
钝裂银莲花主要色素成分及适应机制的研究
2019-04-03 08:02吕婉灵刘左军张小瑞王玉贤
天然产物研究与开发 2019年3期
吕婉灵,刘左军,张小瑞,王玉贤
兰州理工大学生命科学与工程学院,兰州 730050
钝裂银莲花(Anemoneobtusiloba)为毛茛科(Ranunculaceae)银莲花属(Anemone L.)中的一种多年生草本植物,单花顶生,一般产生1~3个独立花茎,花色有黄色、浅黄色和白色3种[1]。主要生长在海拔2900~4000 m的高山草地或铁杉林下。分布区内年平均降水量534 mm,年平均温度为1.6 ℃,从植物区系组成和水热特征来看,属于典型的高寒草甸[2]。目前,对钝裂银莲花的研究主要集中在它的药用价值、交配系统特征和花期资源分配[3],以及在不同海拔条件下钝裂银莲花的繁殖分配和繁育系统[4]等方面,对其花色素方面的研究几乎未见报道。在寒冷的高原地区,钝裂银莲花有其自身的适应机制。近年来高原地区环境变化显著,钝裂银莲花出现了明显的白色花减少,以及浅黄色、黄色花花增多的现象,猜想这一现象可能与其对环境的适应性与生物进化有关。植物的花色素主要有类黄酮、类胡萝卜素和生物碱三种[5],类黄酮又分为黄酮、黄酮醇、黄烷酮和花色苷,黄酮和黄酮醇呈现黄色和无色,花色苷控制红、蓝和紫等颜色,可以分为三种:花葵素、飞燕草素和花青素,它们一般在同种花中单独存在;类胡萝卜素包括胡萝卜素和叶黄素,呈现黄色、橙色、红色和紫色;生物碱是甜菜碱、小檗碱、罂粟碱等的统称,甜菜碱呈无色,罂粟碱呈黄色,小檗碱呈深橙色或黄色。而含有花色素的花都不含有生物碱[6],薛德艳[7]等已成功提取钝裂银莲花的类黄酮和类胡萝卜素,所以本实验重点分析这两类色素。故本研究通过高效液相色谱、液质联用分析黄色、浅黄色和白色的钝裂银莲花花色素成分及含量,确定其导致花色不同的主要色素,进一步分析合成该色素的关键酶。
1 实验材料与方法
1.1 实验材料
实验材料为甘肃省合作(103°22′ E,35°18′ N)的白色、浅黄色、黄色钝裂银莲花,于七月上旬采其盛花期花瓣,去除花蕊,冲洗干净,80 ℃杀青30 min,然后 60 ℃烘干粉碎成末,装于自封袋避光干燥保存。
1.2 紫外-可见分光光度计光谱分析
1.2.1 类黄酮光谱分析
分别称取0.1 g各色干花瓣,加入5 mL甲醇一甲酸(98∶2,v/v)抽提溶剂,黑暗条件下浸提24 h,过滤,收集虑液,滤渣分次加人3 mL,2 mL抽提液,进行第2、3次的抽提,合并收集的虑液,定容至10 mL。用 UV-3000 型紫外-分光光度计在 200~700 nm内进行扫描,比色皿光径1 cm。
1.2.2 类胡萝卜素光谱分析
分别称取0.1 g干花瓣,加入5 mL丙酮:乙醇(石油醚)1∶1的混合溶液,避光浸提24 h,过滤,再加入5 mL浸提,合并滤液,定容至10 mL。用 UV-3000 型紫外-分光光度计在 200~700 nm内进行扫描,比色皿光径1 cm。
1.3 花色素定性定量分析
类黄酮样品制备:取0.1 g干花瓣,加入5 mL甲醇一甲酸(98∶2,v/v)抽提溶剂,于4 ℃冰箱避光浸提24 h,取上清液用滤纸粗滤后,经微孔滤膜(脂溶性0.45 μm)过滤。标准品:槲皮素、山奈酚、木犀草素、芦丁购于上海源叶生物科技公司。
采用Waters 2685型高效液相色谱仪,Agilent 6460 LC/MS型液质联用仪,Zorbax SB-C18色谱柱(4.6 mm×150 mm,5 μm)。色谱条件:流动相组成:A相0.1%磷酸一水溶液;B相乙腈溶液,洗脱程序如下:0 min,1%B;15 min,8%B;35 min,22%B;45 min,42%B;55 min,75%B;65 min,100%B。流速0.2 mL/min; 柱温30 ℃,进样量2 μL,紫外检测波长:300 nm,PAD扫描波长范围190~650 nm。
液质条件:色谱条件同上,质谱条件:离子化方式:电喷雾离子源(ESI),正离子(PI)、负离子(NI)方式;电喷雾离子源参数:喷雾电压:70 V~250 V;扫描区间:m/z100~1 000;氮气流速9.0 L/min,温度350 ℃。
2 结果与分析
2.1 紫外-可见分光光度计光谱分析
2.1.1 类黄酮光谱分析
由钝裂银莲花类黄酮提取液的紫外-可见光光谱分析(图1)可知,在268 nm和330 nm处出现最大吸收峰,符合类黄酮的特征吸收峰。240~285 nm(峰带Ⅱ)为主峰较明显,300~350 nm(峰带Ⅰ)较弱,A-环苯甲酰系统(峰带Ⅱ)和B-环桂皮酰系统(峰带Ⅰ)均含有[8],A-环苯甲酰系统浓度较大。而随着花色加深,类黄酮吸收峰逐渐增大,总含量随花色深浅变化规律性变化(图2),白色较浅黄色最大吸光值分别下降64.03%(268 nm)和67.02%(330 nm),浅黄色较黄色最大吸光值分别下降35.44%(268 nm)和34.78%(330 nm)。

图1 花色素紫外-可见分光光谱分析Fig.1 The UV-visible spectra of petal pigments in Anemone obtusiloba
2.1.2 类胡萝卜素光谱分析
由钝裂银莲花类胡萝卜素提取液的紫外-可见光光谱分析(图1)可知,在268 nm处出现最大吸收峰,符合类胡萝卜素的特征吸收峰,。随花色加深,类胡萝卜素吸收峰逐渐增大,总含量随花色深浅变化规律性变化(图2),白色较浅黄色最大吸光值下降28.19%,浅黄色较黄色最大吸光值下降24.34%。

图2 不同花色花色素的吸光度比较Fig.2 Comparison of petal pigments absorbance of different flower color
类黄酮和类胡萝卜素的总吸光度均随花色变浅而下降,但类黄酮的下降趋势较类胡萝卜素的更明显(趋势线斜率更大),且总含量更高(图2),故类黄酮总含量变化更明显,所以把类黄酮作为钝裂银莲花花色变化的主要花色素定性定量分析 。
2.2 花色素定性定量分析
经过对类黄酮提取物进行HPLC-PAD分析,钝裂银莲花的不同花色在300 nm波长处均可检测到7种化合物,且出峰时间基本一致,表明三种花色所含类黄酮种类一致,根据文献[9]推测其花色的差异性不是由色素种类的不同引起的,而是由某种色素的含量变化导致的(图3)。根据其不同花色同一峰的吸光度变化分析,吸光度的总变化趋势均为随花色变浅而减小,而峰1、2、5、6的化合物在相同花色的吸光度中也随花色变浅而减小,推测其可能是主要色素。
为了进一步确定具体的色素种类,选取总含量较高的样本进行HPLC-MS分析。故选用黄色花的类黄酮提取物进行定性分析。
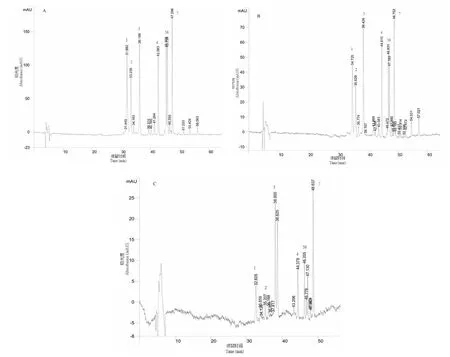
图3 黄色(A)、浅色(B)和白色(C)钝裂银莲花类黄酮的HPLC图谱Fig.3 HPLC chromatogram of flavonids from Anemone obtusiloba yellow(A),pale yellow(B) and white(C)
黄色花类黄酮样品和4个标准品进行HPLC-MS分析,得到图4、图5的总离子流图和图6的样品阳离子质谱图,结合图3,与相关参考文献比较,得出表1的分析结果。

图4 标准品槲皮素(1)、山奈酚(2)、木犀草素(3)和芦丁(4)的总离子流图Fig.4 Total ion chromatogram of standards lquercetin(1),kaempherol(2),luteolin(3) and quercetin-3-O-rutinoside(4)

图5 黄色花类黄酮的总离子流图Fig.5 Total ion chromatogram of flavonid from yellow flower
由图5、图6可知,峰7的 [M+H]+为m/z645.18,碎片离子m/z465为失去一分子葡萄糖所得,再脱去一分子葡萄糖得碎片m/z287,为木犀草素苷元,与文献[10]报道相符合,推测其为木犀草素-3-7-O-葡萄糖苷。峰1、2、3、6的MS2中都有与槲皮素苷元相符的m/z303及其裂解的m/z274碎片,推测其为槲皮素的衍生物。6号峰的 [M]+为m/z625.15,含有碎片离子465和303,说明脱去两分子六碳糖,与文献[11,12]中槲皮素-3-O-芸香糖苷碎片一致。峰3、2的 [M+H]+均为m/z465.1,脱去一分子六碳糖得m/z303,根据文献[15,18]半乳糖苷极性较大,推测峰3为槲皮素-3-O-半乳糖苷,峰2为槲皮素-3-O-葡萄糖苷。峰1的 [M+H]+为m/z449.2,其碎片离子m/z303为脱去一分子鼠李糖,糖基离子为m/z146,与文献[14,17]报道槲皮素-3-O-鼠李糖苷一致。5号峰[M+H]+为m/z611.2,碎片离子m/z287与山奈酚符合,推测其结构中含有山奈酚,碎片m/z445说明失去两分子葡萄糖,与文献[14]报道的山奈酚-3-O-槐糖苷一致。 峰4的 [M+3H]+为m/z483.1,含有杨梅黄素的特征离子m/z318,糖基离子为m/z146,与文献[13,14]的杨梅黄素-3-O-鼠李糖相符合。
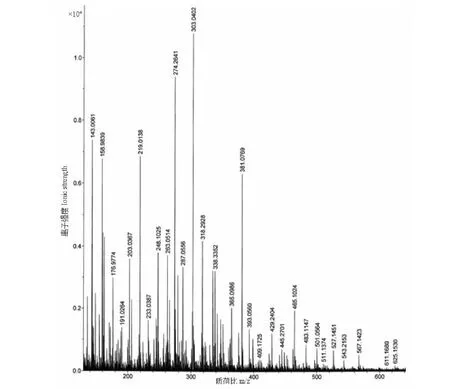
图6 样品的阳离子质谱图Fig.6 Cationic mass spectrograms of sample
根据对各化合物的推测结果及图3的吸光度变化,可以看出槲皮素的糖类衍生物含量占比较大,吸光度随花色变化较明显。
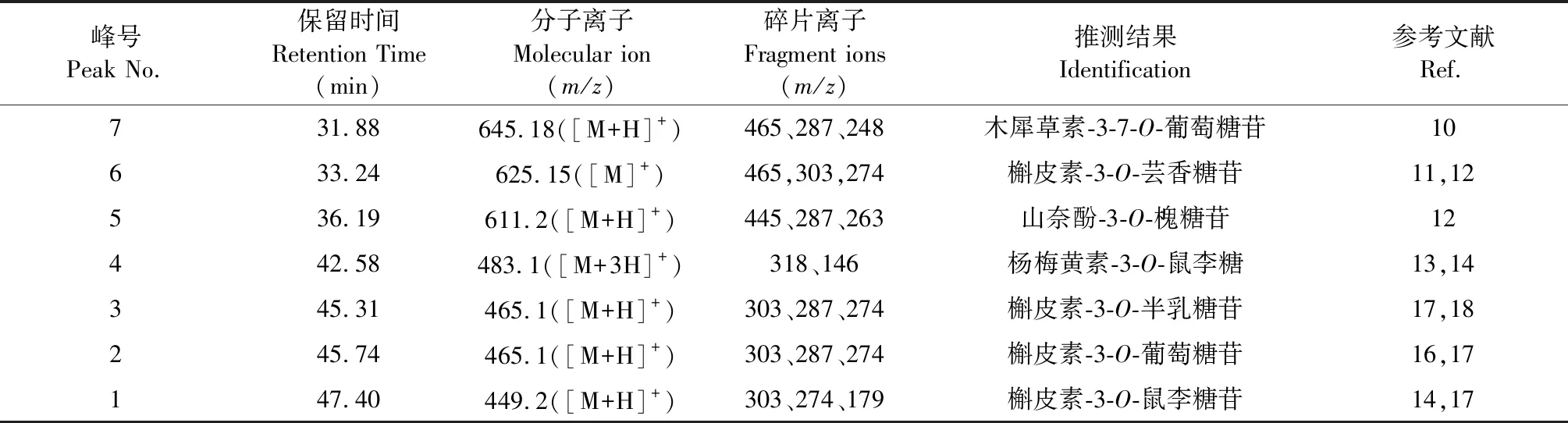
表1 钝裂银莲花黄色花类黄酮HPLC-MS分析结果
3 结论
本研究通过紫外-可见光谱扫描法,HPLC-PAD和HPLC-MS的方法,分析钝裂银莲花花色差异的主要色素及其关键酶。通过对类黄酮的紫外-可见光谱分析,根据谱带Ⅱ和谱带Ⅰ的位置推测,可能含黄酮及其醇类和异黄酮化合物,不含有二氢黄酮及其醇类、橙酮、查尔酮。HPLC-MS测定了类黄酮中的7种色素:木犀草素-3-7-O-葡萄糖苷、槲皮素-3-O-芸香糖苷、山奈酚-3-O-槐糖苷、杨梅黄素-3-O-鼠李糖、槲皮素-3-O-半乳糖苷、槲皮素-3-O-葡萄糖苷、槲皮素-3-O-鼠李糖苷,与紫外-可见光谱结果推测一致。由定性定量结果推测槲皮素的衍生物是导致钝裂银莲花花色差异的主要色素,根据类黄酮代谢途径[19]可以确定合成该色素的酶—类黄酮3′-羟化酶(flavonoid3′-hydroxylase F3′H)是影响槲皮素含量增加的关键酶。
钝裂银莲花生长在环境恶劣的高寒地区,近年来环境变化显著,其白色花逐渐减少,通过本实验,确定了调控其花色深浅的关键酶,推测其花色适应环境的生物机制是通过调控黄酮醇合成酶来影响槲皮素的含量,进而影响槲皮素衍生物的含量,以达到改变花色深浅适应环境的目的。本实验为进一步研究钝裂银莲花的进化机制提供了有力的数据参考,对其花色的研究有比较有意义的价值。
猜你喜欢
家庭医药(2021年18期)2021-12-03
保健与生活(2021年13期)2021-07-14
医药前沿(2020年36期)2020-12-03
北方药学(2020年12期)2020-04-30
中华肺部疾病杂志(电子版)(2020年1期)2020-01-07
中国药理学与毒理学杂志(2019年4期)2019-08-12
恋爱婚姻家庭(2019年30期)2019-03-23
福建茶叶(2019年10期)2019-01-07
中成药(2017年9期)2017-12-19
中南林业科技大学学报(2017年12期)2017-12-19
