
干酪乳杆菌胞外多糖诱导小鼠骨髓来源树突细胞成熟以及分泌IL-6、TGF-β和IL-23
2019-04-02 03:42任琦琦任皓威杨翠翠
食品科学 2019年5期
任琦琦,任皓威,杨翠翠,刘 宁*
(东北农业大学食品学院,乳品科学教育部重点实验室,黑龙江 哈尔滨 150030)
乳酸菌胞外多糖(lactic acid bacteria-derived exopolysaccharide,LAB EPS)是乳酸菌发酵过程中的产物之一,在改善发酵乳制品的质构特性、流变学特性和风味方面具有重要作用[1],也在生物活性方面发挥着重要作用。一些研究者已经报道了LAB EPS的抗肿瘤[2]、抗氧化[3]、调节消化道[4]、免疫调节活性[5]等多种生物学活性。干酪乳杆菌(Lactobacillus casei)作为乳酸菌中的一种,被广泛应用在乳制品发酵中。已有研究表明,干酪乳杆菌对人体健康的潜在益处包括清除有害菌[6]、维持肠道菌群多样性[7]、调节免疫系统[8]、降低胆固醇水平[9]、抗氧化和抗炎作用[10]等,而EPS作为L. casei发酵过程中的产物之一,在防止癌细胞增殖[11]、抗细胞毒性[12]等生物活性方面发挥着重要作用。
近年来的研究表明,益生菌发酵乳长时间干预机体可以提高肠黏膜免疫系统中树突细胞(dendritic cells,DCs)的数量[13-14],而EPS作为发酵产品中重要的生物活性成分,可能发挥一定作用。另据报道,多糖能够刺激小鼠骨髓来源树突细胞(bone marrow-derived dendritic cells,BMDCs)的体外成熟[15-18]。DCs作为抗原提呈细胞(antigen-presenting cell,APC),其功能是目前所知最强大的,它能够引发并调节免疫反应[19]。DCs在摄取抗原和活化T细胞的过程中,未成熟DCs向成熟DCs转化,并且其功能与成熟相关。DCs成熟过程中发生了一系列的变化,如形态发生改变、MHC II类分子和共刺激分子CD86等的高表达,而未成熟的DC只表达很少量的MHC II和CD86[20]以及分泌影响初始型T细胞分化方向的细胞因子[21-22]。对于L. casei EPS在促进未成熟DCs向成熟DCs转化方面的研究鲜见报道,也鲜见其诱导成熟后的DCs分泌与T细胞向辅助性T细胞(helper T cell,Th)17分化增殖相关的细胞因子的研究报道。
本研究通过流式细胞法检测L. casei EPS处理后体外培养的小鼠未成熟BMDCs表面表达MHC II和CD86的情况,研究L. casei EPS对体外培养的小鼠BMDCs促成熟的作用,用酶联免疫吸附检测(enzyme-linked immunosorbent assay,ELISA)法分析不同剂量的EPS对BMDCs分泌与Th17细胞分化和增殖相关的细胞因子白细胞介素(interleukin,IL)-6、转化生长因子-β(transforming growth factor-β,TGF-β)和IL-23的影响,为进一步探讨L. casei EPS通过影响DCs分泌细胞因子来影响初始型T细胞的分化方向提供依据。最后,用CCK-8法检测混合培养体系中异基因淋巴细胞的增殖率,研究L. casei EPS对BMDCs抗原提呈能力的影响。
1 材料与方法
1.1 动物、材料与试剂
BALB/c小鼠(使用许可证号:SYXK(沪)2017-0014),6~8 周龄,雌性,体质量18~22 g,清洁级,由北京维通利华公司提供。
干酪乳杆菌(编号为ATCC393TM),购自美国种质保藏中心。
RPMI 1640完全培养基、胎牛血清、磷酸盐缓冲液(phosphate buffered saline,PBS) 美国Gibco公司;重组小鼠粒细胞巨噬细胞集落刺激因子细胞因子(recombinant mouse granulocyte macrophagecolony stimulating factor,rmGM-CSF)、重组小鼠IL-4(recombinant mouse IL-4,rmIL-4)、小鼠细胞因子ELISA试剂盒(IL-6、TGF-β和IL-23) 美国R & D Systems公司;脂多糖(lipopolysaccharide,LPS)美国Sigma公司;荧光单克隆抗体(PE-CD11c、FITC-MHC II和FITC-CD86) 美国eBioscience公司;CCK-8试剂盒 合肥Biosharp公司。
1.2 仪器与设备
高压灭菌锅 美国ZEALWAY公司;ZFQ85B旋转蒸发仪 上海医械专机厂;真空冷冻干燥机 德国Christ公司;BCD-518WSA型低温冰箱 青岛海尔集团;EPICS XL流式细胞仪 美国贝克曼库尔特有限公司;Model 680酶标仪 美国Bio-Rad公司。
1.3 方法
1.3.1 L. casei EPS的制备
干酪乳杆菌按3%接种量,接种在脱脂乳培养基上,32 ℃培养26 h后,将发酵液置于沸水浴中煮沸10 min,10 000×g、4 ℃离心20 min,去除细菌和变性蛋白沉淀。向发酵液上清液中加入三氯乙酸至终质量浓度为4 g/100 mL ,4 ℃下搅拌12 h后,10 000×g、4 ℃离心20 min,去除变性蛋白,收集上清液。使用旋转蒸发仪在0.9 kPa和65 ℃条件下将上清液浓缩至原体积的1/4。向浓缩上清液加入2 倍体积的冷无水乙醇,在4 ℃下静置12 h后,离心去上清液,得到乙醇沉淀后的多糖。将收集到的多糖产物转移到透析袋(分子质量7 000~14 000 Da)内,将透析袋浸入到去离子水中,每8 h换水一次,持续3 d。将最终获得的EPS溶液进行冷冻干燥,获得冻干粉。
1.3.2 L. casei EPS中糖含量的检测
EPS样品中的糖含量用苯酚-硫酸法[23]测定。以标准葡萄糖浓度为横坐标,490 nm波长处测定的OD值为纵坐标绘制葡萄糖标准曲线。L. casei EPS样品的测定方法同标准曲线测定,称取0.1 mg EPS冻干粉于25 mL比色管中,准确加入蒸馏水至1.0 mL,之后加入1.0 mL 5%苯酚溶液及2.5 mL浓硫酸,摇匀,室温静置10 min,30 ℃水浴20 min后于室温下490 nm波长处测定OD值。EPS样品中的蛋白质和核酸含量用分光光度计分别在280、260 nm波长处检测。
1.3.3 骨髓来源树突细胞的诱导
骨髓来源树突细胞(bone marrow-derived dendritic cells,BMDCs)的诱导参照Inaba等[24]的方法并稍加改进。脱颈处死6~8 周的BALB/c小鼠,在体积分数75%乙醇溶液中浸泡2 min后,将其置于超净台的木板(已消毒)上固定,剔除后腿肌肉并剪下股骨和胫骨。在无菌条件下,用1 mL注射器吸取RPMI 1640完全培养基将骨髓细胞从胫骨和股骨中冲洗出,并将其打成单细胞悬液,300×g离心5 min,得到细胞沉淀。用红细胞裂解液裂解血红细胞,冰敷5 min,300×g离心5 min。用PBS洗涤细胞2 次后,将骨髓单细胞重悬在含10%胎牛血清的RPMI 1640培养液中,并向培养液中添加rmGM-CSF和rmIL-4,使终质量浓度均为20 ng/mL,在37 ℃、5% CO2的培养箱中培养6 d。在为期6 d的培养过程中,每2 d半量吸除培养液,丢弃未贴壁的细胞,然后补充含有rmGM-CSF和rmIL-4的新鲜培养基,由于未成熟DCs具有半贴壁和趋于悬浮的特性,所以在第6天收集疏松贴壁细胞即为未成熟BMDCs。
1.3.4 BMDCs的鉴定
将诱导培养6 d后的BMDCs在4 ℃用体积分数10%的羊血清封闭液封闭细胞表面Fc受体10 min后,细胞离心并悬浮于预冷PBS中,调整细胞浓度为1.0×106个/mL,并加入FITC标记的仓鼠抗小鼠CD11c单克隆抗体FITC-CD11c,振荡均匀,孵育30 min,所有操作都在冰上和黑暗中进行。标记后的细胞用预冷PBS洗涤两次,再用预冷PBS重悬,用流式细胞仪检测。
1.3.5 EPS处理的BMDCs表面标记物CD86和MHC II的检测
松散附着的未成熟BMDCs用无菌移液管收集,300×g离心5 min后,以细胞浓度1.0×105个/mL接种到24 孔板中,每孔中含有添加了20 ng/mL rmGM-CSF和20 ng/mL rmIL-4的1 mL 10%胎牛血清的RPMI 1640培养液。将BMDCs分为PBS组(阴性对照组)、EPS组(实验组,EPS终质量浓度100 μg/mL)和LPS组(阳性对照组,LPS终质量浓度1 μg/mL)。将24 孔板放入37 ℃、5% CO2培养箱中孵育24 h。
孵育24 h后的细胞在冷PBS中洗涤2 次后,放在一个圆底96 孔板中,孔中含有体积分数10%山羊血清封闭液。4 ℃交互10 min后,细胞离心并悬浮于PBS,调整细胞浓度为1×105个/mL,加入PE-CD11c、FITC-CD86和FITC-MHC II,孵育30 min。抗体标记的所有步骤都在冰上和黑暗中进行。标记后的细胞用PBS洗涤2 次,再用预冷PBS重悬,用于流式细胞仪检测。
1.3.6 EPS处理的BMDCs分泌IL-6、TGF-β和IL-23的检测
未成熟BMDCs用添加了20 ng/mL rmGM-CSF和20 ng/mL rmIL-4的含有10%胎牛血清的RPMI 1640培养液悬浮,以浓度1.0×105个/mL接种到24 孔板中。将BMDCs分为PBS组(阴性对照组)、EPS组(实验组,EPS终质量浓度分别为12.5、25、50、100、200 μg/mL)和LPS组(阳性对照组,LPS终质量浓度1 μg/mL)。24 孔板在37 ℃和5% CO2中孵育24 h。
将孵育后的24 孔板中的细胞培养液转移到无菌管中,300×g离心20 min后,收集上清液用于细胞因子的检测。如未及时检测,将细胞上清液于-80 ℃冻存直至使用。细胞因子IL-6、TGF-β和IL-23的浓度检测采用ELISA试剂盒,具体操作步骤参照试剂盒使用说明书。反应终止后,使用酶标仪在450 nm波长处检测OD值,再根据标准曲线确定细胞因子的浓度。
1.3.7 EPS对淋巴细胞增殖影响的测定
脱颈处死BALB/c小鼠,无菌条件下取脾,制备成脾单细胞悬液,用红细胞裂解液裂解血红细胞后,加入淋巴细胞分离液,300×g离心30 min,将分离出的脾淋巴细胞重悬在RPMI 1640培养基中,0.4%台盼蓝检测细胞活力,当细胞存活率大于95%时,调整细胞密度至2.0×105个/mL,获得用于接种的淋巴细胞悬液。向1.3.3节分离诱导出的未成熟BMDCs中加入丝裂霉素C以去除细胞增殖活性,调整经处理的BMDCs细胞浓度至2.0×105个/mL,作为刺激细胞和淋巴细胞悬液共同接种于96 孔板中,细胞接种量为每孔50 μL BMDCs和50 μL淋巴细胞悬液。将接种的细胞混合后,分为PBS组(阴性对照组)、不同剂量的EPS组(实验组,EPS终质量浓度分别为12.5、25、50、100、200 μg/mL)和LPS组(阳性对照组,LPS终质量浓度1 μg/mL),每组设6 个复孔。96 孔板在37 ℃、5% CO2的培养箱中孵育72 h后,向每孔中加入20 μL CCK-8溶液,在37 ℃、体积分数5% CO2的培养箱中继续孵育2 h,使用酶标仪在450 nm波长处检测OD值,按下式计算淋巴细胞增殖率。

1.4 数据统计与分析
2 结果与分析
2.1 多糖纯度的测定分析
葡萄糖标准曲线的回归方程为y=-0.016 6+8.92x(R2=0.999 54)。490 nm波长处测得0.1 mg/mL EPS样品溶液的OD值,将其代入葡萄糖标准曲线回归方程中,得出0.1 mg/mL EPS样品溶液中总糖质量浓度为(0.095 02±0.001 46)mg/mL,可以推算出1 mg EPS冻干粉中的总糖质量为(0.950 2±0.014 6)mg,即L. casei EPS纯度可达95%。紫外分光光度计检测EPS中蛋白质和核酸含量,发现在280、260 nm波长处没有吸收,表明EPS样品中蛋白质、核酸的含量极少,说明纯化的EPS在研究中的活性作用不受蛋白质和核酸的影响。
2.2 诱导出BMDCs的鉴定
CD11c是小鼠DCs的特征性表面分子,通过检测CD11c的表达量,可以间接反映出诱导的BMDCs的数量。由图1可知,流式细胞仪分析出的表达CD11c的细胞数量约占80%,表明通过骨髓造血干细胞分化出的未成熟DCs数量约占80%。
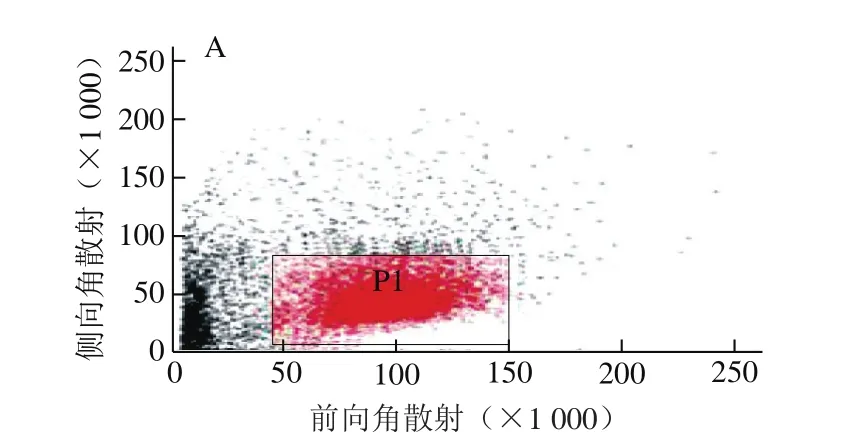
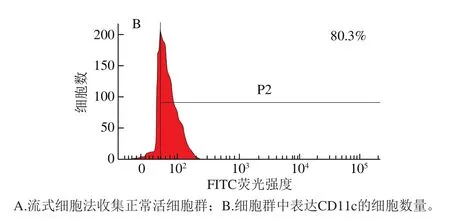
图 1 表达CD11c的BMDCs细胞数量Fig. 1 Number of BMDCs expressing CD11c
2.3 EPS对BMDCs成熟的影响
成熟的DCs表面能表达大量的MHC II类分子和共刺激分子,如CD86,而未成熟DCs只能少量表达这些分子。MHC II类分子和共刺激分子CD86是DCs识别和摄取抗原必要的信号转导,也是DC免疫功能的最佳指标。
2.3.1 EPS对BMDCs表达MHC II的影响
如图2所示,PBS组的DCs表面MHC II类分子的表达量为63.8%,而100 μg/mL EPS组和LPS组的表达量分别为68.1%和70.2%。结果表明,EPS能显著增加DCs表面MHC II分子的表达(P<0.05),并且EPS组的MHC II分子表达量低于LPS组。证明EPS能够促进BMDCs表面表达更多的MHC II类分子,EPS对BMDCs的成熟有促进作用。并且DCs刺激初始型T细胞活化和增殖的一个信号是通过MHC-抗原肽复合物和TCR的相互作用来传递的[25],因此可以推断出,EPS对于DCs刺激初始T细胞活化和增殖也有相应的促进作用。


图 2 流式细胞术分析分别用PBS(A)、EPS(B)和LPS(C)处理后的BMDCs表面MHC II的表达量(D)Fig. 2 Expression of MHC II (D) on the surface of BMDCs treated with PBS (A), EPS (B) or LPS (C) evaluated by fl ow cytometer
2.3.2 EPS对BMDCs表达CD86的影响

图 3 流式细胞术分析分别用PBS(A)、EPS(B)和LPS(C)处理后的BMDCs表面CD86的表达量(D)Fig. 3 Expression of CD86 (D) on the surface of BMDCs treated with PBS (A), EPS (B) or LPS (C) evaluated by fl ow cytometer
如图3所示,PBS组的DCs表面共刺激分子CD86的表达量为68.5%,而100 μg/mL EPS组和LPS组的表达量分别为74.3%和75.1%。结果表明,EPS能提高DCs表面共刺激分子CD86的表达(P<0.05),并且EPS组CD86的表达量低于LPS组。证明EPS能够促进BMDCs表面表达更多的CD86分子,EPS对BMDCs的成熟有促进作用。DCs刺激初始T细胞活化和增殖的一个信号是共刺激信号,通过DCs上的CD80/CD86和T细胞上的CD28的相互作用来传递[23],因此可以推测EPS对于DCs刺激初始T细胞活化和增殖也有相应的促进作用。
2.4 EPS对DCs分泌与Th17细胞分化增殖相关的细胞因子的影响
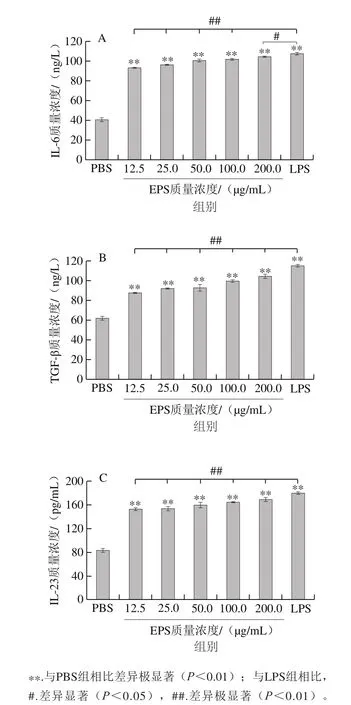
图 4 EPS对DCs分泌与Th17细胞分化增殖相关的细胞因子IL-6(A)、TGF-β(B)和IL-23(C)的影响Fig. 4 Effect of EPS on cytokine secretion of DCs, IL-6 (A), TGF-beta (B)and IL-23 (C) associated with differentiation and proliferation of Th17 cells
在小鼠模型中,促炎细胞因子IL-6和TGF-β能促进Th17细胞分化,IL-6和TGF-β可作为诱导CD4+T细胞分化为Th17细胞的起始因子[26]。许多研究表明,IL-23的缺乏并不影响T细胞向Th17细胞的分化,这意味着IL-23不在初始CD4+T细胞发挥作用,它只是促进Th17细胞的增殖,所以认为IL-23是Th17细胞生存和增殖的关键因素[27]。
由图4A可知,当EPS质量浓度为12.5~200 μg/mL时,均能够显著促进BMDCs表达IL-6,EPS组的IL-6表达量分别为(93.18±0.62)、(96.21±0.62)、(100.51±1.56)、(101.77±0.94)、(104.55±0.62)ng/L,与PBS组表达量(40.66±1.89)ng/L相比具有极显著性差异(P<0.01),并且IL-6表达量与EPS的质量浓度呈正相关,当EPS质量浓度为200 μg/mL时,IL-6的表达量最多。值得注意的是,EPS组与LPS组((107.50±1.24)ng/L)相比,IL-6的表达量显著减少(P<0.05)。
由图4B可知,当EPS质量浓度为12.5~200 μg/mL时,均能够极显著促进B M D C s分泌T G F-β,EPS组的TGF-β表达量分别为(87.56±1.00)、(91.63±0.57)、(92.85±2.87)、(99.35±1.52)、(104.23±2.51)ng/L,各EPS组表达量与PBS组((61.95±1.99)ng/L)相比均有极显著差异(P<0.01),并且EPS质量浓度与促进BMDCs分泌TGF-β的量呈正相关,当EPS质量浓度为200 μg/mL时,TGF-β的表达量最多。LPS组TGF-β表达量为(115.05±1.49)ng/L,EPS组与之相比,TGF-β的表达量极显著减少(P<0.01)。
由图4C可知,当EPS质量浓度为12.5~200 μg/mL时,均能够极显著促进B M D C s分泌I L-2 3,EPS组IL-23表达量分别为(152.22±2.58)、(1 5 2.7 3±3.7 1)、(1 5 9.3 0±3.7 8)、(163.84±1.43)、(168.38±3.78)pg/mL,与PBS组的表达量(82.53±3.78)pg/mL相比差异极显著(P<0.01),并且IL-23表达量与EPS质量浓度呈正相关,当EPS质量浓度为200 μg/mL时,IL-23的表达量最多。值得注意的是,EPS组与LPS组IL-23的分泌量(179.22±1.20)pg/mL相比,IL-23的表达量极显著减少(P<0.01)。
以上结果表明,一定质量浓度的EPS能够促进DCs分泌IL-6、TGF-β和IL-23,并且呈剂量依赖性,说明EPS可以通过促进DCs分泌IL-6和TGF-β来促进初始T细胞向Th17细胞方向分化,分泌IL-23促进Th17细胞的增殖。
2.5 EPS影响BMDCs对异基因淋巴细胞增殖的刺激作用
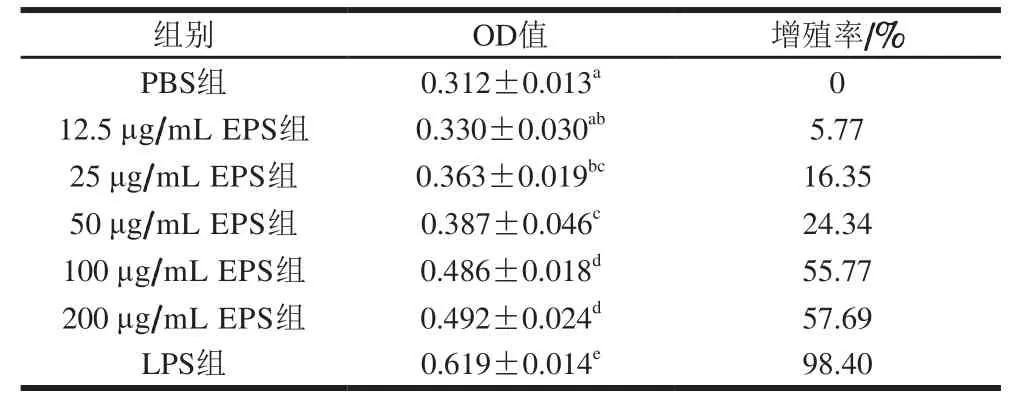
表 1 EPS对脾淋巴细胞增殖的影响Table 1 Effect of EPS on the proliferation of mouse splenic lymphocytes
EPS通过BMDCs的刺激诱导异基因淋巴细胞的增殖,进而能够反映EPS对BMDCs抗原提呈能力的影响。如表1所示,EPS质量浓度为25~200 μg/mL实验组的淋巴细胞增殖率与阴性对照组相比显著增多(P<0.05),与阳性对照组相比显著减少(P<0.05),表明EPS提高了异基因淋巴细胞的增殖能力,说明EPS能够增加BMDCs的抗原提呈能力。
实验组的淋巴细胞增殖率随着EPS质量浓度的增加呈上升趋势,但是50 μg/mL EPS与25 μg/mL EPS组的淋巴细胞增殖率相比未显著增加,25 μg/mL EPS与12.5 μg/mL EPS组的淋巴细胞增殖率相比未显著增加,而100 μg/mL EPS和200 μg/mL EPS组与其他实验组的淋巴细胞增殖率相比增加显著(P<0.05),但是这两个实验组的淋巴细胞增殖率相比没有显著性差异,表明100、200 μg/mL EPS更有利于淋巴细胞增殖,且促进效果相当。
3 讨 论
DCs发挥抗原提呈功能和启动体外免疫调节与其是否成熟密切相关,MHC II分子和CD86是成熟DCs的标志物,也是活化T细胞的两个必要因素,在激发免疫反应中起着不可或缺的作用[25]。本研究结果表明,L. casei EPS能够促进DCs表面MHC II分子和CD86的表达,说明EPS可能导致DCs的成熟,也可以进一步推测EPS可能通过增强DCs抗原呈递能力来促进T细胞的活化,进而影响T细胞的分化和增殖。
在2005年,Th17细胞作为区别于Th1和Th2型细胞的第3种效应性Th细胞亚群被发现,Th17细胞在组织炎症和自身免疫性疾病发生的过程中起到重要的调节作用[28-29]。DCs可以通过分泌不同的细胞因子来介导T细胞的活化,其中DCs分泌的IL-6、TGF-β和IL-23可以诱导T细胞向Th17细胞的分化和增殖[21-22]。本研究结果表明,L. casei EPS能够促进DCs分泌IL-6、TGF-β和IL-23,说明EPS可能通过促进DCs分泌与向Th17细胞分化相关的必要细胞因子来诱导T细胞向Th17的分化和增殖。而DCs通过分泌不同细胞因子来介导T细胞分化的方向是DCs成熟之后功能性的表达之一,进一步验证出EPS对于DCs成熟的促进作用。
DCs刺激淋巴细胞增殖可以反映出其抗原提呈能力,DCs的提呈抗原能力随其发育成熟而逐步增强,因此,可以通过淋巴细胞的增殖率来反映出DCs的成熟状态[30]。本研究结果表明,L. casei EPS能够使BMDCs对异基因淋巴细胞的增殖产生明显刺激作用,进一步验证出EPS能够增强BMDCs的抗原提呈能力,并能诱导其成熟,同时刺激淋巴细胞增殖能力增强,进而影响细胞免疫。
LPS是一种常见的内毒素,它能够使细胞产生强烈的免疫反应或损害,通常在研究中设计为阳性对照[15-16]。本研究结果表明,干酪乳杆菌EPS对BMDCs表面MHC II分子和CD86以及分泌的细胞因子的表达量的影响均比LPS较小,并且对BMDCs刺激淋巴细胞增殖的能力小于LPS,说明EPS能够适度地调节DCs免疫反应。
综上所述,在体外实验中,L. casei EPS对BMDCs具有诱导成熟的效应活性,能够增加BMDCs表面MHC II分子和CD86的表达,促进BMDCs分泌与Th17分化和增殖相关的细胞因子IL-6、TGF-β和IL-23,并且可以促进BMDCs的抗原提呈能力,刺激淋巴细胞增殖,但是L. casei EPS通过影响DCs分泌细胞因子来影响初始T细胞的分化方向,以及涉及的相关信号转导通路与分子机制还有待进一步验证。
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
清华金融评论(2022年4期)2022-04-13
现代临床医学(2021年4期)2021-07-31
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
中西医结合心脑血管病杂志(2016年20期)2016-03-01
医学研究杂志(2015年12期)2015-06-10
医学研究杂志(2015年3期)2015-06-10
中国药理学通报(2014年2期)2014-05-09
中国医学科学院学报(2013年2期)2013-03-11
