
广东内伶仃岛猕猴种群动态及种群生存力分析
2019-04-18 02:52:18楚原梦冉昝启杰徐华林徐润林
野生动物学报 2019年2期
楚原梦冉 昝启杰 杨 琼 张 鹏,3 徐华林 徐润林*
(1.中山大学生命科学学院,广州,510275;2.广东内伶仃福田国家级自然保护区管理局,深圳,518040;3.中山大学社会与人类学学院,广州,510275)
猕猴(Macacamulatta)隶属哺乳纲(Mamalia)灵长目(Primates)猴科(Cercopithecidae),为国家 Ⅱ 级重点保护野生动物,是世界上分布最广,栖息地类型最为多样的非人灵长类动物。在我国的西南、华南、华北、华东以及西北的部分地区均有分布。20世纪80年代,刘振河第一次记录到珠江口(内伶仃岛、担杆岛、二洲岛)及川山群岛(上川岛)海区4个岛屿上有野生猕猴的分布。由于特殊的地理位置和自然环境,内伶仃等4个岛屿猕猴种群对我国灵长类动物岛屿生物地理学、保护生物学等方向的研究有着重要的价值[1]。
内伶仃岛作为国家级自然保护区,是珠江口现存有猕猴的4个海岛中面积小、植被保存最好的岛屿,其优越的生境条件为野生动物的生存提供了丰富的食物资源和良好的栖息环境。猕猴是内伶仃岛岛屿生境中重要的动物类群,受岛屿隔离的影响非常明显,因而是理想的研究对象。近年来在内伶仃岛已对猕猴的种群分布格局及行为特征、食源植物分布、栖息地特征及生境质量、繁殖模式与社会结构等问题进行了研究[2-5]。种群动态研究,需要以长年的种群数量及分布格局为基础,研究种群大小或数量在时间和空间上的变动规律,这不仅是种群生态学的核心问题,更是体现生物物种受生存环境限制的一个重要指标[6-8]。通过对其研究可以了解在栖息地有限的生态环境下种群能达到的最大数量,为更加合理地保护野生动物提供理论依据[9-11]。
因此,本文是在内伶仃岛1984—2016年猕猴种群数量及分布变化格局的研究基础上,深入研究了种群数动态变化,并应用漩涡模型(VORTEX 10)以2016年为时间起点,通过100次模拟,预测了内伶仃岛野生猕猴未来100年的种群变动趋势。最终通过参数敏感度分析,探讨影响猕猴种群动态的主要因子,并针对研究结果与实际情况提出保护管理策略。
1 研究地点
广东省内伶仃岛位于深圳西南深圳湾外,珠江口东侧的伶仃洋。地理位置为22°23′49″—22°25′35″N,113°46′18″—113°49′49″E,东距香港6 n mile,西距珠海13 n mile,北离深圳蛇口约9 n mile。全岛西北至东南长约4 km,南北宽约2 km,最高峰尖峰山高度为340.9 m,总陆地面积为4.98 km2。
该岛属南亚热带季风气候。年平均气温为23.0℃,1月为全年最冷月,平均11.5℃,7月为全年最热月,平均32.2℃,1953年以来记录到的最高气温为38.7℃,最低气温为0.2℃。雨量充沛,年降雨量2 055.80 mm,全年无霜。植物生长茂盛,维管束植物600多种,植被覆盖率80%以上。主要植被类型有:南亚热带常绿针叶林、南亚热带针阔叶混交林、南亚热带常绿阔叶林、南亚热带红树林、南亚热带常绿灌丛和南亚热带灌草丛等[12-13]。为猕猴种群的生存和发展提供了良好的自然条件。
2 研究方法
2.1 内伶仃岛猕猴种群现状
在内伶仃猕猴种群中,选取了两群作为主要观察对象。观察群每天在11:00以少量人工投食外,其他时间猴群均处于自由状态。2014年开始进行个体编号识别,并持续跟踪观察记录。
通过整理广东内伶仃福田国家级自然保护区提供的由1989—2008年的历史猴群种群数量及分布数据,以及查阅相关的文献,对内伶仃岛猕猴种群的发展历史进行汇总。为保证调查的一致性,通过运用与前人一致的定点调查法[3,14-15],于2014—2016年的1月,连续3年(共90 d)的野外调查获得了内伶仃岛猕猴现今的种群和分布格局数据。最终,将3年调查结果的平均值作为VORTEX模型的初始种群。
2.2 种群繁殖参数估计
2.2.1 年龄结构与性比
猕猴是一种群居性较强的物种,个体间通过相互作用而构成一个有组织的群体。年龄结构和性比是猕猴种群的主要结构特征,共同影响猕猴种群的增长及动态变化[16]。根据2014—2016年观察群猕猴雌性发情交配期和产仔期的记录,内伶仃猕猴的出生性比为20雌∶22雄=0.9∶1,与1∶1没有显著差异,故此本研究将内伶仃岛猕猴种群出生性比定为1∶1。在观察群中当年生婴猴(0—1岁)和青少年猴(2—4岁)分别占20%以上,成年个体(4—12岁)在猴群中的比例达42%以上,老年个体(≥13岁)比例较小,不足10%。
2.2.2 繁殖系统
猕猴的交配体制为一雄多雌制的婚配制度。内伶仃岛猕猴于每年9—12月发情。雄性在3.5岁性成熟,4岁时便可以参与繁殖,繁殖能力较强,一般繁殖寿命可达20年[17]。在实际观察中,我们也曾记录过老年怀孕的个体,因此本研究中将猕猴的最大繁殖年龄拟定为20岁。
借鉴前人的研究结果以及内伶仃岛猕猴实际的繁殖情况,拟定内伶仃岛猕猴的种群增长为密度制约型,即参加繁殖的雌性占全部成年雌性的比例P(N)会随种群大小N的变化而变化。因此选用的漩涡模型公式为:
K为环境容纳量。其中P(K)表示种群达到环境容纳量时,参加繁殖的雌体占总成年雌体的比率,P(0)表示种群大小接近0时的比率。B表示P(N)随N的变化强度,当P(N)为N的二次方程,则能更好模拟密度制约型种群的増长。A为阿利效应(Alle Effect),表示种群数量极低时寻觅配偶难度造成配对率的降低,在VORTEX中取默认值1。拟猕猴种群达到K值时P(N)为20%,种群接近0值时P(0)为80%。
2.2.3 繁殖率
根据3年的观察记录,具备产仔能力雌性中,77%的个体3年内生产1胎,13.1%的个体3年内生产2胎。连续2次生产时间间隔最短为295 d,最长为447 d,平均(362±16)d。未出现双胞胎的现象。因此,拟定内伶仃岛猕猴单胎比例为100%。
2.2.4 死亡率
由于较低的捕食压力和人为干扰,内伶仃岛猕猴一般为自然死亡。本研究结合保护区部分历史记录和实际调查,统计了猕猴种群各年龄阶段的死亡率情况(表1)。
表1 内伶仃岛猕猴种群死亡率

Tab.1 Mortality investigation of Macaca mulatta in the reserve
2.2.5 近交衰退
在VORTEX模型中,应用致死当量(Lethal equivalents)来表示种群由于近交造成的衰退程度,并模拟种群遗传基因变化情况。Ralls等对兽类的致死等价系数研究后认为,每个二倍体平均有3.14个致死基因当量[18]。依此观点,本研究选择3.14作为VORTEX软件致死等价系数。
2.2.6 交配垄断
在VORTEX模型中,估计物种的交配垄断有3个指标:繁殖系统中的雄猴比例(%,Male in breeding pool),平均每个繁殖周期能够生育后代的雄猴比例(%,Mean successfully siring offspring in the average breeding cycle)及每年繁殖雄猴的平均子女数(Males number of litters sired by successful males per-year)。根据泊松分布,只需选择输入其中1个指标,便可计算出其他2个指标。根据在繁殖期的记录,内伶仃岛猕猴约有80%的成年雄性参与繁殖。
2.2.7 环境容纳量
根据常弘等的报道[3],内伶仃岛猕猴的理论容纳量在1 200—1 400只,在本次模拟过程选择1 400只为最大种群容纳量。
2.2.8 其他参数
VORTEX模型中,将收获(Harvest)、人为补充(Supplementation)、自然灾害(Catastrophes)也作为模拟参数,但内伶仃岛作为一个封闭的岛屿受外界干扰极小,加之保护区管理措施得当,近50年来也未出现疫情的爆发,未来出现自然灾害的可能性也极低。故本研究将以上参数都拟定为0。
3 结果
3.1 内伶仃岛种群数量及分布格局的动态变化
根据2014—2016年的调查统计结果,内伶仃岛现有猕猴24群,共计猕猴989只(989±100),其中最大的猕猴群体数量达50只以上,最小的群体为25只,平均每群(32±9)只。若将独猴的数量计算在内,则在有效调查区域内的猕猴种群总数约为1 000只。群体平均大小为33只。通过最新的调查结果,并结合内伶仃岛25年的研究历史资料(表2),可发现内伶仃岛猕猴种群呈现由集中到均匀化的分布趋势,由最初的主要集中分布在西南一侧,到如今除投食群在岛内东南部逐渐壮大外,其余地区均有猴群分布(图1)。
表2 内伶仃岛猕猴种群调查数据

Tab.2 The population size of Macaca mulatta in the reserve

图1 内伶仃岛猕猴种群近30年的分布格局变化Fig.1 The distribution of group during last 30 years
3.2 理想环境中种群动态变化趋势
从模拟结果来看,在理想环境中(没有灾害、捕杀等),内伶仃岛猕猴种群在最初20年内迅速增长,从1 000只增长到1 200只,并维持在容纳量范围内。随后种群数量基本稳定并维持在1 200只左右。其内禀增长率r=0.024,周期增长率λ=1.047,净生殖率R0=1.552,雌体的平均世代时间T=7.55,雄体的平均世代时间T=6.74。
在理想环境中,内伶仃岛猕猴种群的基因杂合率随时间推移有缓慢下降的趋势。近交繁殖率系数呈逐渐增加的趋势,但始终维持在0.02以内(图2,B),100年后随着种群的发展近交程度相对较低,能够满足种群的健康发展。

图2 理想情况下经过100次模拟后种群数量动态变化曲线(A)和近亲繁殖率系数(B)Fig.2 Mean population size over time(A)and inbreeding coefficient(B)for the base model over 100 simulations
3.3 参数灵敏度分析
通过理想模型各参数与其对种群动态趋势变化进行分析,以确定不同参数因子对种群动态的影响[8]。根据计算公式:Sx=[(Δx/x)÷(Δp/p)],其中Δx/x代表种群变化率,Δp/p代表参数变化率。影响内伶仃岛猕猴种群动态参数灵敏度指数见表3。
表3 种群动态因子参数灵敏度指数
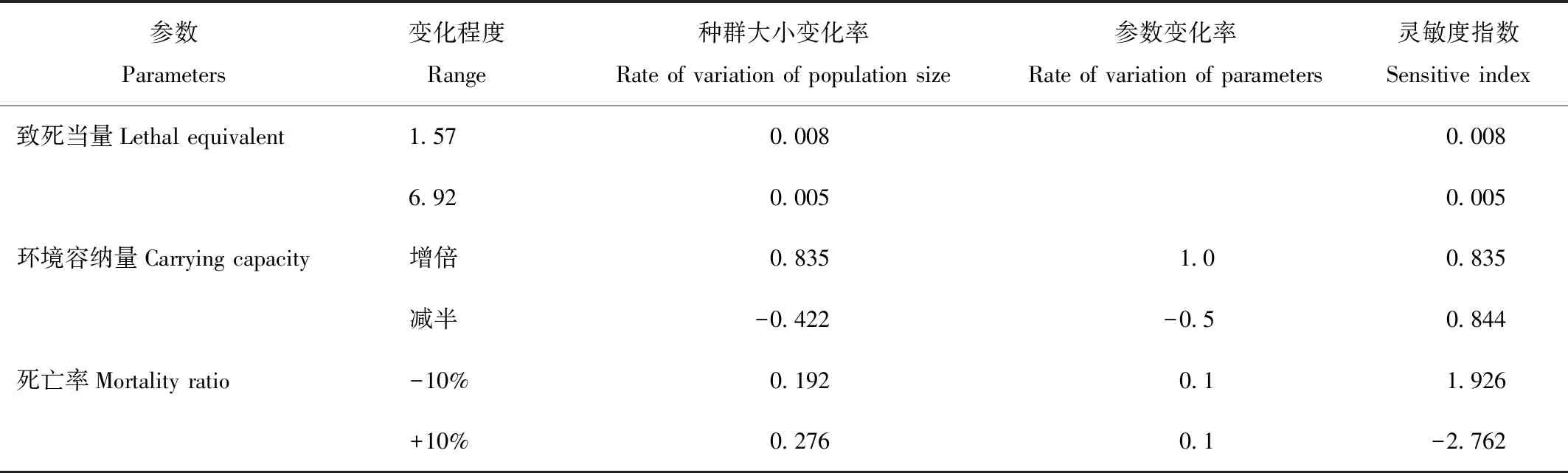
Tab.3 Sensitive indexes of different of factors affecting the dynamics of the population
3.3.1 近交衰退对猕猴种群数量的影响
对猕猴种群在有、无近交时的种群动态变化进行的模拟显示,较高的近交致死等价系数(LE=6.92),在50年内对种群数量的增长有较明显的抑制,但对种群的发展不会产生显著的影响(图3,A)。
3.3.2 环境容纳量对猕猴种群数量的影响
在不同的环境容纳量下进行的模拟结果显示,当环境容纳量减半时(K=700),猕猴种群数量下降明显,100年后减少了50%,仅为582只;环境容纳量在增倍时(K=2 800),种群数量会持续繁殖,100年后种群数量可达2 647只。
3.3.3 死亡率对猕猴种群数量的影响
在种群容纳量不变(K=1 400)情况下,猴群死亡率增加10%时种群数量迅速降低,内禀增长率r=-0.002 3,100年后种群数量下降至700只左右;但猴群死亡率减少10%时,种群内禀増长率为r=0.031,100年后种群数量可达2 300只左右。
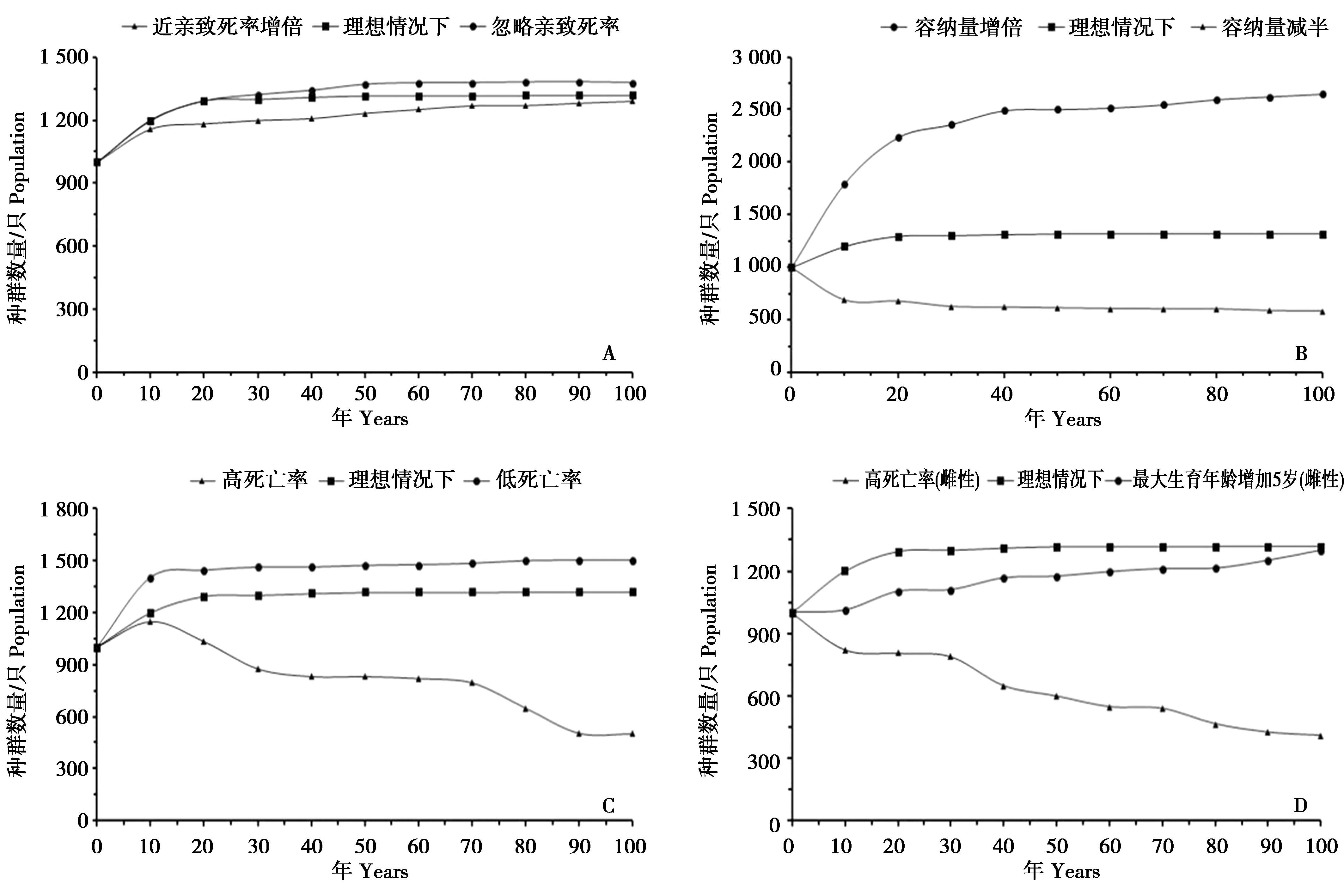
图3 在不同情况下经过100次模拟后猕猴种群数量动态变化曲线Fig.3 The changes of population number in different lethal equivalents(A),mortality(B),carrying capacity(C)and flame survival rate(D)within 100 times
4 讨论
4.1 种群分布格局的动态变化
由于没有较为连续性的种群数量调查数据,因此在本研究中很难模拟出内伶仃岛猕猴种群增长模型,但从现有的6次调查数据来看,其基本符合“S”形增长曲线(图4)。具体表现1989—2000年为猕猴种群数量的快速增长期,2000年以后种群增长速度有所下降。从种群分布格局发展变化角度看,在30年的时间里,内伶仃岛猕猴种群发展情况良好,并呈现出均匀化分布的趋势。出现这一现象的主要原因可能是:(1)内伶仃岛植被差异性不大,且总体的分布较为均匀。(2)地形比较简单,海拔落差幅度相对较小,从而对猴群的分布没能造成直接的影响。
比较还发现,通过VORTEX软件模拟所得到的猕猴的内禀增长率较常弘等所估计的数值略低(r=0.69)[3],出现这种差异的原因可能与某些种群参数估计上的差异有关,同时,也可能与静态生命表本身的缺陷有关。这种情况在利用VORTEX对长江江豚(Neophocaenaphocaenoidesasiaeorientalis)和野生大熊猫(Ailuropodamelanoleuca)进行种群生存力分析时也有所体现[19-20]。由此可见,种群数量调查的连续性在种群生存力分析时尤为重要。

图4 内伶仃岛猕猴种群数量动态变化模拟值与真实值之间的差异Fig.4 The different between the analog value and the census results of Macaca mulatta
4.2 影响种群持续发展的主要因素
通过模拟100年后内伶仃岛猕猴种群动态变化趋势可以发现,在理想环境中,猕猴种群灭绝概率为0%,并能维持在环境容纳量水平内。这充分说明,内伶仃岛猕猴种群具有较强繁殖能力。参数灵敏度分析表明,猴群死亡率是影响猕猴种群发展的重要因素,同时环境容纳量、近交繁殖也可能左右着猴群未来的发展方向。
不同年龄段死亡率高低决定着猕猴种群的发展。灵敏度分析显示,猕猴种群对死亡率较为敏感,尤其是雌性死亡率关系到种群的未来发展。雌性死亡率的轻微变化都会对整个种群的增长速度产生较明显的作用,在其他参数不发生改变的情况下,调整雌性猕猴死亡率(增加10%)及最大繁殖年龄(减少5岁)都会对整个猕猴种群产生影响(图3,D)。同时,雌性个体充当着生育者的角色,任何雌性的死亡都意味着遗传多样性的流逝,而最大繁殖年龄可能直接对繁殖速度产生影响,这与对海南长臂猿(Nomascushainanus)的研究结果一致[21-23]。
环境容纳量为影响内伶仃岛猕猴种群增长的另一个限制因子。内伶仃岛完好的生境和保护管理措施,使得猕猴种群能在30年的时间里迅速发展壮大,但当模拟种群容纳量减半时,100年时间里种群数量下降迅速。因此,在内伶仃岛需特别注意保持现有生境的完整性,避免由于外来物种的入侵,割裂猕猴现有的生境。种群容纳量能保证猕猴维持一定水平的遗传多样性,并适应环境的随机变化,是保证种群长期存活的根本因素。但由于侧重点不同,估算环境容纳量的不同方法,可能会出现差异较大的结果。例如,如果从食物资源角度分析,内伶仃岛的猕猴食源植物达200多种[15]。那么,只要猕猴的主要食源植物不受灾害等影响造成大面积枯亡,环境容纳量将不会成为其种群发展的限制因子。但是,如果从家域面积角度分析,岛内猕猴的家域面积约0.6—0.9 km2(3年观察数据),由此得出环境容纳量又会低于本研究的设置值。因此,只有通过常年的观察和数据的积累才能更加准确地去估算种群容纳量。
一个物种要长期生存,必须要有足够的遗传变异量来适应环境变化,即物种产生的遗传变异必须等于遗传漂变的损失量。遗传漂变使种群每个世代损失1/2Ne(Ne为有效种群数量)的遗传多样性[24]。从本研究的结果看,内伶仃岛猕猴种群动态在有无近交衰退的情况下,种群数量的发展在未来前50年内会表现出一定的影响,50年后的发展基本相同。出现这一结果的原因可能是:(1)内伶仃岛猕猴种群是由最初的200只发展到目前的1 000只左右,其近交繁殖系数可维持在1%左右,因此,近交衰退对猕猴种群的影响并不显著。(2)在自然选择的过程中,隐性致死基因会不断减少,近交产生的后代个体会在成年前死亡,从而大大降低了隐性致死基因的比例。但是,由于封闭的地理环境,猴群无法与外界实现基因交流,因此近交衰退现象仍然是影响内伶仃岛猕猴种群数量进一步发展的关键因素。
4.3 保护和管理建议
根据本研究结果,特对内伶仃岛猕猴种群保护和管理提出以下建议。
做好应对突发事件的应急预案。未来的保护工作需进一步加强疫情的监测和预防工作,防范传染疾病的爆发。猕猴种群数量除受本身生殖因素的影响外,目前内伶仃岛局部地带出现绞杀性的有害藤本植物薇甘菊(Mikaniamicrantha)等外来物种入侵[25-26],对植物群落发展产生不利影响,有可能造成猕猴种群数容纳量降低,根据实际情况制定详细的应对自然灾害、极端恶劣天气、火灾等情况发生的应急预案,避免现有栖息地遭到破坏。
精细化管理,实现社会效益与生态效益的统一。通过各种管理手段将工作的每一个细节精细化、数据化,从而提高管理工作的执行力和效率。管理数据化是科学指导保护管理的基础,也是大量研究工作的重点内容。只有通过常年科研数据的积累才能更好地履行保护区的职能,实现社会效益与生态效益的统一。
构建长效的管理模式。种群动态研究需要常年的数据积累,本研究中发现保护区过去在猕猴种群数量监控方面的持续性不甚理想,难免会对将来种群动态的估计带来困难。因此,需要注意种群数量监控工作持续性和连贯性,同时结合其他方式,如决策分析、专家意见、权衡分析、多维方法和管理策略模型,可以弥补取样误差、缺乏验证等不足,同时提供更高效的管理模式。
提高保护区工作人员整体的科学素养。在保护区内对护林员进行培训,并定期及不定期组织其进行参观和学习,提高其科学素养,以更好地开展保护工作。此外,规范护林员的巡护记录,做好所在管护区域的信息资料收集,及时发现问题,采取相应措施。
致谢:本工作得到深圳市城市管理局项目(2015-1)资助。同时,广东内伶仃福田国家级自然保护区管理局工作人员、中山大学生命科学学院常弘教授为我们提供有关的数据和资料。对他们以及所有参考文献的作者,我们表示衷心的感谢!
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
小猕猴学习画刊(2022年10期)2022-01-01 04:48:21
童话世界(2020年32期)2020-12-25 02:59:14
小猕猴智力画刊(2019年4期)2019-05-08 21:56:28
小猕猴智力画刊(2019年3期)2019-04-19 00:02:10
红土地(2018年7期)2018-09-26 03:07:38
领导文萃(2016年23期)2016-12-16 16:06:35
当代畜禽养殖业(2014年10期)2014-02-27 07:59:49
今日中学生(初二版)(2013年11期)2014-01-23 05:33:58
中学生物学(2008年6期)2008-08-29 09:23:38
