKCSNITTLGPLVLPMSEQLLFFATLVARKLFKMKVKPYIPDFKLAFEHFCIHAGGRAVLDELESiKCSNITTLGPLVLPMSEQLLFFATLVGKKLFKMKLKPYIPDFKLAFEHFCIHAGGRAVLDELEToKCSNITTLGPLVLPMSEQLLFFATLVGKKLFKMKLKPYIPDFKLAFEHFCIHAGGRAVLDELE∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.∗∗∗.∗.∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.MoKCSKNLQLTDWHIEPSRMTLYRFGNTSSSSLWYELAYTEAKGRVKKGDRTWQIAFGSGFKCNSDzKCSKNLQLSDWHMEPSRMTLYRFGNTSSSSLWYELAYTEAKGRVRKGDRTWQIAFGSGFKCNSSiKCSKNLQLSDWHMEPSRMTLYRFGNTSSSSLWYELAYSEAKGRIKRGDRTWQIAFGSGFKCNSToKCSKNLQLSDWHMEPSRMTLHRFGNTSSSSLWYELAYSEAKGRIRKGDRTWQIAFGSGFKCNS∗∗∗∗∗.∗∗∗.∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.∗∗∗∗∗...∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗MoKCSAVWEALRTINPAKEKNPWMTEIHQFPINVPQVSAI--DzKCSAVWKALRTINPAKEKNPWMDEIHQFPVDVPKVSAI--SiKCSAVWKALRTINPAKEKNPWMDEIHQFPVEVPKVSAI--ToKCSAVWKAIRTVNPAKEKNPWMDEIHQFPVDVPKVSALKA∗∗∗∗.∗∗.∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗∗.∗∗.∗∗∗.
注:MoKCS.蒜头果KCS(登录号:MK210592);DzKCS.榴莲KCS(登录号:XP_022729498.1);SiKCS.芝麻KCS(登录号:XP_011081914.1);ToKCS.山黄麻KCS(登录号:PON90748.1)。
图2 KCS蛋白序列比对

图3 蒜头果KCS与其他植物KCS蛋白系统进化树

注:MoF1、MoF2、MoF3、MoF4分别为花谢1、2、3、4个月后蒜头果果实KCS基因;MoLe为花谢2个月后蒜头果树叶KCS基因。
图4 蒜头果KCS基因表达情况
3 讨 论
神经酸(C24∶1)具有独特的工业用途和潜在的药用保健功效,近年来得到了国内外的广泛关注。特别是从植物中获取神经酸受到许多研究者的青睐。虽然很多植物含有神经酸,但是大多含量很低,蒜头果是目前神经酸含量最高的植物,因此可以在揭示蒜头果中神经酸生物合成机理的情况下,利用分子辅助育种等方式培育出高产神经酸的蒜头果新品种,或利用基因工程通过异源表达的方式利用酵母[23-24]大量获得神经酸。因此,本研究开展蒜头果中神经酸生物合成关键酶KCS基因的克隆及表达分析。
目前已有一些关于KCS基因通过异源表达提高神经酸含量的研究,如Guo等[18]报道通过引入银扇草的KCS基因来增加酵母和转基因植物中神经酸含量;Taylor等[25]报道了在碎米荠属油籽中,一种KCS基因的分子克隆和特性,以及其在碎米荠属油籽中的异源表达,为潜在的医疗和工业用途提供高神经酸油。针对MoKCS基因,未来的研究可以将MoKCS基因导入目前油脂高产的植物中,通过MoKCS及超长链脂肪酸生物合成复合体合成神经酸;同时,也可以将MoKCS导入目前已经开发出来的高产油脂酵母中,通过酵母发酵的方式获得神经酸。最终为神经酸产业发展奠定坚实的基础。
4 结 论
本研究利用转录组测序技术首先获得蒜头果果实中KCS基因序列,并利用RT-PCR技术克隆获得全长开放阅读框MoKCS1,其全长1 539 bp,编码含512个氨基酸,生物信息学分析结果显示,MoKCS1含有KCS蛋白家族的保守结构域“FGNTSSSS”“GSGFKCNSAVW”和“GMGCSA”,与已报道的KCS具有高度一致性,表明该蛋白属于KCS家族蛋白。蒜头果KCS与榴莲(Duriozibethinus)KCS的蛋白序列同源性最高,为80.66%,与芝麻(Sesamumindicum)和山黄麻(Tremaorientalis)KCS同源性分别为79.49%、78.72%。通过与NCBI中已知KCS构建分子进化树分析,显示MoKCS1非常特殊,自成一支,说明MoKCS1非常特殊。荧光定量PCR显示MoKCS1在不同个体中以花谢3个月的蒜头果果实表达量最高,而花谢3个月正是蒜头果果实膨大期,神经酸的累积期。这说明MoKCS1与蒜头果神经酸生物合成密切相关。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
环球时报(2022-09-20)2022-09-20
中国种业(2021年11期)2021-11-25
今日农业(2020年24期)2020-12-15
东方少年·阅读与作文(2019年2期)2019-06-11
生物学教学(2019年3期)2019-03-22
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
