
双孢菇渣堆肥基质对夏季草莓生长及光合特性的影响
2019-03-06 04:24,,,,,,,
河南农业科学 2019年2期
,, ,,,,,
(山西省农业科学院 现代农业研究中心,山西 太原 030031)
草莓因其果实色泽鲜艳,风味独特,具有较高的营养价值而深受广大消费者的青睐[1]。近年来,为了提早或延后草莓的上市时间,设施草莓栽培技术在世界各地纷纷涌现。草莓的基质栽培是设施栽培的主要形式,是未来草莓栽培的发展方向,以草炭为主要原料的复合基质栽培是发展的重点[2-3]。然而,草炭属于一种不可再生资源,过量开采易破坏生态环境,世界各国均提出要限制草炭资源开采,保护生态环境,也造成了草炭市场价格的提升。国内外学者在寻找草炭替代品方面进行了大量的研究,对减轻环境压力和保障设施草莓栽培的可持续发展都具有重要意义[4-6]。
近年来,随着人们生活水平的提高,食用菌需求增长迅猛,2015年,我国食用菌产量达到3 471.1万t,年均增速12%[7]。食用菌产业的快速发展,产生大量的菌渣,菌渣是很好的可再生资源,菌渣替代草炭成为当前研究的热点。WEVER等[8]研究认为,菇渣是一种潜力较大的泥炭替代物,但存在电导率和pH值偏高等问题。魏启舜等[9]研究认为,未经处理的菇渣可以部分替代泥炭栽培草莓,但替代量有限。李美英等[10]研究发现,菇渣与珍珠岩配比可以调节菇渣水气比,改善作物根际环境,与蛭石配比可以增加其持水孔隙度,增强保水能力。赵荷娟等[11]研究表明,加入一定量的发酵双孢菇菌糠能够提高基质养分含量,有机质、氮、磷、钾含量都有一定的提高,而按V(菌糠)∶V(草炭)∶V(珍珠岩)∶V(蛭石)=1∶3∶1∶1基质配比下种植的草莓生长优于其他处理。齐慧霞[12]研究表明,以草炭、棉籽皮(食用菌废料)作为栽培基质,草莓植株的营养生长和生殖生长均优于其他栽培基质。张全军[13]研究表明,草炭、菌渣、砂土复合基质有利于提高草莓果实的产量和品质。张辉明等[14]研究了不同配比复合基质对草莓生长的影响,结果表明,基质配比为V(菇渣)∶V(锯木屑)∶V(玉米秆)=3∶4∶4时,能显著促进草莓的生长,提高单株产量,在草莓生产中可以替代专业生产基质。樊金山等[15]以发酵腐熟的杏鲍菇渣为原料,研究得出V(草炭)∶V(菌渣)∶V(珍珠岩)∶V(蛭石)=3∶1∶1∶1是草莓生长的最佳基质组合,能提高其产量和品质。不同基质原料形成的复合基质对草莓生产的影响各不相同,不同地区菇渣的种类不同,组成和性质也存在较大差异,很难形成性质稳定的产品。为此,将双孢菇渣堆肥发酵腐熟后部分替代泥炭,用于夏季草莓的设施基质栽培研究,通过分析不同基质配比对夏季草莓生长和光合特性的影响,以获得适宜基质配方,为夏季草莓基质栽培和双孢菇渣资源化利用提供理论依据和技术支持。
1 材料和方法
1.1 试验概况及试验材料
试验于2017年5月15日至11月18日在山西省朔州市山阴县薛圐圙乡半道地农业园区1号拱棚内进行。拱棚长60 m、宽8 m、脊高3 m。
草莓品种为蒙特瑞(Monterey),草莓种苗由山西好乐农业开发有限公司提供。
双孢菇渣来源于山西宇昊蘑菇种植有限公司,发酵腐熟后理化性状为:容重0.725 g/cm3,总孔隙度54.03%,持水空隙度43.41%,气水比0.24,pH值7.22,电导率(EC值)6.09 mS/cm;草炭购于北京大汉有限公司;珍珠岩购于太原华海保温材料有限公司;其他辅助材料购于当地农资市场。
1.2 试验设计
试验采用单因素完全随机设计,设6个处理,包括5个配方和1个对照,每个处理重复3次。采用全封闭地上槽式栽培模式(槽长10 m、宽30 cm、高40 cm),槽间距60 cm,底部装填煤矸石约15 cm后铺设聚乙烯塑料布,防止土壤盐分进入基质,然后槽中间置凹槽(深5 cm、宽10 cm),沿凹槽铺设DN32的包有无纺布渗水管,用沙子覆盖铺平,再填装栽培基质直至装满整个栽培槽。草莓株距为20 cm,水肥通过自行设计的水肥一体化灌溉系统施加,水肥和草莓栽培日常管理统一进行。具体配方见表1。

表1 不同处理草莓基质配方(体积比)Tab.1 Substrate formula of strawberry in different treatments(by volume)
1.3 测定项目及方法
1.3.1 生长指标测定 分别于6月1日、6月21日、7月11日、7月31日和8月20日的8:00—11:00测定草莓生长指标。用直尺测量草莓的株高,中心展开叶向外第3片叶的叶柄长、叶长与叶宽。用游标卡尺测其叶柄中部直径。计算叶面积,叶面积=叶长×叶宽×0.73[16]。每个重复取5株挂牌进行测量,最后求平均值。
1.3.2 光合指标测定 在初果期,选择晴朗天气进行光合指标测定[光照强度约为1 000 μmol/(m2·s),温度为(30±1)℃]。测量净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。并计算气孔限制值(Ls):Ls=1-Ci/Ca,其中,Ca为测定时空气中的CO2浓度[17]。测定时选取长势良好且一致的植株,植株的叶片由内到外第2~3片叶子,选好叶片后标记固定,重复3次,取其平均值。
1.4 数据处理
采用Excel 2003对原始数据进行处理,用SAS 9.1统计软件对数据进行单因素方差分析和LSD 多重比较。
2 结果与分析
2.1 不同处理对草莓株高的影响
由表2可知,在5个生长时期,T1—T4处理株高均高于CK,T5处理株高均低于CK。T1处理株高在5个时期均极显著或显著高于CK,在7月11日和7月31日均高于其他处理,但与T2和T3差异不显著。T2处理的株高除7月11日和7月31日外,均显著高于CK。T3处理株高除7月11日外也均极显著或显著高于CK。T3处理草莓株高除在7月11日和7月31日2个生长时期低于T1外,其他生长时期株高从高到低依次均为T3>T1>T2>T4>CK>T5。
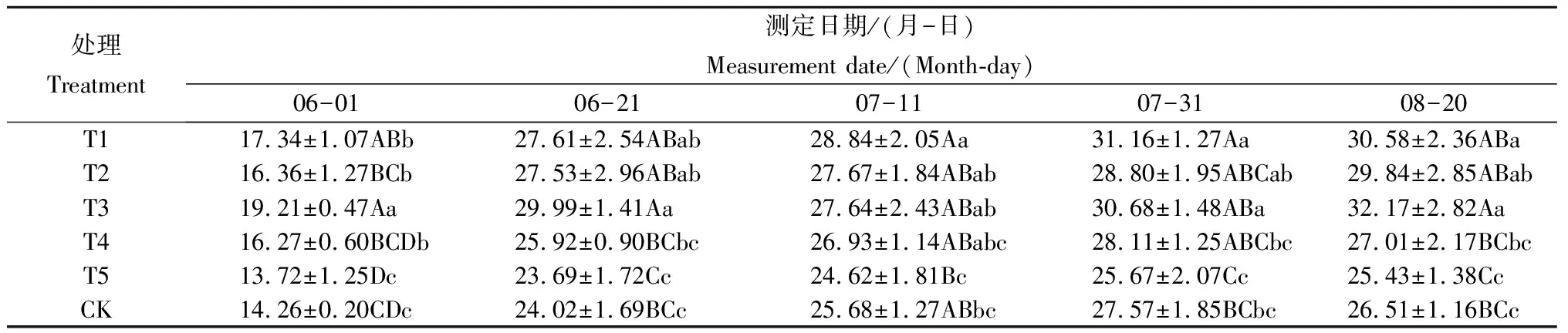
表2 不同处理不同生长时期草莓株高比较
注:同列不同大、小写字母分别表示差异极显著(P<0.01)、显著(P<0.05)。下同。
Note:Different capital letters and little letters in the same column indicate extremely significant difference at the 0.01 level and significant difference at the 0.05 level.The same below.
2.2 不同处理对草莓叶柄长的影响
由表3可知,不同时期T5处理叶柄长均低于CK,其余处理大体上高于CK。T1处理在7月11日和7月31日均极显著高于CK,T3处理在6月21日、8月20日极显著高于CK。T2和T4处理在8月20日均高于CK,但差异不显著。结果表明,T3和T1处理相比其他处理更有利于叶柄的伸长,除7月11日外,T3处理在各个时期叶柄长均优于T1处理。

表3 不同处理不同生长时期草莓叶柄长比较
2.3 不同处理对草莓叶柄粗的影响
从表4可以看出,T3处理在6月21日、8月20日2个生长时期叶柄粗均显著高于CK,其他时期各处理叶柄粗与CK差异均不显著。T3和T4处理的叶柄粗在7月11日低于CK,T4处理的叶柄粗在7月31日也低于CK,T5处理在5个时期叶柄粗均低于CK。除此之外,各处理在不同时期叶柄粗均高于CK,但差异均不显著。由此可见,在整个生长期T3处理相比其他处理叶柄增粗更明显。

表4 不同处理不同生长时期草莓叶柄粗比较 Tab.4 Comparison of strawberry petiole diameter at different growth stages with different treatments mm
2.4 不同处理对草莓叶面积的影响
由表5可知,5个处理在6月1日和6月21日2个生长时期草莓叶面积均高于CK,且T1—T4处理与CK差异呈极显著水平,T5与CK差异不显著,叶面积从高到底依次为T3>T1>T4>T2>T5>CK。到7月11日,5个处理的叶面积则全部低于CK,T1—T4处理与CK差异均不显著,T5则显著低于CK。在7月31日,T5处理草莓的叶面积显著低于CK,其他处理与CK之间无显著差异。在8月20日,5个处理的叶面积均高于CK,T1处理与CK差异显著。除7月11日外,T1和T3处理叶面积在不同时期均高于其他处理,但差异不显著,说明T1和T3处理对叶面积的增加效果略优于其他处理。

表5 不同处理不同生长时期草莓叶面积比较Tab.5 Comparison of strawberry leaf area at different growth stages with different treatments cm2
2.5 不同处理对草莓光合特性的影响
由表6可知,除T2的净光合速率比CK高外,不同处理双孢菇渣堆肥基质栽培草莓的净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均低于CK;气孔限制值均高于CK。T1处理草莓的净光合速率显著低于T3处理,极显著低于其他处理。T1和T4处理草莓的蒸腾速率显著低于T3处理和CK,与其他处理之间无显著差异。T1处理草莓的气孔导度极显著低于CK,显著低于T2和T5处理,而与其他处理之间无显著差异。各处理胞间CO2浓度与CK之间均无显著性差异。T4处理草莓的气孔限制值极显著高于CK,显著高于T1和T2处理。

表6 不同处理草莓光合特性比较Tab.6 Comparison of strawberry photosynthetic characteristics with different treatments
3 结论与讨论
基质是无土栽培成败的关键,它不仅起固定和支持植物的作用,更重要的是为植物生长发育提供环境,基质的结构和营养特性直接影响植物的栽培效果[18]。时连辉等[19]研究表明,经处理的菇渣部分替代草炭与其他基质原料进行混配,才能作为园艺基质使用,与本次试验结果相似。同时本试验还发现,随着双孢菇渣堆肥添加比例的增加,草莓的株高、叶柄长、叶柄粗和叶面积均有降低的趋势,与魏启舜等[9]、赵荷娟等[11]的研究结果相一致,但与马义胜[20]的研究结果不一致,可能与菇渣的种类及组成不同有关。光合作用是植物生长发育、物质积累的基础,叶片是植物进行光合作用的主要器官,是决定植株光合能力的主要因素[21]。植物光合作用受到限制的原因通常分为气孔限制和非气孔限制[22-23]。本研究结果表明,双孢菇渣堆肥基质不同程度降低了草莓的胞间CO2浓度和气孔导度,导致净光合速率下降,蒸腾速率降低,说明各处理对水分运输和调节的能力较差,对草莓的气孔限制增加,不利于植物对空气中CO2的利用,植株光合作用效果较差,进一步说明各处理光合作用的下降主要是由气孔因素引起的,与本试验各处理气孔限制值不同程度升高相一致。FARQUHAR等[24]研究认为,营养液EC值对草莓叶片净光合速率的影响是与叶肉细胞光合活性下降有关的非气孔因素造成的,本研究中双孢菇渣EC值较高,双孢菇渣堆肥基质不同程度降低草莓叶片净光合速率是否与其EC值过高有关有待进一步研究。综上所述,在草莓初果期,双孢菇渣堆肥部分替代草炭形成的复合基质虽不同程度降低其净光合速率和蒸腾速率,但在整个生长期能促进草莓的生长,其中以T3处理[V(双孢菇渣)∶V(草炭)∶V(珍珠岩)=1∶2∶1]效果较好。
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31
小天使·四年级语数英综合(2022年6期)2022-06-21
作文·小学低年级(2021年4期)2021-11-02
作文小学中年级(2021年4期)2021-05-07
农村百事通(2020年20期)2020-11-23
贵州农业科学(2020年10期)2020-11-22
农产品加工(2020年5期)2020-04-24
热带农业科技(2019年3期)2019-07-19
南方农业·下旬(2018年12期)2018-03-28
江苏农业科学(2016年11期)2017-03-21
