
景宁荒田湖林场黄山松与木荷混交林主要树种径级结构与空间分布格局研究
2019-03-17 07:31许元科吴初平叶丽敏侯建花袁位高朱锦茹黄玉洁焦洁洁王志高
浙江林业科技 2019年5期
许元科 ,吴初平,叶丽敏,侯建花,江 波,袁位高,朱锦茹,黄玉洁,焦洁洁,王志高
(1.景宁畲族自治县生态林业发展中心,浙江 景宁,323500;2.浙江省林业科学研究院,浙江 杭州,310023;3.景宁畲族自治县自然资源和规划局,浙江 景宁,323500)
森林群落中,群落优势种一般个体数相对较多,体积相对较大,对非优势种具有不对称性竞争优势,通过改变林下环境(光照、水分和土壤等)影响物种分布[1]。群落优势种年龄结构和空间分布格局不仅反映了不同个体的空间配置情况和数量动态及发展趋势,也在很大程度上反映了种群与环境的相互关系及其在群落中的作用和地位。分析优势种的径级结构和空间分布格局,对于了解群落的结构和动态以及进一步对森林资源进行管理具有重要的意义。
种群分布格局是种群的生物学特性以及各种生物因素和非生物因素综合作用的结果。研究表明,由于种子扩散的限制和生物学特性以及生境异质性等因素的作用,森林中绝大多数树种呈聚集型分布[2-3]。群落优势树种的分布格局反映了其在群落中的生长状态和资源利用状况,但是物种的生态需求随生活史发育阶段而变化[4]。如对浙江古田山中亚热带常绿阔叶林两个优势树种甜槠Castanopsis eyrei 和木荷Schima superba 的研究表明其在空间上呈现聚集分布的特征,但聚集强度随年龄或径级的增加有下降的趋势[5];对江西九连山鹿角锥C.lamontii,米槠C.carlesii,罗浮柿Diospyros morrisiana 和细枝柃Eurya loquaiana 的研究也得出类似的结果,但是优势种具有明显的生境偏好[6]。对江西井冈山重要值排前4 位的优势物种马银花Rhododendron ovatum,甜槠,黄丹木姜子Litsea longate 和石木姜子L.longate var.faberi 的研究发现,在检测的大部分尺度上,马银花、黄丹木姜子和石木姜子呈显著聚集分布,而甜槠则呈随机分布[7]。因此资源利用方式的差异是导致群落优势树种空间分布格局差异的主要因素。
针叶树种和阔叶树种对资源的利用方式存在差异[8],黄山松Pinus taiwanensis 和木荷混交林是亚热带中山地森林中常见的一种林型,木荷和黄山松共存于群落上层。对长白山阔叶红松林中紫椴Tilia amurensis 和红松P.koraiensis 空间分布格局的研究表明,两个树种在空间上总体呈聚集分布,但物种在较低的林层呈明显的聚集分布,而在较高的林层则呈随机或规则分布,聚集度随林层的增高而降低[9]。前期的研究对不同演替阶段的黄山松和木荷混交林的空间分布格局关注较多,如研究表明黄山松一般表现出聚集分布的特点,而在成熟林中主要为随机型或聚集型分布[10-11]。木荷种群在黄山松幼林中为随机分布,在木荷+黄山松混交阶段为聚集型分布[12]。对黄山松和木荷混交林中不同生长阶段的林木空间分布格局及其尺度变化特征的关注较少,而不同生长阶段的树种分布状态是森林管理措施制定的主要依据之一。
本文以浙江景宁中山地带的黄山松和木荷混交林为研究对象,分析其径级结构特征和不同生活史阶段的空间分布格局及其尺度依赖性特征,研究资源利用方式和尺度对其空间分布格局的影响,为黄山松和木荷混交林管理提供科学依据。
1 研究区域概况
研究区域位于浙江省景宁畲族自治县林业总场荒田湖分场,119°41′45"~119°43′34"E,27°55′36"~27°58′27"N。荒田湖分场是景宁大仰湖自然保护区的一部分。属中亚热带海洋性季风气候,年平均气温12.8℃,极端最低气温-14.0℃。无霜期约19.6 d。年日照时数1 617.6 h,年降水量1 918 mm,年平均相对湿度80%以上,自然气热条件具有温暖湿润、雨量充沛、四季分明、热量资源丰富的特点。地貌属于浙南中山区,海拔613.8~1 556.9 m,以深切割山地为主。保护区共有维管植物1 867种,具有伯乐树Bretschneidera sinensis,南方红豆杉Taxus wallichiana var.mairei,蛇足石杉Huperzia serrata 等38种国家及省级重点保护野生植物。保护区自然环境多样,植物资源丰富、种类多样、分布广泛,具有良好的资源优势和潜在的经济优势。
2 研究方法
2.1 样地设置
在浙江省景宁县荒田湖林场的黄山松、木荷混交林中坡面位置相对一致、干扰较少的典型地段建立3个50 m×50 m 森林固定监测样地,样地内按10 m×10 m 划分样方进行调查。记录样地中胸径(DBH)1 cm 以上乔木层树种,DBH≥5 cm 的树种名称、位置(坐标)和胸径等因子,并记录样地海拔、坡度和坡向等立地因子。
2.2 统计分析
采用物种丰富度、多度、Shannon-Weiner 指数和Pielou 均匀度指数度量群落结构特征,其中物种丰富度即物种数目,S;多度即林木个体数目,N;其余指数计算公式如下[13-14]:Shannon-Weiner 指数(H′)计算:

Pielou 均匀度指数(J′)计算:

上述两个公式中,Pi为第i个物种的多度占所有物种多度之和的比例,S 为物种总数。
重要值计算:

本研究用树木的胸高断面积表示优势度。采用每5 cm 为一个径级分别对黄山松和木荷的径级进行划分,统计每个径级的个体数目。采用Ripley’s G单变量函数(G 函数)和对物种空间分布格局特征进行分析,采用在完全随机零模型下进行99 次Monte Carlo 检验,计算上、下包迹线(envelopes),即99%置信区间。用种群实际分布数据计算不同尺度上的G(r)值,在单变量分析中G(r)在上、下包迹线之间为随机分布,高于上包迹线为聚集性分布,低于下包迹线则为规则分布。点格局分析通过R3.5.2 软件中的spatstat 软件包完成[15]。
3. 结果与分析
3.1 乔木层群落结构与主要树种径级结构
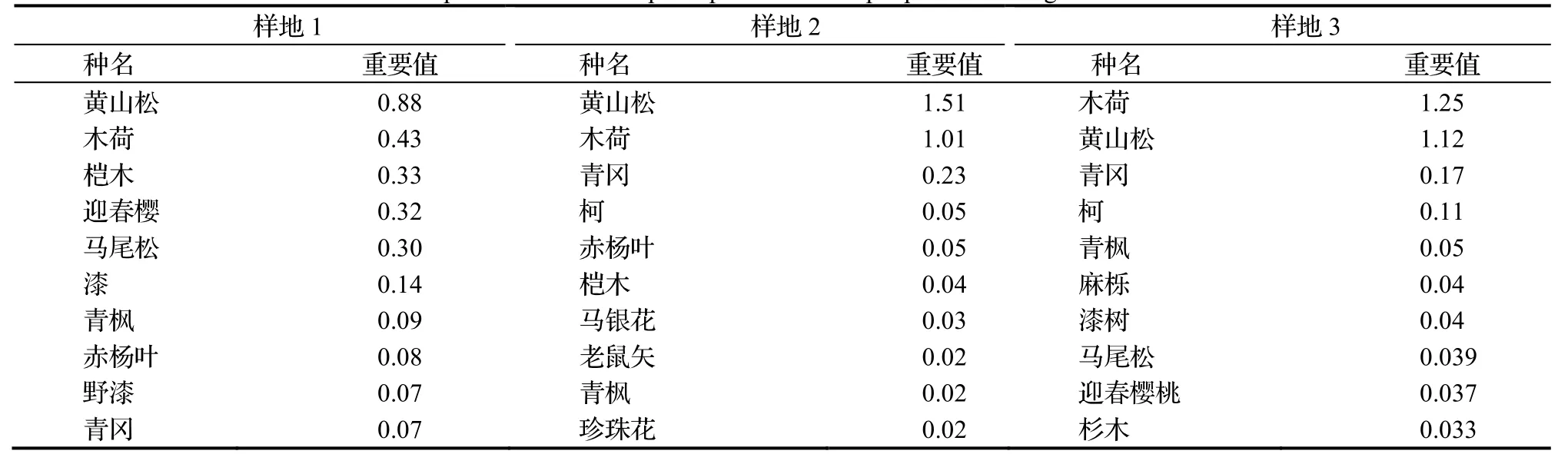
表1 荒田湖林场3个样地重要值前十位物种Table 1 Importance value of top 10 species in 3 sample plots of Huang tian hu forest farm
由表1 可知,3个样地中黄山松和木荷种群的个体数均占优势,其中黄山松的个体数在样地1,2 和3 中占比分别为21.9%,49%和33.4%;木荷分别为16.7%,40.7%和52.4%,仅样地3 中的木荷种群数量多于黄山松种群。样地1 与样地2、样地1 与样地3、样地2 与样地3 的物种相似度分别为40.7%,46.4%和52.6%,重要值排名前十的物种相似度分别为33.3%,42.9%和42.9%,其中样地2 和样地3 的群落结构相似度相对较高。对3个样地中的物种数目(物种丰富度)、林木株数(多度)、Shannon-Weiner 多样性指数和Pielou 均匀度指数的分析发现,四个指标表现出一致的结果(表2),即样地2 的物种多样性最低,但是密度最大,样地1 物种多样性最高最复杂,样地2 的群落结构最为简单。黄山松种群的径级结构在3个样地中均呈正态分布。木荷种群为偏正态性分布,径级>5~10 cm 的个体数目最多,大径级个体数较少(图1)。
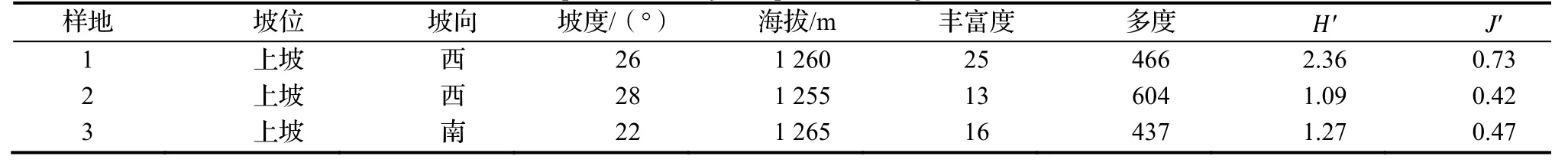
表2 荒田湖林场3个样地多样性指标Table 2 Species diversity of 3 plots in Huangtianhu Forest Farm

图1 木荷和黄山松种群径级结构Figure 1 Size class distributions of Schima superba and Pinus taiwanensis
3.2 空间分布格局
3个样地中,黄山松和木荷种群的大树和幼树分布在样地不同位置(图2A,图3A),表明其在空间分布上存在分离的现象。木荷种群在不划分和划分径级的情况下,在3个样地中均表现出小尺度上的聚集分布和较大尺度上为随机分布格局(图2B,图3B)。
黄山松种群空间分布格局在3个样地间差异明显,在不划分幼树和大树的情况下,样地1 和样地3 在不同尺度下均为随机分布,而样地2 在小尺度上为均匀分布,较大尺度上呈现随机分布。划分为幼树和大树后,样地1 幼树和大树为随机分布;而样地2 的幼树所有尺度上随机分布,大树则在小尺度上为均匀分布;样地3 的幼树在小尺度上为聚集分布,大树均为随机分布(图2C,图2D,图3C,图3D)。
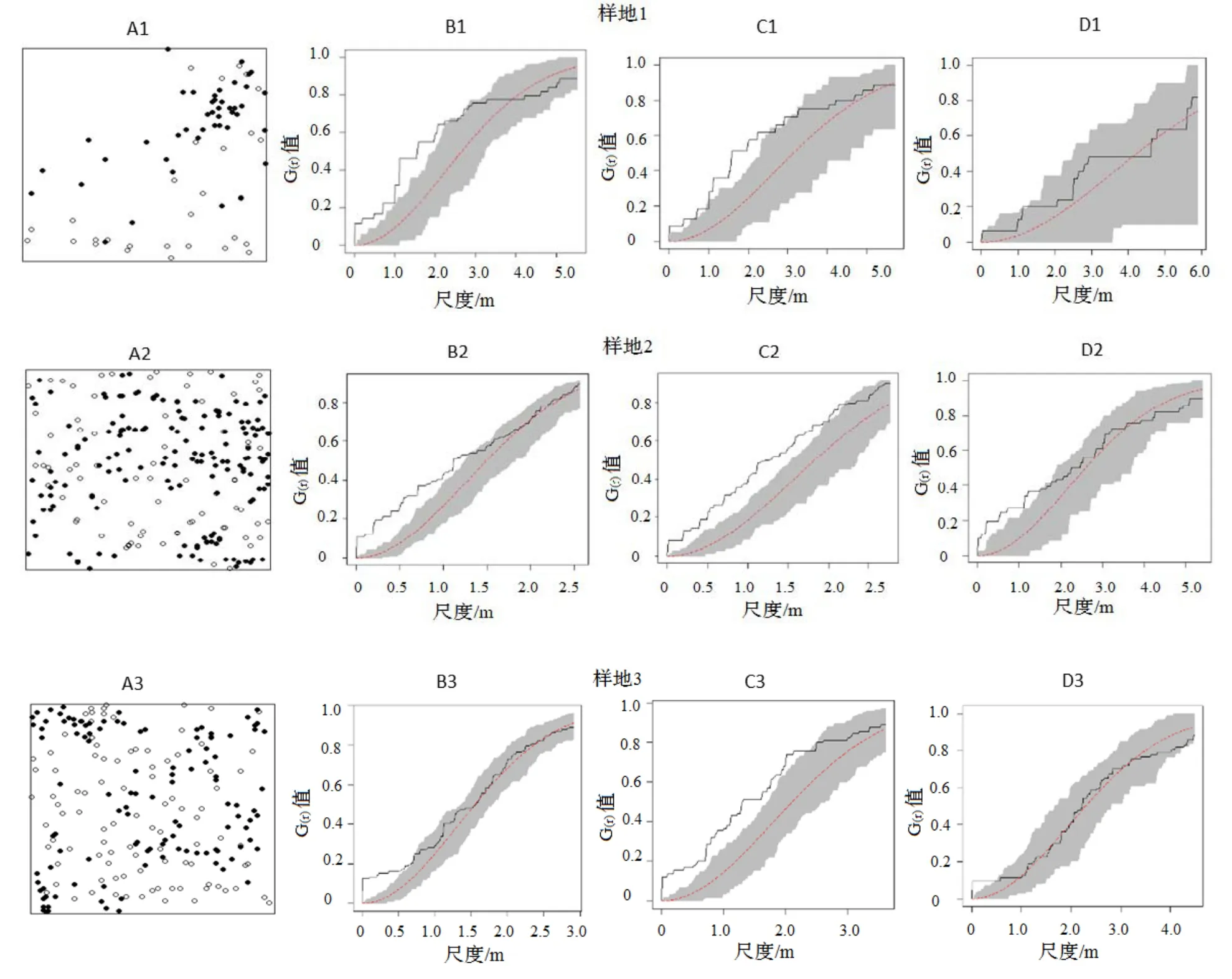
图2 木荷种群分布格局Figure 2 Spatial patterns of Schima superba
4 结论与讨论
以上研究结果表明,荒田湖林场3个50 m×50 m 针阔混交林样地群落结构存在差异,样地1 的物种多样性相对较高,样地2 和样地3 的群落结构较为相似,物种多样性相对较低。3个样地中,黄山松种群径级结构呈正态分布,结构稳定,而木荷种群的径级结构呈偏正态分布,低龄级个体相对较多,种群结构呈增长型,显示木荷种群正处于入侵黄山松的阶段[6]。
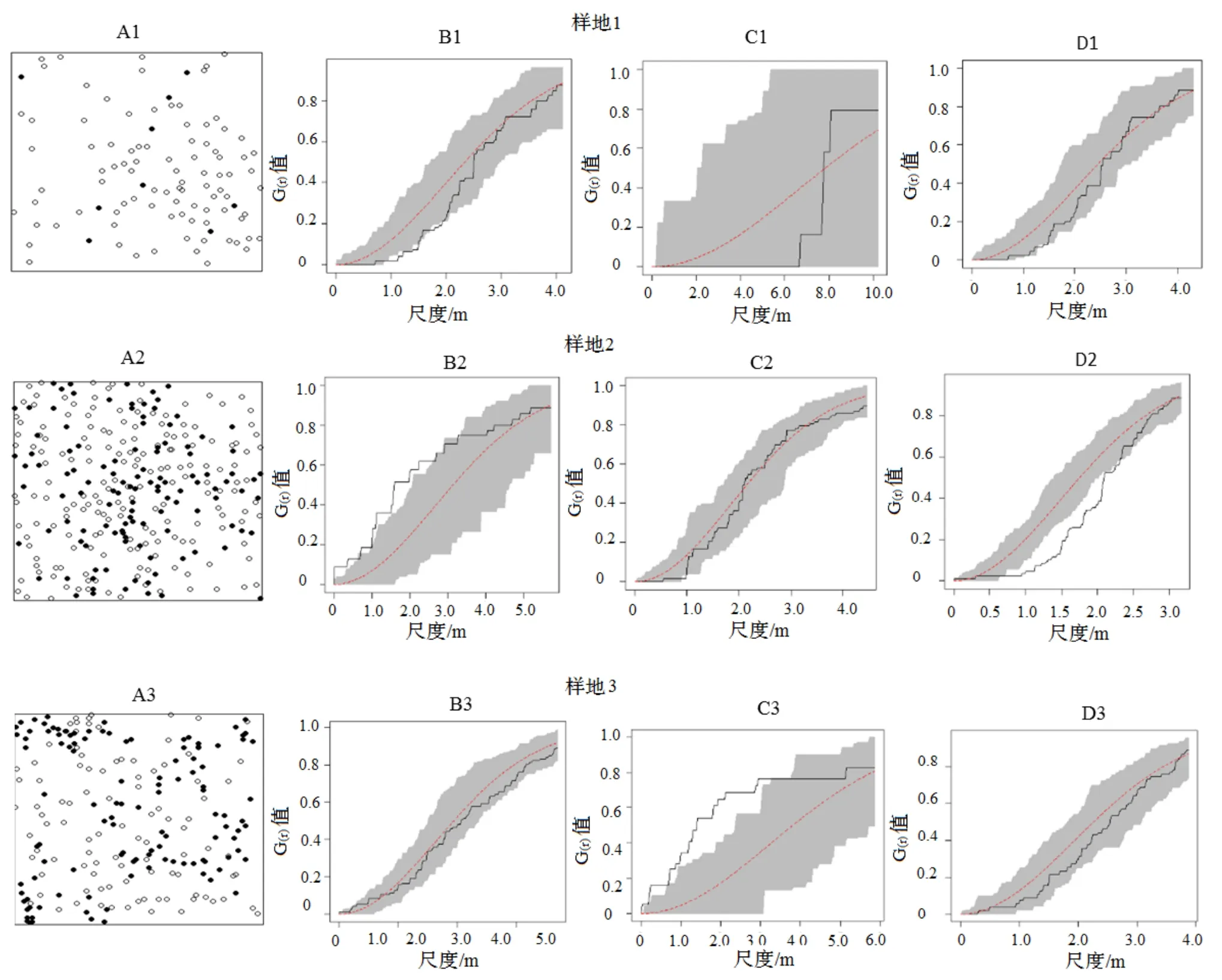
图3 黄山松种群分布格局Figure 3 Spatial patterns of Pinus taiwanensis
本研究发现,黄山松和木荷的不同生活史阶段在空间上具有隔离现象,其空间分布格局在样地间和不同生活史阶段存在差异。植物的空间分布格局与其生长习性、更新、扩散特征和生境异质性等有关[2-3]。黄山松和木荷的种子散布方式均为风力传播[19-20],黄山松为阳性树种,主要依赖林窗进行更新[16];木荷在幼苗阶段具有耐阴性,可在林下或林窗中自然更新[17]。因此黄山松和木荷种子在风力传播下,在林窗内萌发,从而形成母树与幼苗分布的空间隔离现象。
植物不同生活史阶段的生态需求存在差别[4]。本研究将黄山松和木荷分成划分成两个生活史阶段,发现划分生活史阶段和不划分生活史阶段情况下,种群分布格局差异较大,因此研究种群的空间分布格局应考虑个体的发育状态。黄山松一般表现出聚集分布的特点,而在成熟林中主要为随机型或聚集型分布[10-11]。木荷在黄山松幼林中为随机分布,在木荷+黄山松混交阶段为聚集型分布[12]。陈德良[21]在百山祖的研究表明,木荷整个种群和不同发育阶段的分布格局都呈聚集分布。戴云山黄山松聚集格局主要发生在0~26 m 尺度上,>26 m 尺度主要为随机或均匀分布[22]。对浙江古田山木荷种群0~100 m 尺度上的研究发现,随着龄级的增加,植物的自疏或它疏作用增加,木荷幼树到大树种群聚集程度下降[5]。成年阶段的黄山松种群由于种内和种间竞争加剧,其分布格局也会由聚集分布向随机分布转变[18]。因此扩散限制和密度制约效应导致了小尺度上种群聚集分布,而在较大尺度上聚集强度下降。
本研究还发现,黄山松和木荷的空间分布格局具有尺度依赖性特征,即随尺度的增加,聚集类型由聚集分布向随机分布向转变。由于重力的原因,风力传播的扩散方式使得种子以母树为中心聚集分布,但也使得母树周围同种个体的密度制约效应增加,引起个体死亡率增加[23-24]。本研究中木荷种群的大树和小树的分布格局均为小尺度的聚集分布和大尺度的随机分布格局,但是大树的聚集强度减弱;而黄山松种群的大树和小树在群落结构复杂的样地1 中均为随机分布格局,样地2 的大树呈均匀分布,而样地3 的幼树呈聚集分布。因此,树木大径级个体聚集强度的减弱除了与种内竞争导致自梳效应有关外,还与群落结构的复杂性引起的种间关系的变化有关。
本研究对景宁荒田湖林场针阔混交林黄山松和木荷种群分布格局的进行了研究,发现不同生活史阶段个体在空间上存在隔离现象,其分布格局也存在差异。因此在制定经营管理措施时应结合经营目标,合理规划经营面积,同时考虑不同地段的林分结构差异,针对优势树种的不同生长阶段的空间分布特征,合理控制密度,优化林分结构,促林分健康发展。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业科学(2022年5期)2022-08-20
中国林副特产(2022年4期)2022-08-08
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年3期)2022-06-13
散文诗世界(2022年5期)2022-05-23
现代园艺(2021年23期)2021-12-01
园艺与种苗(2021年3期)2021-05-26
花卉(2021年2期)2021-01-21
新农业(2020年18期)2021-01-07
