
核桃主枝环剥后不同部位光合光响应特性
2019-03-02 13:31余晓娥徐业勇虎海防王宝庆
新疆农业科学 2019年12期
余晓娥,欧 源,徐业勇,虎海防,王宝庆,齐 成
(1.新疆农业大学林学与园艺学院,乌鲁木齐 830052;2、新疆林业科学院,乌鲁木齐 830063;3、新疆林业科学科技推广处,乌鲁木齐 830063;4、新疆林业科学科园林绿化研究所,乌鲁木齐 830063)
0 引 言
【研究意义】光合作用是植物生长发育不可或缺的生理活动,通过光合作用,绿色植物的叶绿素可将太阳光转化为化学能[1]。植物在光合作用时,不仅受到植物自身的影响,还受到周围生态环境因子的影响[2]。在光合作用中,最主要的生态因子为光照,随着光照强度的变化,植物光合能力也发生了变化[3]。在对植物光合作用进行钻研时发现植物光合作用的光响应曲线反应的是植物光合有效辐射与光合速率两者的关系,同时也反映了植物光合特性中的一些相关生理参数,在研究中将通过相关参数对植物光合作用是否正常进行判别,也用于判别植物在不同光照条件下植物对光的适应能力及植物光合作用的能力等,因此,测定植物的光合作用并拟合光响应曲线来获取植物的光合生理特性,为新疆核桃育种提供一定的科学数据[4-12]。【前人研究进展】核桃为胡桃科核桃属落叶乔木,是我国主要的经济林树种之一,其果实含有大量的蛋白质、氨基酸以及糖类等人体必需的矿物元素。近年我国从事核桃光合钻研的专家学者较多,但对环剥以后核桃光合的研究仍然较少。我国核桃品种丰富,尤其在新疆地区,核桃种类较多,研究和分析核桃主枝环剥[13-15]之后的生理变化及其特征,对核桃的栽植、嫁接以及优化果实品质、提高核桃产量等具有重要研究意义[16-20]。【本研究切入点】以环剥核桃主枝上部、下部、未环剥核桃3种叶片为研究对象,测定了不同部位核桃叶片的光合特性,分析了不同部位核桃叶片在不同光强下光合能力的规律[21-23]。低产劣质树树冠内堂空虚,结果枝量明显降低,且结果部位外移的核桃进行嫁接。研究核桃主枝环录后不同部位光合光响应特性。【拟解决的关键问题】选取长势一致的3株核桃树在内堂空虚的主枝中下部位进行环剥,萌发新枝后进行嫁接,并选取三株完整核桃树作为对照,利用Li-6400便携式光合分析仪测定核桃在17不同光量子通量密度下的光响应曲线,研究核桃主枝环剥之后上、下部位核桃叶片对光能利用的差异,为新疆核桃提质增效提供技术支撑。
1 材料与方法
1.1 材 料
试验地选择在新疆阿克苏地区温宿县佳木镇,地处天山中部托木尔峰南麓,塔里木盆地西北的边缘,属于大陆性干旱荒漠气候,年降水稀少,季节分配不均,昼夜温差大,春季升温快而不稳,秋季降温迅速,1年四季气候干燥,光照充足。春天多以沙尘天气为主,夏季的强对流天气易于发生冰雹、暴雨等。年均温10℃左右,年均降水量65.4 mm,年均日照时数2 747.7 h,年均无霜期185 d,土壤以沙土为主,质地适中,呈弱碱性。
1.2 方 法
供试材料新温185 6年生核桃树,在4月上旬,选取长势一致的3株核桃树在内堂空虚的主枝中下部位进行环剥,宽度0.5 cm,待其萌发新枝后,选取3株完整核桃树作为对照,分别以环剥上部、下部(图/A),未环剥(图/B)为1次处理,共3次处理。图1
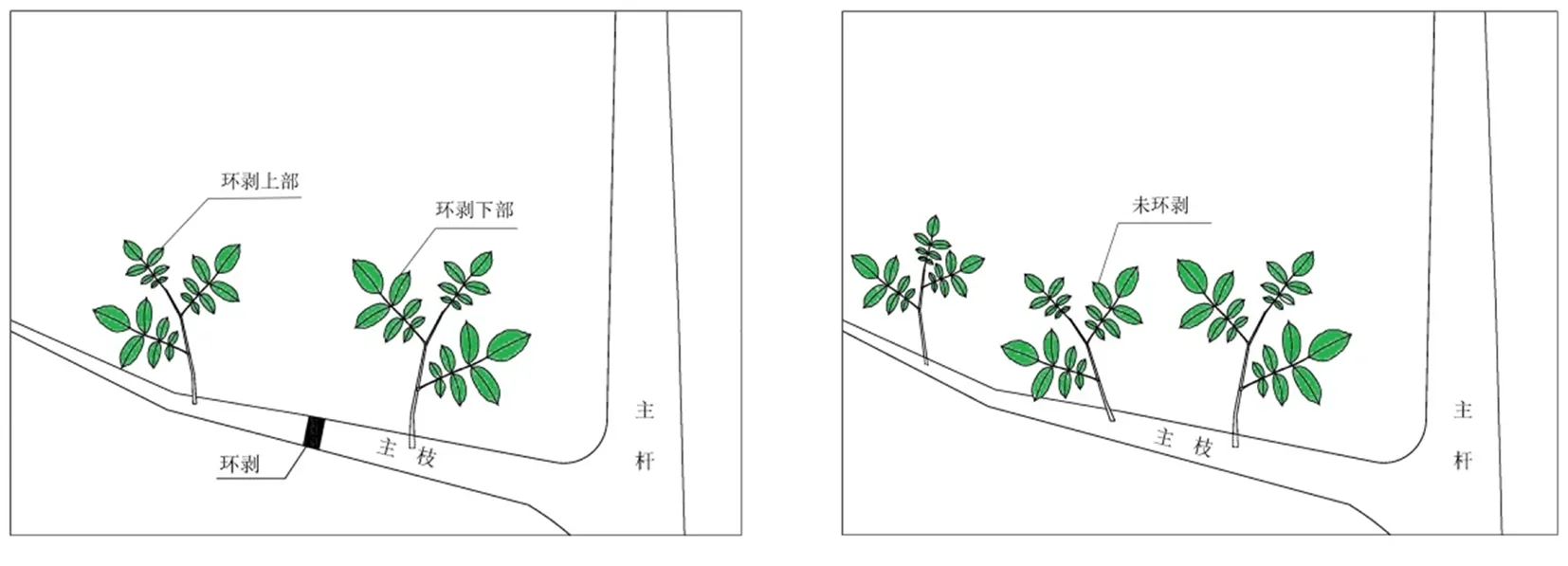
图1 环剥示意
Fig. 1 Sculpture diagram
试验于2019年6月晴朗的天气在10:00~11:30。运用美国Li-6400便携式光合分析仪进行光响应测定,供试核桃活体叶样为17个(0、20、50、100、150、200、250、300、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000)不同光量子通量密度(photosynthetic photo flux density,PPFD),分别于6月9日、6月16日、6月23日对每个处理选取3片叶样完整无损,且光照充足的成熟叶片进行光响应参数测定,叶室选择红蓝LED光源,测定不同叶片在不同光量子梯度下的净光合速率(Pn,μmol/(m2·s))、光合有效辐射(PAR,μmol/(m2·s))、胞间CO2浓度(Ci,μmol/(m2s))、气孔导度(Gs,μmol/(m2·s))、蒸腾速率(Tr,μmol/(m2·s))、叶片温度(Tleaf,℃)等生理参数,最终得到不同叶片的光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Amax)、暗呼吸速率(Rd)、以及表观量子效率(AQY)、叶片瞬时水分利用效率(WUE)等参数。
叶片瞬时水分利用效率 (WUE, μmol/(m2·s)) 的计算公式为:WUE=Pn/Tr.
1.3 数据处理
采用Excel 2010以及SPASS17.0处理试验中所的数据。
2 结果与分析
2.1 不同部位核桃主枝环剥之后叶片的光响应曲线特征参数
研究表明,3种不同部位叶片的光响应曲线特征参数之间存在较大差别,未环剥核桃树的叶片Amax、LSP、LCP、Rd值在3种方式中最大,环剥下部叶片AQY值最大。因为Amax反应的是植物叶片的最大光合能力,即未环剥核桃叶片的光合能力最大,其次是环剥下部,最后为环剥上部的叶片。AQY反映的是植物在弱光条件下的光合能力,Rd与植物叶片的生理活动有关,环剥核桃上部与下部叶片的光合与未环剥核桃相比较,未环剥核桃叶片的光合能力最大,生理活性最强,对PAR的反应能力最强。表1
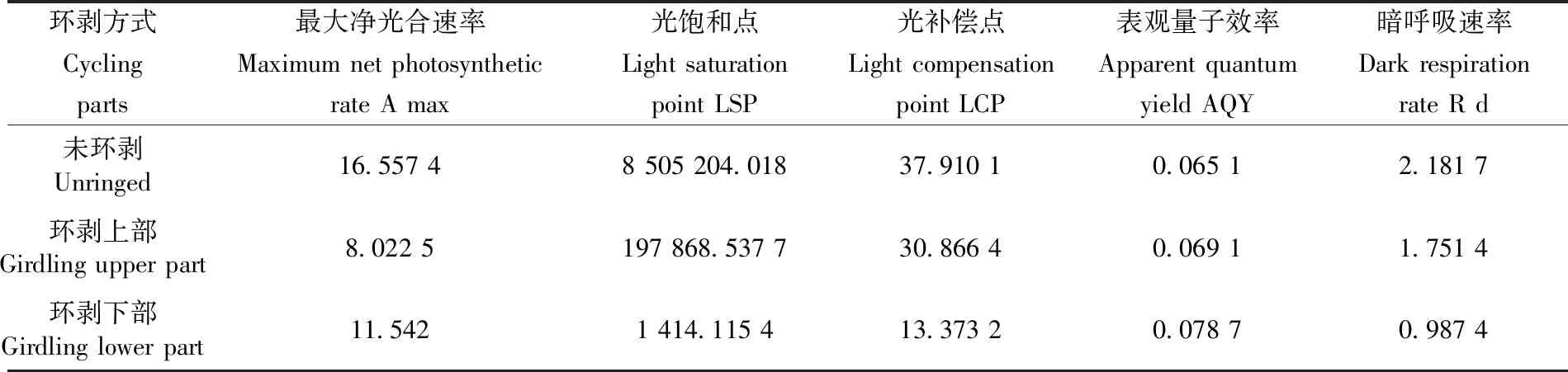
表1 3种不同部位核桃主枝环剥光响应曲线特征参数Table 1 Characteristics of the walnut-carved bud response curve in three different parts
2.2 不同部位核桃主枝环剥光合速率与光强的变化关系曲线
研究表明,3种不同部位核桃主枝环剥的Pn均随PAR的增大而增大,但是达到一定程度时则出现下降状态,并且一开始增幅较大,随之慢慢变小。环剥上部叶片的光合最小且在3种环剥方式中所有值均为最低,其中Pn的Amax为8.022 5,LSP为197 868.537 7,LCP为30.866 4,Rd为1.751 4,这说明环剥上部的光合能力相对较弱,未环剥与环剥下部的PAR在0~600时,环剥下部的Pn值高于未环剥的值,自600 μmol/(m2·s)开始,环剥下部的值开始低于未环剥的值且未环剥逐渐开始趋于平缓并呈下降趋势,而未环剥的Pn值随PAR的增大依然呈上升趋势至2 000左右开始呈现下降趋势,其中未环剥的Amax为16.557 4,LSP为8 505 204.018,LCP为37.910 1,Rd为2.181 7;而环剥下部的Amax为11.542,LSP为1 414.115 4,LCP为13.373 2,Rd为0.987。图2
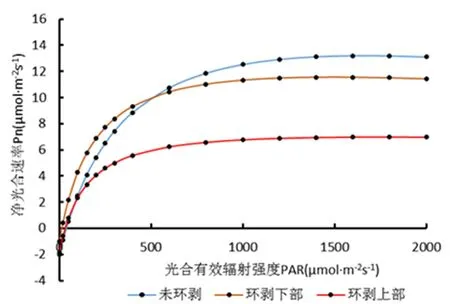
图2 不同部位核桃主枝环剥光合速率与光强的变化关系曲线
Fig. 2 Chart of the relationship between the rate of photosynthesisof walnut inscribed buds and the light strength of different parts
2.3 不同部位核桃主枝环剥Gs、Tr、Ci、Tleaf、WUE与光强的变化关系曲线
研究表明,随着PAR强度的增大,3种部位Gs、Tr的值均呈上升趋势,且Gs环剥下部>Gs未环剥>Gs环剥上部值,Tr环剥下部>Tr未环剥>Tr环剥上部值。胞间CO2浓度值,随PAR强度的增大而减小至一定程度后有略微的上升,PAR增加至1 000 μmol/(m2·s)时,胞间CO2浓度值最低,之后出现略微上升趋势。在弱光条件下Pn的值持续加快上升,而Gs上升较缓慢,气孔出现限制现象,使得CO2的供应无法满足Pn的需求,导致Ci值急剧下降。在光反应后期,对气孔刺激度增强,使得气孔发生较大程度开放,后期Gs 增幅变大;在气孔大程度开放以后,CO2供应充足,Ci 出现升高趋势。环剥不同部位Tleaf值,变化具有差异性,环剥下部与环剥上部的Tleaf值均随PAR强度的增加持续增加,但未环剥的Tleaf值光强达到800 μmol/(m2·s)后开始出现下降趋势,到1 400 μmol/(m2·s)时低于环剥下部的Tleaf值。各种部位叶片的生理活性对PAR的反应均有所不同,环剥上部的各种指标均是最低的,而环剥下部的和未环剥的均数值较高,原因可能是环剥上部由于环剥原因使得营养养分由木质部向上运输时由于核桃导管刻伤而将营养养分截留下来,未能运输至环剥上部,使得核桃养分补充不及时,最终导致各项指标均最低。3种不同部位的WUE表现趋势均,随着PAR 的增强而迅速上升而后缓慢下降,WUE主要取决于植物的光合速率与蒸腾速率,光强在400 μmol/(m2·s)之前环剥下部的WUE比环剥下部和未环剥的光反应均敏感,光强在400 μmol/(m2·s)之后WUE值未环剥>环剥上部>环剥下部。图3
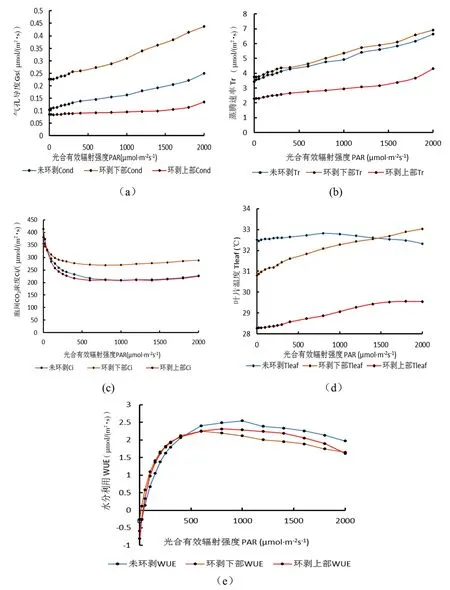
图3 不同部位核桃主枝环剥Gs、Tr、Ci、Tleaf、WUE与光强的变化关系曲线
Fig. 3 The relationship between walnut-carvedGs,Tr,Ci, Tleaf, WUE and light strength in different parts
3 讨 论
Amax是反应植物进行光合作用的一项重要指标,在对环剥之后核桃不同部位的叶片进行光合光响应测定后,发现各个部位的Amax都存在着较大差异。其中Pn最高的为未环剥核桃叶片,其次为环剥下部,最后为环剥上部,说明核桃叶片的光合作用未环剥>环剥下部>环剥上部。在光响应特性指标中有两个各重要的植物光合生理指标LSP和LCP,反映的是植物对光照强度的要求,若LSP与LCP值均较低则表明植物对弱光利用较强,若LSP与LCP值均较高表明植物生长发育对强光利用较强,若LSP值较高而LCP值较低表明植物对光环境的适应性较强,相反,若LSP值较低而LCP值较高,则表明植物对光环境的适应性相对较弱[10]。对环剥之后核桃不同部位的叶片进行光合光响应测定,发现各个部位的光合生理参数都存在着较大差异。未环剥与环剥上部的LSP与LCP值均高于环剥下部的,表明未环剥与环剥上部的核桃叶片对强光的适应性比环剥下部的强。AQY反映的是植物在弱光条件下的光合能力,而AQY值,环剥下部>未环剥>环剥上部,则表明环剥下部的核桃叶片比未环剥与环剥上部的核桃叶片对弱光的利用率更强表明环剥下部在树体下部的光和反应依然是较好的。前人在光合研究中多偏向于光合对果实品质的研究,却对于核桃主枝环剥嫁接研究并不多,对核桃可通过芽、进行嫁接的条件下对核桃的叶片进行光合测定,研究环剥嫁接[20]对核桃生长发育的影响,提高嫁接成活率、实现提质增效具有一定的参考价值。
4 结 论
4.1 核桃主枝环剥之后不同部位Pn未环剥>环剥下部>环剥上部,未环剥核桃光合能力最强,环剥之后环剥下部高于环剥上部,环剥下部相比较环剥上部,适宜嫁接、改造。
4.2 核桃主枝环剥之后不同部位AQY值,环剥下部>未环剥>环剥上部,环剥下部的核桃叶片比未环剥与环剥上部的核桃叶片对弱光的利用率更强,表明环剥下部在树体下部的光合反应最好,所以此部分枝条是适宜进行嫁接、栽培的。
猜你喜欢
北方果树(2022年2期)2022-11-10
北方果树(2022年3期)2022-11-10
江西通信科技(2022年2期)2022-08-08
福建农林大学学报(自然科学版)(2021年5期)2021-10-08
落叶果树(2021年1期)2021-01-29
农家致富顾问·上半月(2020年1期)2020-08-10
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
福建农业科技(2016年10期)2016-03-07
少儿科学周刊·儿童版(2015年10期)2015-11-07
