
人SIRT6蛋白的原核表达及纯化
2019-02-28 03:26罗沙柳任鑫鑫罗菲梁迎春马超刘坤冉芳施亚娇李玲叶棋浓
生物技术通讯 2019年6期
罗沙柳,任鑫鑫,罗菲,梁迎春,马超,刘坤,冉芳,施亚娇,李玲,叶棋浓
1.贵州大学 医学院,贵州 贵阳550025;2.军事科学院 军事医学研究院 生物工程研究所,北京100850;3.山西医科大学 第二医院,山西 太原030000;4.大连医科大学 肿瘤干细胞研究院,辽宁 大连,116044;5.中部战区总医院 卫勤部科训科,湖北 武汉430072
Sirtuins(SIRT)家族中的沉默信息调节因子2(silent information regulator 2,Sir2)最早在酵母中被发现[1]。迄今,科学家们在哺乳动物中已经确定 了7种与Sir2 同 源的基因,即SIRT1~SIRT7[2]。人类的SIRT6基因位于染色体19P13.3 区域,包括8个外显子,编码的蛋白全长355个氨基酸残基,相对分子质量39×103,等电点9.12[3]。SIRT6 最初被认为是单-ADP-核糖基转移酶,可以通过分子内机制完成放射性标记的转移,这表明SIRT6 可能利用ADP-核糖基化来自动调节自身活性,但是其最主要作用还是作为组蛋白脱乙酰酶[4-9]。
目前研究证明,SIRT6在基因组稳定性、代谢、染色质调控、端粒完整性、基因转录和糖脂代谢等方面都起关键调控作用,调节糖尿病、心脏病、肥胖、癌症及寿命、衰老等相关病理生理的发生发展[10-12]。SIRT6在体内控制许多代谢途径,包括糖酵解、糖异生、甘油三酯合成和LDL-胆固醇稳态,SIRT6 缺乏导致主要代谢缺陷[13-16]。对于癌症,SIRT6 可以通过多种机制发挥抑癌作用,敲除SIRT6的小鼠胚胎成纤维细胞(MEF)比野生型MEF细胞生长增殖快,且敲除SIRT6 有促进肿瘤发生的作用[13];敲除SIRT6基因的肿瘤细胞的乳酸生成、葡萄糖摄取等均增加[17]。对116例乳腺癌病人的组化分析显示SIRT6 高表达的病人生存预后差,SIRT6 通过调控FOXO的表达降低肿瘤细胞对化疗药的敏感性;在前列腺肿瘤组织和前列腺癌细胞中SIRT6表达都高于正常或癌旁组织,且敲低SIRT6 降低细胞活力,增加药物的敏感性等;这又从另外一个层面说明SIRT6 发挥致癌作用[17-20]。有文献报道SIRT6 能与缺氧诱导因子HIF1α相互作用[17],从而调控肿瘤的代谢重编程。
在本研究中,我们构建了带GST 标签的人SIRT6基因原核表达载体,表达纯化获得重组GST-SIRT6 蛋白,验证了SIRT6与HIF1α之间的关系,有助于进一步探讨SIRT6在肿瘤发生发展中的调控机制。
1 材料和方法
1.1 材料
大肠杆菌DH5α、BL21(DE3)感受态细胞购自北京博迈德科技发展有限公司;人乳腺文库、带GST 标签的pGEX-KG 原核表达载体为本室保存;PCR 试剂、限制性内切酶及DNA 连接酶购自TaKaRa 公司;质粒提取、胶回收试剂盒购自Pro⁃mega 公司;GST-Sepharose 4B 珠购自Pharmacia 公司;HRP 标记的抗GST 单克隆抗体购自GE Healthcare Life Sciences 公司;引物合成及测序由北京博迈德科技发展有限公司完成。
1.2 人SIRT6基因编码区的获取
根据GenBank 中的人源SIRT6基因编码序列设计上游引物5'-CGGGATCCATGTCGGTGAATT ACGCGGCGGGG-3'和下游引物,5'-CCGCTCGAG TCAGCTGGGGACCGCCTTGGCCT-3'。以本室保存的乳腺文库为模板,用Primer Star DNA 聚合酶扩增目的基因片段(PCR 程序:95℃预变性5 min,95℃变性30 s,60℃退火30 s,72℃延伸75 s,共35个循环,72℃再延伸5 min)。琼脂糖胶检测PCR 产物,切出目的基因片段,用胶回收试剂盒回收目的片段。
1.3 GST-SIRT6 重组质粒的构建及鉴定
将回收的目的基因片段及载体GST 分别用限制性内切酶BamHⅠ/XhoⅠ于37℃双酶切4~6 h 或过夜,琼脂糖凝胶电泳回收双酶切后的PCR 产物及载体。将双酶切的PCR 产物及载体用T4DNA连接酶于16℃连接6 h 以上后转化大肠杆菌DH5α感受态,挑菌进行菌液PCR 鉴定,将鉴定得到的阳性克隆用质粒提取试剂盒提取质粒,双酶切鉴定,并送北京博迈德生物公司测序。
1.4 重组质粒GST-SIRT6的诱导及表达鉴定
将测序正确的GST-SIRT6 重组质粒转化大肠杆菌BL21 感受态并涂于含氨苄青霉素的LB 板上,37℃恒温箱中培养过夜,挑取单菌落于装有5 mL 含氨苄青霉素的LB 培养基的10 mL 试管中,于37℃、200 r/min 摇床上振荡培养过夜,按1∶50 将菌液稀释,30℃培养至D600nm约为0.6,用终浓度1 mmol/L的IPTG 小量诱导,调定温度至20℃继续振荡培养4~6 h。收集诱导前、后的菌液,裂解后行Western 印迹,鉴定有无SIRT6 蛋白表达。
1.5 GST-SIRT6 融合蛋白的纯化
挑选出能诱导GST-SIRT6 蛋白的重组大肠杆菌BL21 接种于5 mL 含氨苄青霉素的LB 中,37℃培养过夜,将菌液转移到300 mL 含氨苄青霉素的LB 中,30℃、200 r/min 振荡培养4~6 h,直至菌液D600nm为1.0~1.5,再 按1∶10 000的比例 加 入IPTG,20℃、200 r/min 培养16~24 h。离心收集菌体沉淀,加入裂解液重悬,冰浴30 min 使菌体充分裂解,再用超声波裂解细菌,收集上清液,加入GST-Sepharose 4B 纯化珠,4℃旋转结合4 h 或过夜,离心收集结合蛋白后的GST-Sepharose 4B 纯化珠,缓冲液洗脱未结合的蛋白后得到纯化的融合蛋白,行SDS-PAGE,考马斯亮蓝染色鉴定。
1.6 GST-pulldown 实验
用5 μg MYC-HIF1α真核表达载体分别转染2个6 cm 皿的293T细胞系,24~48 h后收细胞,超声波裂解半小时,4℃、12 000 r/min 离心5 min,取25 μL 上清,加入等量2×SDS 上样缓冲液作为结合前的蛋白样,将剩余上清与已纯化的GST、GST-SIRT6 蛋白珠于4℃结合4~6 h,再用IP缓冲液洗涤蛋白珠,弃上清,加入2×SDS 上样缓冲液,Western 印迹检测。
2 结果
2.1 GST-SIRT6 重组质粒的构建与鉴定
以本实验室保存的人乳腺文库为模板,PCR扩增人SIRT6基因的编码序列,获得1059 bp的DNA片段,与预期大小一致(图1)。将BamHⅠ/XhoⅠ酶切后的PCR 产物与GST 载体连接,转化大肠杆菌DH5α感受态,挑菌后进行菌液PCR 鉴定,获得与目的条带1059 bp 大小接近(图2)的克隆,初步认定是带有人源SIRT6基因的阳性重组克隆。选取阳性克隆提质粒,酶切鉴定,可切出长度分别约为5000和1059 bp的条带,而相应的空载体酶切后只见大条带,与预期结果相符(图3)。重组质粒测序结果表明,插入的DNA片段序列与人SIRT6基因的编码序列一致(序列略)。
2.2 GST-SIRT6 重组质粒的表达鉴定
分别用HRP 标记的抗GST 单克隆进行West⁃ern 印迹,结果见图4。泳道4 相对分子质量68×103左右有抗体特异结合条带,即GST-SIRT6 融合蛋白(谷胱甘肽转移酶GST 相对分子质量26×103左右,SIRT6 为42×103左右)。泳道4 相对分子质量26×103~42×103是蛋白降解带。结果与预期一致,说明GST-SIRT6 融合蛋白表达正确。
2.3 融合蛋白GST-SIRT6的纯化
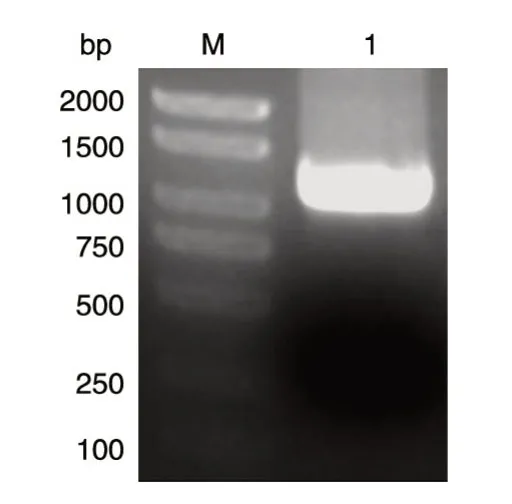
图1 PCR 扩增人SIRT6的编码序列
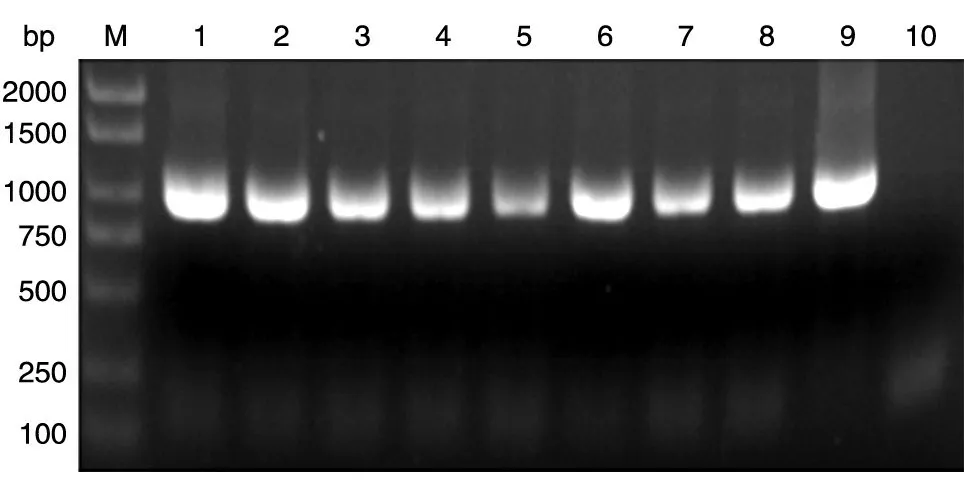
图2 重组质粒GST-SIRT6的菌液PCR 结果电泳图谱
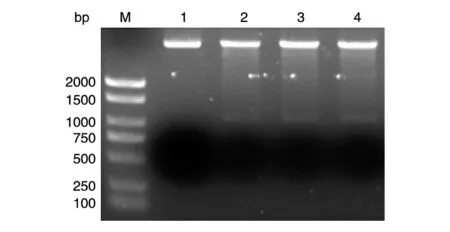
图3 重组质粒GST-SIRT6的BamHⅠ/XhoⅠ双酶切电泳图谱
利用GST-Sepharose 4B 亲和珠对GST-SIRT6进行纯化,行SDS-PAGE,考马斯亮蓝染色,泳道2可见相应蛋白条带出现(图5)。
2.4 GST-pulldown 实验
将MYC-HIF1α转染293T细胞,所提取的蛋白和纯化的GST、GST-SIRT6 蛋白珠结合,进行SDS-PAGE,分别用MYC-HRP和GST-HRP 抗体检测,结果如图6。结合后孵育MYC-HRP 抗体,泳道2 出现特异蛋白条带,证明MYC-HIF1α与GSTSIRT6 融合蛋白有相互作用。
3 讨论
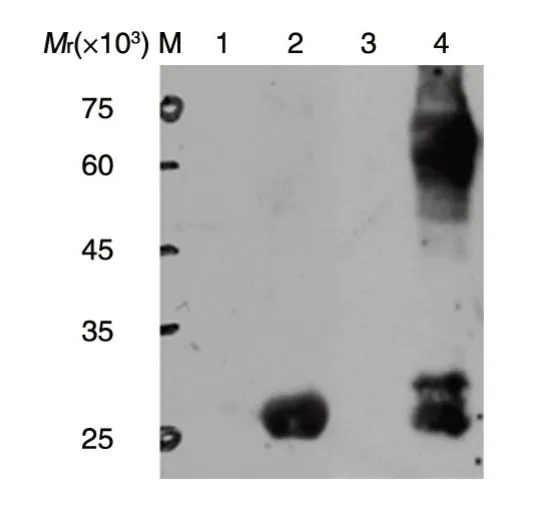
图4 Western 印迹检测融合蛋白的表达
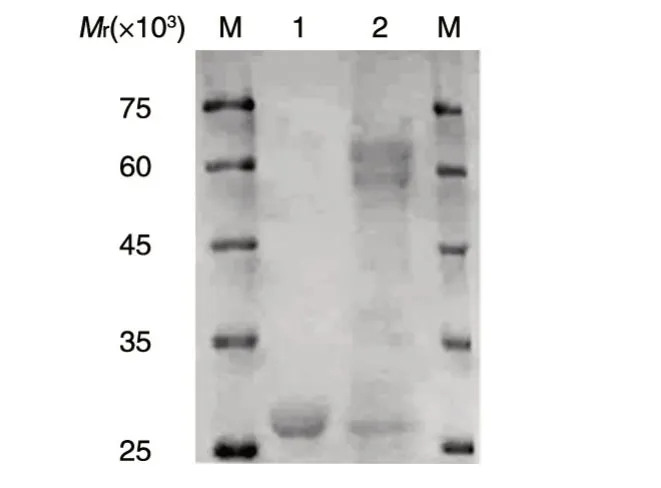
图5 重组蛋白GST-SIRT6的纯化
目前研究发现SIRT6在肿瘤中的作用具有争议。SIRT6 最近被证明是一种明显的肿瘤抑制因子,在人类多种癌症中SIRT6 蛋白水平偏低,如在肝癌、胰腺癌、卵巢癌和结直肠癌等中都下调或突变[18-20]。在卵巢癌中,SIRT6 通过降低Notch3 抑制肿瘤增殖[21]。SIRT6 还通过控制Lin28b 抑制胰腺癌的发生发展。然而,SIRT6在前列腺癌、皮肤癌和乳腺癌等其他一些癌症类型中高表达,并作为癌基因发挥作用,在小细胞肺癌中也有类似的现象[22]。在癌症发生发展中,增加蛋白质合成对癌细胞的存活、增殖、发展和转化至关重要[23-24]。Ravi 等发现SIRT6 可以通过与转录因子SP1 结合,一起转录调控mTOR 信号通路,从而调控蛋白质合成来影响肿瘤的发生发展[25]。另有报道,SIRT6通过ERK1/2/MMP9 通路调控骨肉瘤的迁移和侵袭[26]。SIRT6 通过抑制糖酵解相关基因的表达来调节葡萄糖代谢的稳态。SIRT6 通过在HIF1α靶基因启动子上使H3K9 脱乙酰化而起到HIF1α转录活性的共抑制剂的作用,当SIRT6 失活时,HIF1α被激活,糖酵解基因启动子的乙酰化与多个代谢基因的表达增加,并最终导致糖酵解的增加和线粒体呼吸的减少[17]。SIRT6与SIRT1 直接结合后可以去乙酰化hnRNPA1,从而抑制肝癌细胞糖酵解和增殖[27]。综上所述,SIRT6在肿瘤的发生发展中都起到了关键作用。
SIRT6 通过一些酶反应尤其是去乙酰化,可以模拟染色质在致密状态(如SREBP1和PCSK9)来抑制基因表达[28]。有报道发现SIRT6 可以通过HIF1α来调控Pdk1、Ldha、Pfkl1 等糖酵解相关基因[13],从而调控肿瘤代谢重编程[17]。在此我们进一步验证了SIRT6与HIF1α在体外有相互作用,为后续研究SIRT6与HIF1α在肿瘤中对肿瘤糖代谢的影响进而影响肿瘤的发生发展奠定了基础。
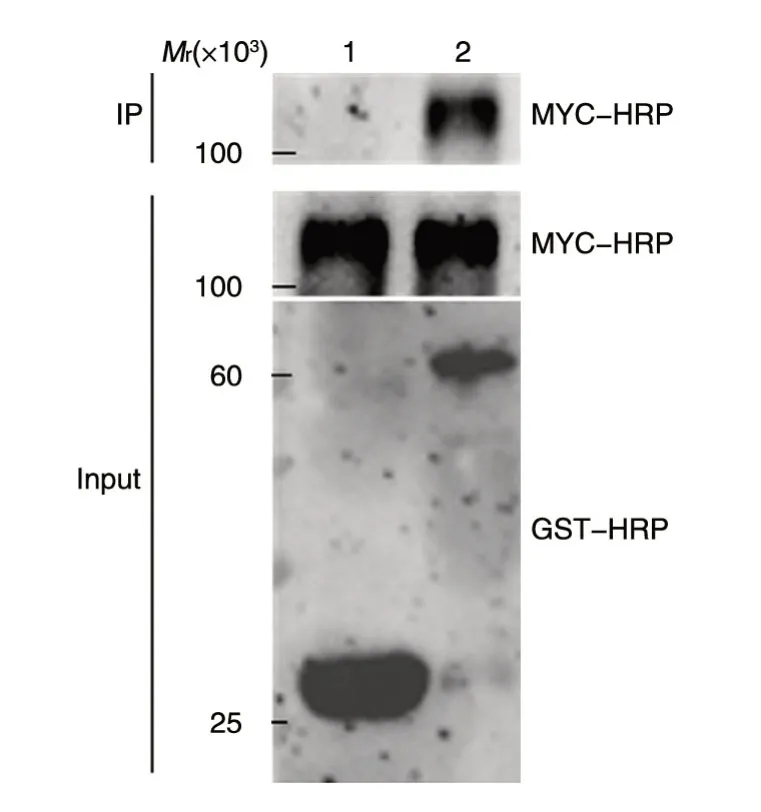
图6 GST-pulldown 验证GST-SIRT6与MYC-HIF1α的体外相互作用
本研究构建了带GST 标签的SIRT6 原核表达载体,并在IPTG诱导下表达了GST-SIRT6 融合蛋白,Western 印迹鉴定证实了目的蛋白的表达。该重组质粒的构建,为后续研究SIRT6在肿瘤中的发生发展制备了材料。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
江西农业学报(2021年4期)2021-04-20
水生生物学报(2021年1期)2021-02-04
疯狂英语·新悦读(2020年7期)2020-07-30
食品科学(2020年4期)2020-03-11
癌变·畸变·突变(2020年1期)2020-02-12
中国循证心血管医学杂志(2020年11期)2020-01-08
中成药(2019年12期)2020-01-04
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医学装备(2016年6期)2016-12-01
