
基于叶片性状变化下云杉幼苗更新的C,N,P调控机制
2019-03-15 08:59仇宏格杨秀清马慧晶韩有志
山西农业科学 2019年3期
仇宏格,杨秀清,梁 楠,马慧晶,冯 帆,史 婵,韩有志
(山西农业大学林学院,山西太谷030801)
森林的天然更新是森林生态系统进行自我繁衍与恢复的手段,也是森林群落动态的重要组成部分,对未来森林群落的结构以及生物多样性具有深远的影响,对森林生态系统的稳定及进化都具有重要作用[1]。森林更新主要通过幼苗的更新来实现。幼苗更新受自身和环境条件等多个因素调控作用的影响,且以叶片的更新响应最为敏感[2]。C,N和P作为森林树木生长发育所必需的最主要的营养元素[3],参与地球的生物化学循环[4],通过影响森林生态系统的养分周转而影响树木的养分积累与发育状况。森林更新过程中,更新幼苗自身对C,N,P吸收和利用能力以及环境中C,N,P营养水平的变化均会对幼苗的生长以及生理机能产生影响。而且这些影响往往更显著地体现在幼苗叶片性状的变化上,从而反映C,N,P等营养元素对幼苗更新的调控作用[5]。因此,研究基于叶片性状变化下C,N,P对森林更新幼苗的影响,有助于从元素水平上探究森林更新背后潜在的生态学机制。
云杉是华北山地典型的天然林树种资源,属我国特有的寒温性常绿针叶树种[6],也是华北地区重要的用材林树种,对该地区的水土保持和生态环境建设也同时起到了重要作用。本研究以位于华北山地关帝山林区的云杉次生林为研究对象,分析了林下云杉幼苗随更新年龄变化下的叶片形态特征,叶片 C,N,P含量特征,叶片 C,N,P含量与根系 C,N,P含量的相关性及其对叶片形态特征的影响,旨在深入了解叶片性状变化下云杉幼苗更新的C,N,P调控作用,为进一步揭示云杉更新的生态学机制提供重要补充。
1 研究区概况
研究地设在山西省庞泉沟国家自然保护区内,地理坐标为 37°47′45″~37°55′50″N,111°22′33″~111°32′22″E,该区气候属温带季风性大陆山地气候,季节变化明显,年平均温度为4.3℃,1月平均气温-10.2℃,7月平均气温17.5℃,年均降水量822.6mm,年均蒸发量1268mm。土壤类型为棕壤土。
2 研究方法
2.1 样地设置与调查

表1 样地基本情况
在研究地云杉林分布区(海拔为2 010 m)选择立地条件基本一致的地段,设置3块面积为20 m×20 m样地,样地基本情况列于表1。在对样地进行林分调查的基础上,进一步将样地划分为16个5 m×5 m的调查单元,仔细查数更新苗,测量并记录各更新苗的年龄、高度,并挂牌标记。
2.2 样品采集与处理
2017年8月初,在每个样地内分别选择长势较好的各龄级(3,5,7,10,13 a)云杉更新幼苗 5 株作为样株,采集各样株冠层东南方向中上部枝条上健康、完整的功能叶片,放入密封袋,带回实验室。取部分新鲜叶片用于叶片形态指标的测定,其余叶片于85℃烘箱内烘干48 h至恒质量后进行粉碎用于叶片C,N,P含量的测定。同时,在选定的样株干基部50 cm范围内用平板利铲挖取20 cm×20 cm×10 cm土块,采集1~3级的细根作为样品(除去碎土等杂物),装封口袋,标记并带回实验室。将根系清洗、晾干后于85℃烘箱内烘干、粉碎、过筛,再烘干至恒质量备用[7]。
2.3 测定项目及方法
使用叶面积仪(上海,MRS-9600TFU2L)扫描得到各龄级云杉幼苗叶长、叶宽、叶面积、叶周长,并计算叶片的长宽比及比叶面积等叶片形态指标。叶片及根系的烘干样经硫酸-过氧化氢消煮后,采用重铬酸钾-外加热法测定其有机C含量,全氮含量用凯氏定氮法测定,全磷含量用钼锑抗比色法测定[8]。
2.4 数据分析
数据分析采用SPSS 20.0统计软件进行。叶片各形态指标,叶片和根系C,N,P含量采用单因素方差分析(one-way ANOVA),并通过LSD-t检验法检验数据间显著性。叶片形态指标与叶片C,N,P含量,叶片与根系C,N,P含量进行Pearson相关性分析。
3 结果与分析
3.1 不同龄级云杉更新幼苗叶片的形态特征
方差分析显示(表2),云杉幼苗叶片各形态特征指标随更新过程中龄级的增加而变化。其中,叶长、叶宽、叶周长、叶面积均随着龄级的增加而增加,叶长宽比和比叶面积随着龄级的增加而减小。显著性分析表明,叶长和叶周长表现出13 a显著大于3 a,叶宽表现出7 a以上各龄级均显著大于3 a,13 a显著大于7 a以下各龄级。叶长宽比则显示为13 a显著小于3 a。叶面积显示10 a以上各龄级显著大于5 a以下各龄级。比叶面积表现为10,13 a分别显著小于3 a。叶面积和比叶面积的变化表明,叶面积随年龄增加的幅度相对于干质量的增加较小。叶长宽比的变化表明,叶片长度随年龄增加幅度相对于叶宽较小。云杉更新苗叶片形态特征随龄级的变化可能与幼苗C,N,P营养元素对更新的调控作用有关。
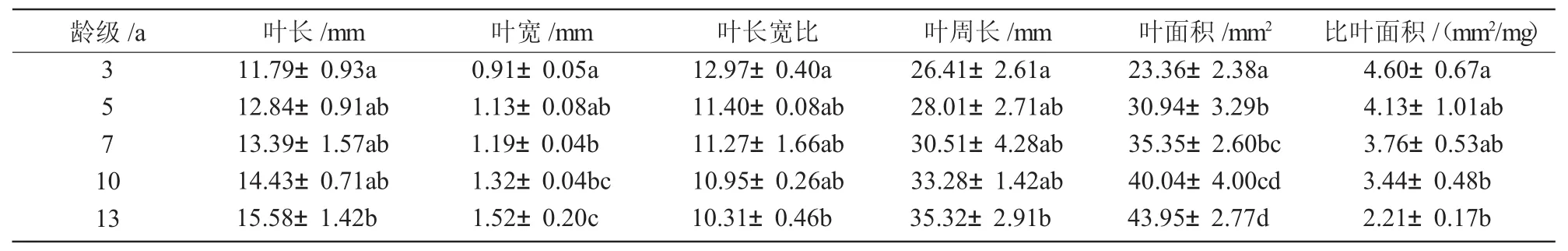
表2 不同龄级云杉更新幼苗叶片的形态特征
3.2 不同龄级云杉更新幼苗叶片与根系的C,N,P含量特征
分析不同龄级云杉更新幼苗叶片的C,N,P含量特征,结果表明,随着龄级的增加,叶片有机碳与全磷含量在增加,全氮含量在减少(图1)。显著性分析显示,10 a幼苗叶片有机碳含量显著高于5 a以下各龄级(5,3 a),13 a幼苗叶片的有机碳含量显著高于其余各龄级。叶片全氮含量表现为3,5,7 a分别显著高于10,13 a。叶片全磷含量在各龄级间未表现出显著差异。
对不同龄级云杉更新幼苗根系C,N,P含量量特征的分析表明,随着龄级的增加,根系C,N,P含量均增加,且有机碳含量在10,13 a与3,5,7 a各龄级间分别呈显著性差异。10 a以上与5 a以下各龄级间全氮含量差异显著,7 a仅与13 a间表现出显著差异。根系全磷含量在各龄级间未表现出显著差异(图 1)。

3.3 云杉更新幼苗叶片形态与C,N,P含量的相 关性

表3 云杉幼苗叶片形态与C,N,P含量的相关性
进一步对各龄级云杉幼苗叶片的形态指标与C,N,P含量做相关性分析,结果表明,叶片各形态指标与其C,N,P含量均显著相关(表3)。其中,叶片有机碳和叶全磷含量分别与叶长宽比、比叶面积呈显著负相关,与叶长、叶宽、叶面积以及叶周长呈显著正相关;叶全氮含量则相反。同时分析也得出,叶长与叶宽、叶面积、叶周长间分别呈极显著正相关。叶宽与叶面积、叶周长呈极显著正相关,但与叶长宽比和比叶面积呈极显著负相关。叶面积与叶周长呈极显著正相关,但与比叶面积呈极显著负相关。叶周长与比叶面积呈极显著负相关。这些结果表明,叶片各形态指标间相互关联,且叶片的C,N,P含量对这些形态特征存在显著影响。
3.4 云杉更新幼苗叶片与根系C,N,P含量的相关性
对各龄级云杉幼苗叶片C,N,P含量与根系C,N,P含量的相关性分析表明(表4),叶片与根系2个器官间除全磷含量无显著相关性外,有机碳含量呈显著正相关,全氮含量呈显著负相关。同时,根系中的有机碳和全氮含量呈极显著正相关。叶片中有机碳与全氮含量呈极显著负相关,但与叶全磷呈极显著正相关。叶片中全氮与叶全磷含量呈极显著负相关。这些分析结果说明,根系中C,N含量的大小及其相互作用很大程度上对叶片C,N含量及C,N,P间的相互作用存在显著影响。

表4 云杉幼苗叶片与根系C,N,P含量的相关性
4 讨论
4.1 云杉更新过程中幼苗叶片的形态特征
在同一立地条件下,树龄是影响幼苗生长发育的内在因素。随着树龄的增加,幼苗逐渐向不同的生长发育阶段过渡,其内部组成结构随之发生变化,幼苗各器官在性状表现上会有所差异。本研究发现,云杉更新苗的叶长、叶宽、叶周长、叶面积均随着龄级的增加而增加,而叶长宽比和比叶面积随着龄级的增加而减小。不同年龄幼苗的这些性状上的变化和差异与幼苗对营养元素吸收和利用能力的差异所导致的幼苗叶片营养元素累积水平密切相关[9]。而且各营养元素间的相互关联性也决定了云杉幼苗叶片各形态指标间存在显著的相关性,在幼苗更新过程中,这种叶片形态间的关联变化对云杉幼苗的生长发育具有重要的生态意义[10]。
4.2 云杉幼苗更新过程中的C,N,P调控机制
C,N,P作为植物体最主要的营养元素,是构成植物干物质、氨基酸、蛋白质、核酸等的生物合成及酶的重要组成部分,调控着植物体生长发育过程中的各种代谢机能且彼此存在着密切的关系[11]。森林更新过程中,幼苗叶片作为更新发生发展的重要同化器官,一方面通过光合作用积累养分,另一方面还要吸收通过根系输送的来自土壤中的各种养分,并依靠不同养分元素间的相互作用和周转来促进幼苗的生长和发育。随着更新幼苗年龄的增加,叶片光合积累养分和通过根系吸收养分的能力也在增加,幼苗叶片物质投资和分配格局也发生变化,导致叶片形态特征也有所差异。
本试验通过探测叶片C,N,P元素水平对更新过程中幼苗叶片形态特征的影响,并进一步主要探测了更新过程中叶片C,N,P变异及其与根系C,N,P变异的相关性,以期在对幼苗叶片性状的年龄变异做出解释的基础上探讨云杉幼苗更新过程中的C,N,P调控机制。试验表明,云杉幼苗叶片有机碳与全磷含量随年龄的增大而增加,全氮则在减少。在云杉更新过程中,随着幼苗年龄的增长和生长速度的加快,幼苗体内需要大量的核糖体来合成蛋白质,而核糖体是植物体中的重要P库[12],因此,随着龄级的增加,叶片的P含量在逐渐增加,这符合生长速率假说[13]。随着生长速度的加快,幼苗的光合作用也在增强,消耗了大量的酶,而酶的本质又是蛋白质,因此,随着龄级的增加,叶片的N含量在减少。光合作用会固定碳,随着龄级的增加,光合作用的增强,叶片积累了大量的碳,因此,随着龄级增加,叶片碳含量在增加。
叶片C,N,P含量与叶片形态特征的相关性表明,有机碳和全磷可促进叶长、叶宽、叶面积及叶周长的增加,同时抑制叶长宽比和比叶面积的增加。这是因为P是蛋白质合成的重要元素,而生长素结合蛋白就控制着细胞的伸长与伸宽[14-15]。而全氮对幼苗更新过程中叶片形态特征的影响与有机碳、全磷相反,幼苗叶片氮含量与比叶面积呈显著正相关。这可能是由于云杉在幼苗阶段针叶不具备较大的光捕获面积,生长速率相对较低,叶片中大部分物质用于构建保卫结构或增加叶肉细胞密度[16],形成比叶面积较小的叶片来延长寿命,同时使叶片内部的水分向表面扩散的距离或阻力增大,从而减少水分的散失,提高植物的水分利用效率[17-18]。同时有研究表明,植物叶片含N越多,其比叶面积越大[19],因此,云杉幼苗叶片氮含量与叶面积显著正相关。
5 结论
云杉幼苗叶片形态指标随更新过程中龄级的增加而变化。其中,叶长、叶宽、叶周长、叶面积均随着龄级的增加而增加,叶长宽比和比叶面积随着龄级的增加而减小。
云杉幼苗叶片有机碳与全磷含量随年龄的增大而增加,全氮则在减少;根系中C,N,P随更新年龄增加均呈增加趋势。叶片和根系C,N,P含量随更新年龄的增加而变化且各含量间相互关联。
云杉幼苗更新过程中,叶片有机碳和全磷促进叶长、叶宽、叶面积及叶周长的增加,同时抑制叶长宽比和比叶面积的增加;全氮对幼苗更新过程中叶片形态特征的影响与有机碳、全磷相反。
根系中C,N含量及C,N,P间的关联变化对叶片C,N含量及C,N,P间的相互作用存在显著影响。叶片C,N,P含量受根系C,N,P含量的影响,并通过影响叶片形态特征从而对云杉幼苗的更新过程起调控作用。
猜你喜欢
农业科技与信息(2022年12期)2022-08-09
生态学报(2022年13期)2022-07-30
西北农林科技大学学报(自然科学版)(2021年7期)2021-08-09
现代农村科技(2020年7期)2020-12-19
河南农业大学学报(2020年2期)2020-05-22
新疆农业科学(2020年1期)2020-02-14
花卉(2020年6期)2020-01-07
河南农业科学(2019年12期)2019-12-05
江苏农业学报(2018年5期)2018-09-10
启迪与智慧·教育版(2018年12期)2018-02-26
