
急性热应激对西伯利亚鲟肝功指标及肝脏热休克蛋白表达的影响
2019-03-11 05:23:56王晓雯朱建亚刘丽丽马国庆
四川农业大学学报 2019年1期
王晓雯,张 蓉,朱建亚,刘丽丽,马国庆,朱 华
(北京市水产科学研究所/渔业生物技术北京市重点实验室,北京100068)
鲟鱼属于硬骨鱼纲、辐鳍亚纲、鲟形目,是我国重要的淡水经济鱼类,在20多个省市均有养殖[1]。西伯利亚鲟(Acipenser baeri)是最重要的鲟鱼养殖品种之一,为亚冷水性鱼类,适宜的生长温度为16.7~24.7℃[2]。鱼类作为变温动物,水体温度是其生长发育最重要的环境因子之一,不但可直接影响鱼类的摄食、生长、免疫和生殖等活动,而且水温改变可间接对鱼类产生影响。超出鱼类生理适应范围的温度都会引起鱼类的应激反应,导致其生理失衡和免疫力下降[3-5]。近年来,全球气候变暖对水产养殖带来的影响逐渐引起人们的关注。随着鲟鱼养殖规模不断扩大,养殖水体负载越来越大,夏季长时间的高温超过鱼类的适应范围,引起鱼类的应激反应、甚至病害频发,给鲟鱼养殖业带来巨大损失[6-7]。
鱼类通过内分泌和生理功能的改变来应对环境的变化,有学者研究表明高温对鱼类的生长[7-9]、抗氧化酶活[8-11]、消化酶活[11-12]、代谢和非特异性免疫指标[13-14]等方面均有不同程度影响前期,王静波等[6]和田照辉等[15]学者们对鲟鱼响应热应激的研究集中于血清抗氧化指标、非特异性免疫指标和鳃组织热休克蛋白70基因(hsp70)的表达变化。然而肝脏作为鱼类消化系统的一部分,是鱼类最主要的消化腺,在鱼类的物质代谢、解毒和防御生命活动中扮演着极为重要的角色,高温应激下鲟鱼的肝脏是否能应对胁迫免受损伤值得研究。血清谷丙转氨酶(ALT)、谷草转氨酶(AST)是动物体内的两种重要氨基转移酶,在平衡机体氨基酸以及蛋白质、脂质与糖类间物质转化中发挥重要作用[16]。健康状况下,ALT和AST主要存在于肝脏中,只有少量被释放到血液中,因此是检测肝脏功能是否正常的重要指标[12]。血液中蛋白含量高低是衡量鱼体的生理状态的指标之一,相对稳定的蛋白浓度表明鱼体生理状态平稳[17],血清蛋白质主要也是在肝脏合成并释放到血液中,故血清蛋白含量也可反应机体肝脏的健康程度。热休克蛋白(heat shock protein,HSP)是一类在生物进化中最保守的应激蛋白,很多学者发现其在环境胁迫因子(高温、重金属和微生物感染等)刺激下表达量显著升高[18-21],D.Parsell、D.A.Picard等[22-23]的研究表明HSP70和HSP90均可通过降解异常蛋白促进蛋白的正确重折叠而在应对热胁迫时发挥重要调节作用。
本研究将从西伯利亚鲟血清的肝功指标和肝脏hsp70和hsp90β的表达变化这两方面重点探究西伯利亚鲟的肝脏对热应激的响应。本研究结果拟为鲟鱼抗高温应激调控提供基础数据,将有助于鲟鱼生产者理解热应激的生理变化、评估热应激的不利影响并做好热应激预防。
1 材料和方法
1.1 试验鱼及饲养
西伯利亚鲟幼鱼来自北京市十渡鲟鱼繁殖基地,体重为(34.905±3.761)g,挑选规格统一的健康西伯利亚鲟进行试验。试验前在循环水系统中暂养7 d,水温(23±1)℃。养殖容器为圆柱形PP水槽,内径约1 m,水深0.5 m;试验用水为曝气后的自来水。试验期控制水温(23±1)℃,日投饵量为鱼体质量的1%,每天分3次投喂;定期排污。取样前24 h内停止投喂。
1.2 半致死高温的确定
将120尾23℃暂养的试验鱼随机转入30℃、31℃和32℃和33℃的水体(60 cm×40 cm×40 cm),每个温度组设置3个平行,每个平行10尾鱼。充入足够的空气,观察96 h内鲟鱼的死亡情况,及时捞出死鱼,计算累计死亡率。
1.3 高温胁迫试验
根据半致死高温试验结果,选择30℃作为急性热应激的水温。23℃暂养7 d后,采集6尾鱼作为0 h样品,其余60尾西伯利亚鲟迅速转入30℃的水体,并设置3个水槽,作为平行组,进行热应激试验。分别在热应激后3、6、12、24、48和96 h,每组随机选取6尾鲟鱼,MS-222麻醉后尾静脉取血,采集肝脏样品,液氮速冻后转至-80℃保存,用于提取RNA。采集的血液室温放置2 h,4℃静置16 h,3 000 r/min离心10 min,取上清,制得血清,用于测定血清的肝功指标。
1.4 血清谷丙转氨酶、谷草转氨酶活力和总蛋白浓度的测定
谷丙转氨酶活力(ALT)、谷草转氨酶活力(AST)测定采用微量酶标法。
谷丙转氨酶活力以卡门氏单位表示,定义为1 mL液体,反应液总容量3 mL,波长340 nm,1 cm光径,25℃,1 min内与底物L-丙氨酸和α-酮戊二酸所生成的丙酮酸,使还原态烟酰胺腺嘌呤二核苷酸氧化成氧化态烟酰胺腺嘌呤二核苷酸而引起吸光度每下降0.001为一个单位(1卡门氏单位=0.482 IU/L,25℃)。
谷草转氨酶活力以卡门氏单位表示,定义为1 mL液体,反应液总容量3 mL,波长340 nm,1 cm光径,25℃,1 min内与底物L-天门冬氨酸和α-酮戊二酸所生成的草酰乙酸,使还原态烟酰胺腺嘌呤二核苷酸氧化成氧化态烟酰胺腺嘌呤二核苷酸而引起吸光度每下降0.001为一个单位(1卡门氏单位=0.482 IU/L,25℃)
血清总蛋白浓度的测定采用考马斯亮蓝法,使用考马斯亮蓝测定试剂盒测定。
白蛋白浓度采用溴甲酚绿比色法。有非离子型表面活性剂存在时,溴甲酚绿可与白蛋白形成蓝绿色复合物,颜色的深度与样本中白蛋白浓度成正比。以上测定均使用测试试剂盒(南京建成生物工程研究所),在Econ全波长酶标仪(BioTek)上进行测定。
1.5 西伯利亚鲟肝脏组织RNA的提取和cDNA的合成
取大约3 mg肝脏组织加入800μL TRIzol(Takara),按照该试剂附的操作方法采用氯仿抽提法提取总RNA,1%琼脂糖凝胶电泳检测。
以提取的总RNA(浓度为300 ng/μL)为模板,采用TransScript First-Strand cDNA Synthesis Super Mix试剂盒(Takara),以Oligo(dT)为反转录引物,按试剂盒操作手册合成cDNA。根据NCBI上已有的hsp70(HM348777.1)和hsp90β(JX477807.1)cDNA序列,使用Primer 5.0软件设计实时荧光定量引物,分别为hsp70-F,hsp70-R,hsp90β-F,hsp90β-R,引物序列见表1,并由北京擎科新业生物技术有限公司合成。
标准曲线的制作:对部分样品cDNA进行混合,进行4倍梯度稀释,以稀释后的cDNA为模板进行实时荧光定量PCR,每个模板设3个重复。荧光定量PCR反应体系(20μL):SYBR Premix ExTaq10μL,上下游引物各0.4μL,模板1μL,ddH2O 8μL,ROX II 0.2μL。反应条件为:95℃预变性5 min,95℃15 s,59.6℃30 s,40个循环。实时连续测定扩增过程中的荧光值,建立标准曲线。
以西伯利亚鲟18S rDNA(引物设计为18S-F,18S-R)作为内参,使用ABI7500荧光定量PCR仪,对热应激后所有肝脏cDNA样品进行分析,反应体系及反应条件与标准曲线的建立一致,每个样品设3个重复。设定0 h样品该基因的表达量为“1”,结果采用2-ΔΔCt法计算。

表1 试验中用到的引物序列Table 1 Primers used in the experiment
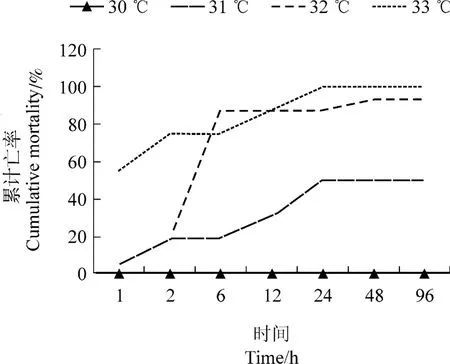
图1 不同温度应激下西伯利亚鲟幼鱼的累计死亡率Figure 1 The accumulative total mortality of Acipenser baeriunder different water temperature
1.6 数据处理
试验数据均用平均值±标准差表示。利用SPSS17.0软件的ANOVA单因素方差分析,对不同组织间均值进行统计学显著性差异分析。首先对数据进行正态分布和方差齐性检验,如果ANOVA分析有意,再用Duncan进行两两比较。显著性水平设为P<0.05。
2 结果与分析
2.1 西伯利亚鲟幼鱼的半致死高温
96 h内不同高温应激下西伯利亚鲟的死亡情况,见图1。试验结果表明,西伯利亚鲟幼鱼(34.905±3.761)g的半致死高温为31℃。30℃应激下,鲟鱼没有发生死亡,考虑到急性热应激试验选择一个温度高但不发生死亡的温度,所以选择30℃作为本试验的急性热应激温度。
2.2 热应激对西伯利亚鲟的血清谷丙转氨酶活力、谷草转氨酶活力的影响
在热应激后(见图2),西伯利亚鲟血清谷丙转氨酶活力逐步升高,48 h时达到峰值,显著高于0时刻(P<0.05)。然而在96 h时,其谷丙转氨酶活力迅速降低至0时刻水平。
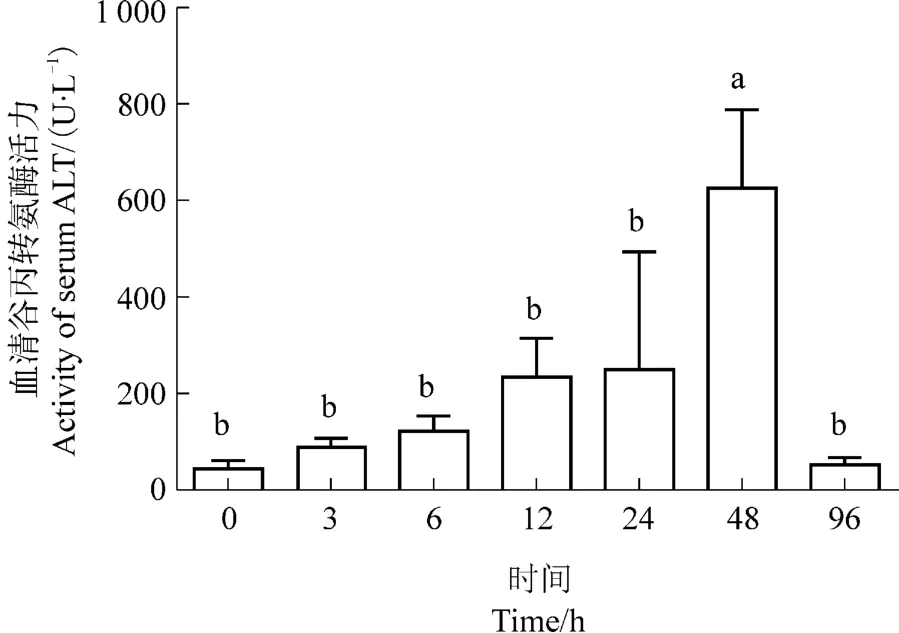
图2 热应激对西伯利亚鲟血清谷丙转氨酶活力的影响Figure 2 Effects of heat stress on the activity of serum ALT in Acipenser baeri
在热应激6 h,西伯利亚鲟血清谷草转氨酶活力开始缓慢升高,48 h较24h迅速升高,并达到峰值(图3)。96 h时,西伯利亚鲟谷草转氨酶活力有所下降,但仍显著高于0时刻的酶活力(P<0.05)。
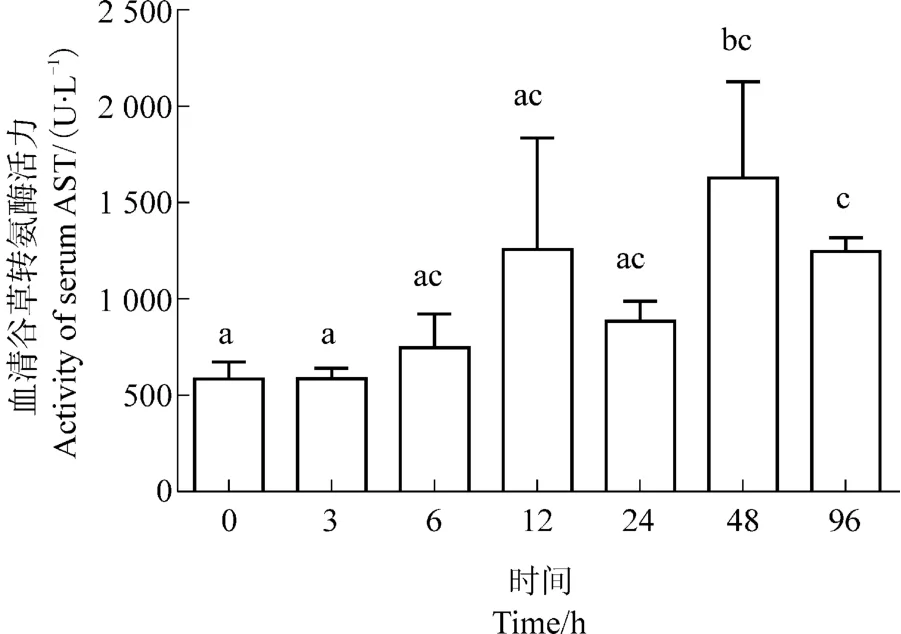
图3 热应激对西伯利亚鲟血清谷草转氨酶活力的影响Figure 3 Effects of heat stress on the activity of serum AST in Acipenser baeri
2.3 热应激对西伯利亚鲟血清蛋白浓度的影响
如图4所示,热应激后3~12 h,西伯利亚鲟幼鱼的血清总蛋白浓度缓慢上升,于12 h达到一个较高点,至24和48 h血清总蛋白浓度有所下降,96 h达到0时刻水平。可见,血清总蛋白浓度整体维持恒定,鱼体生理状态平稳。
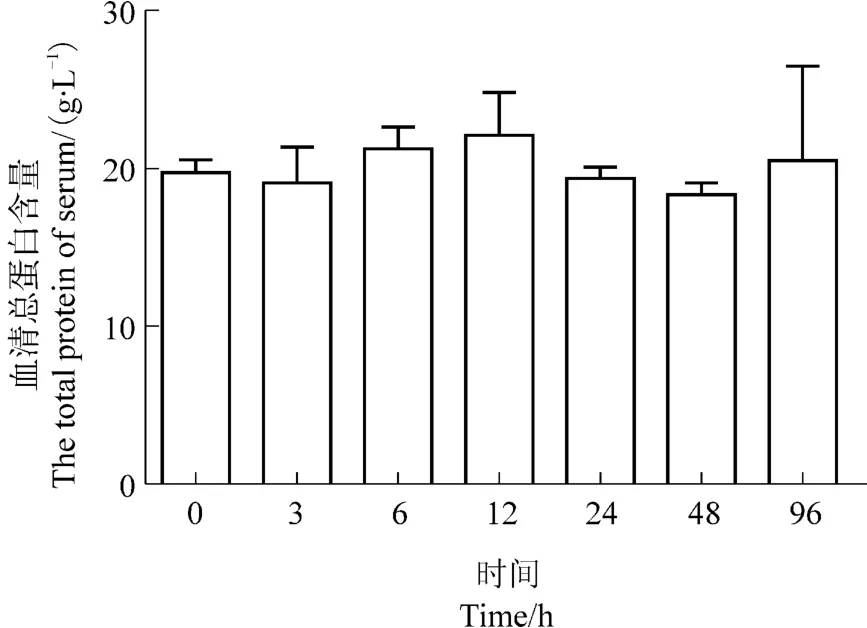
图4 热应激对西伯利亚鲟总蛋白浓度的影响Figure 4 Effects of heat stress on the serum total protein concentrations in Acipenser baeri
西伯利亚鲟幼鱼血清白蛋白浓度除24 h外,在热应激后不同时间点,均显著低于0时刻,但各时间点之间无显著差异(见图5)。可见,高温刺激后,西伯利亚鲟幼鱼肝脏的功能受到影响,白蛋白合成量有所减少。
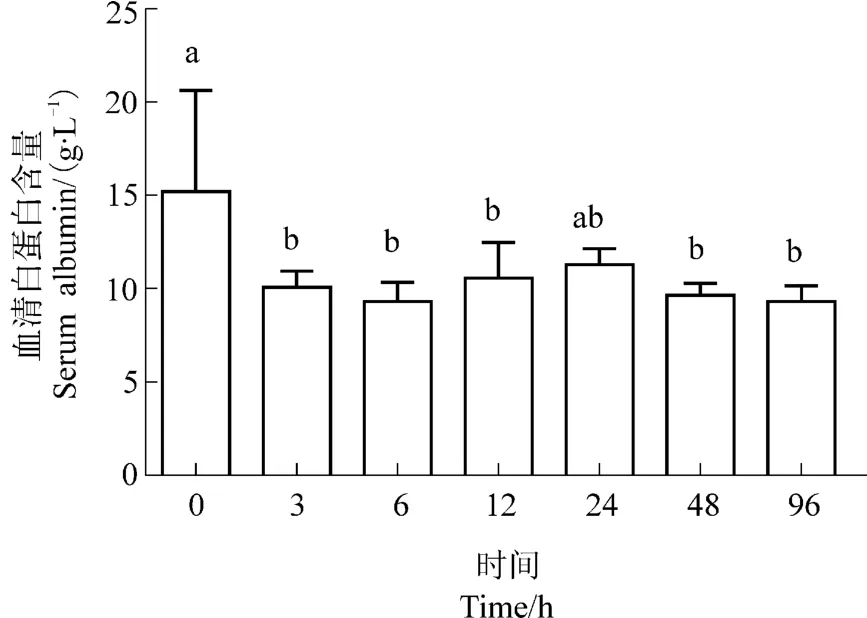
图5 热应激对西伯利亚鲟白蛋白浓度的影响Figure 5 Effects of heat stress on the serum albumin concentrations in Acipenser baeri
2.1 热应激下西伯利亚鲟肝脏hsp70和hsp90β的表达情况
应用Applied Biosystems7500荧光定量PCR仪自带软件程序设置生成标准曲线。以CT值作为纵坐标,基因相对拷贝数的对数作为为横坐标制作标准曲线,结果如表2所示。3个标准曲线的R2值均大于0.99,具有良好的线性关系,扩增效率E值均位于96.25%~110%之间,说明这3对荧光定量引物可用于后续试验。
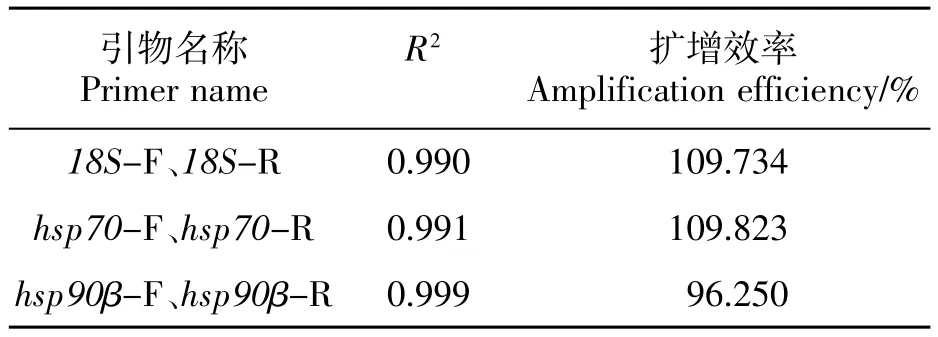
表2 3对引物的扩增效率及拟合度Table 2 Amplification efficiency and degree of fitting of three pairs of primers
热应激下西伯利亚鲟肝脏hsp70和hsp90βmRNA的相对表达量均呈现先升高后降低的趋势(图6,7),二者表达量在6 h达到峰值(P<0.05),hsp70mRNA的转录水平达到0时刻的15.61倍,hsp90βmRNA的转录水平达到0时刻的15.55倍。西伯利亚鲟肝脏hsp90β对热应激的反应更敏感,3 h时其相对表达量即升高到0时刻的7.43倍,高于hsp70mRNA的相对表达量(4.66倍)。48 h,hsp70mRNA和hsp90βmRNA的转录水平均迅速降低,于热应激96 h时恢复到0时刻水平。
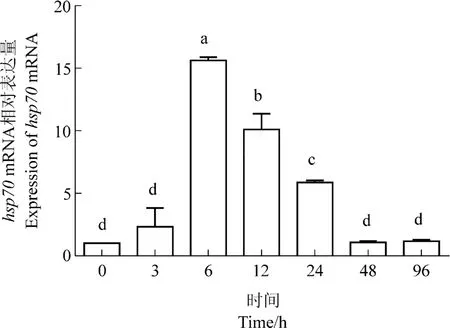
图6 热应激对西伯利亚鲟肝脏hsp70 mRNA相对表达量的影响Figure 6 Effects of heat stress on the expression levels of hsp70 mRNA in Acipenser baeri liver
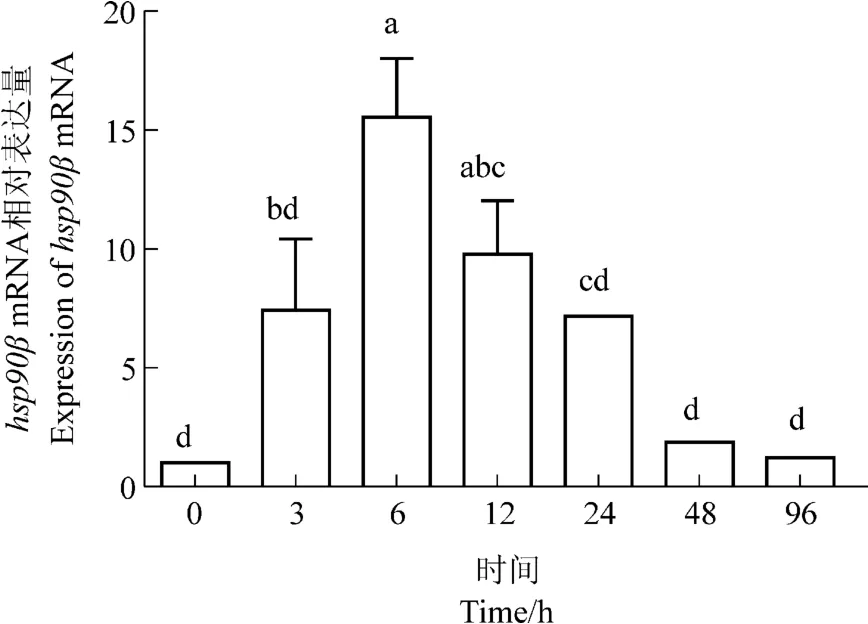
图7 热应激对西伯利亚鲟肝脏hsp90βmRNA相对表达量的影响Figure 7 Effects of heat stress on the expression levels of hsp90βmRNA in Acipenser baeri liver
3 讨论与结论
水温是水产养殖中最重要的环境因子之一,与鱼类的代谢相关,影响鱼类生理生化,当水温超过鱼体耐受极限将导致鱼体代谢紊乱,甚至发生死亡[24]。研究西伯利亚鲟幼鱼的致死温度有利于了解其耐高温能力,为鲟鱼的养殖提供参考数据。本研究采用急性应激的方法,得到西伯利亚鲟幼鱼从23℃的养殖水温中直接升到31℃可引起死亡。李文龙等[24]在3种鲟科鱼类临界水体温试验中(7.6±0.3)g的小体鲟、史氏鲟和俄罗斯鲟的存活最高温度分别为32℃、33℃和33℃,较本研究中西伯利亚鲟的致死温度略高,这是由于李文龙等的研究是采取梯度升温的方式,鱼体已逐渐产生耐受性,所以耐受的温度比急性升温的耐受温度要高。
鱼类的血清酶主要来自特定的组织器官,其活性的高低与相应组织器官的代谢水平和健康程度有关,血清酶活的变化反映了组织器官功能的变化。ALT和AST是机体最重要的反映物质转化的代谢酶,由肝脏分泌表达,在鱼体健康状况下,释放到血液中的含量较少。因此ALT和AST的酶活变化是反应机体肝脏功能的重要指标。管标等[12]在虹鳟(Oncorhynchus mykiss)受到热应激后,血清ALT和AST活性均在恢复时有显著升高,肝脏受到破坏作用。李佳凯等[25]的研究显示大黄鱼(Larimichthys crocea)幼鱼从25℃到32℃维持7 d的高温胁迫后血清ALT和AST活力极显著高于对照组。在本研究中,西伯利亚鲟幼鱼在急性热应激下血清ALT和AST酶活力呈现先升高后降低的趋势,均在热应激后48 h达到峰值,显著高于0时刻(P<0.05),表明鲟鱼受到热应激48 h内肝脏功能受到影响,并在96 h逐步恢复。血清蛋白主要在肝脏合成,肝功能的损伤程度影响着蛋白的合成,血清总蛋白及白蛋白含量检测可及时反映肝脏的蛋白合成能力。半滑舌鳎(Cynoglossus semilaevis)在受到高温胁迫后,血清白蛋白持续下降[26]。高温和低温急性胁迫均会使期许氏平鮋(Sebastes schlegelii)血清总蛋白和白蛋白水平著降低[27]。大黄鱼在高温胁迫7 d时,血清总蛋白含量同样显著低于对照组[25]。本研究中,急性热应激后,西伯利亚鲟幼鱼的血清总蛋白含量总体维持稳定,在胁迫24 h和48 h其含量略有降低,而白蛋白含量在胁迫3~96 h除24 h外均显著低于0时刻(P<0.05)。说明高温胁迫后,西伯利亚鲟幼鱼的肝脏损伤不是特别严重,血清蛋白能基本维持稳态,这与他人的研究结果不同,可能高温胁迫对不同鱼类血清蛋白的影响各有不同。而白蛋白作为营养物质的载体,为机体提供能量,参与维持血浆渗透压,急性热应激后,机体大量消耗白蛋白,以满足机体较高的能量需求。
热休克蛋白(HSP)是一类保守的,参与生物体应对多种应激过程的蛋白质,如应对热应激、清除体内氧自由基、诱导细胞凋亡和加强免疫反应[28]。HSP70和HSP90是这个家族中分别为70 kDa和90 kDa大小的蛋白质家族[29]。本研究中,西伯利亚鲟幼鱼肝脏的hsp70和hsp90β均在急性热应激后出现显著性上调表达,表明肝脏存在一定的损伤,热休克蛋白可能通过上调表达而发挥保护细胞的作用。这与Rendell J.L.等[30]研究发现hsp90在O.mykiss的肝脏组织中有显著的上调表达,Wu C.X.等[31]在高温应激下得到草鱼(Ctenopharyngodon idella)hsp90表达量上调最显著的组织是肝脏,以及Wang Y.等[32]在鲤鱼(Cyprinus carpio)肝脏中hsp70的显著上调表达相一致。众多研究表明HSP70可快速并显著地上调表达以适应多种环境应激,被广泛地作为一个生物标志[33]。本研究也证实了西伯利亚鲟肝脏hsp70出现显著上调反应以应对热应激,补充了田照辉等[15]的关于西伯利亚鲟受到急性热应激并维持仅3 h的研究,得到hsp70在热胁迫后表达量达到峰值的时间是6 h。HSP90也是真核细胞中一类可与超过400种蛋白结合的伴侣蛋白,在机体受到多种应激条件下,如热应激或冷应激,均能上调其表达[34]。栉孔扇贝(Chlamys farreri)受到热刺激后,hsp90mRNA表达量显著上调[35],泥鳅(Misgurnus anguillicaudatus)在高温应激下,hsp70、hsp90α和hsp90βmRNA表达量均迅速升高[36]。并且,Wu C.X.等对草鱼的研究显示热应激和冷应激均会显著上调多种组织HSP90基因的表达量[31],这些均与本研究结果相一致。
综上所述,本文对一种软骨硬鳞鱼类-西伯利亚鲟幼鱼肝脏组织在受到急性热应激下的血清肝功指标及肝脏热休克蛋白70和90的转录表达进行分析。结果提示我们:西伯利亚鲟幼鱼的肝脏功能在急性热应激下受到一定影响,但是在应激96 h后均恢复到正常水平;肝脏中hsp70和hsp90β均上调表达,参与生理调节,以应对高温对肝脏细胞的损伤。
猜你喜欢
古今农业(2021年2期)2021-08-14 01:43:14
小读者(2019年20期)2020-01-04 02:13:32
武汉冶金管理干部学院学报(2018年3期)2018-11-15 08:16:06
大自然探索(2017年1期)2017-02-14 00:13:36
动物营养学报(2015年10期)2015-12-01 03:30:54
动物营养学报(2015年10期)2015-12-01 02:26:22
应用海洋学学报(2014年1期)2014-11-22 07:17:50
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:21:00
探索地理(2013年3期)2014-01-09 06:37:28
食品科学(2013年23期)2013-03-11 18:29:51
