
广东省中山市五桂山土沉香空间遗传结构
2019-02-13 06:20刘盼盼廖浩斌蒋谦才谭宗健孙红梅王瑞江曹洪麟黄向旭赖思茹梁丹王峥峰
热带亚热带植物学报 2019年1期
刘盼盼, 廖浩斌, 蒋谦才, 谭宗健, 孙红梅, 王瑞江, 曹洪麟, 黄向旭, 赖思茹, 梁丹, 王峥峰*
广东省中山市五桂山土沉香空间遗传结构
刘盼盼1, 廖浩斌1, 蒋谦才1, 谭宗健1, 孙红梅1, 王瑞江2, 曹洪麟2, 黄向旭2, 赖思茹2, 梁丹2, 王峥峰2*
(1. 中山市国有森林资源保护中心,广东 中山 528400;2. 中国科学院华南植物园, 广东省应用植物学重点实验室,广州 510650)
为了解五桂山的土沉香()种群空间遗传结构,利用18个微卫星位点对143株胸高直径大于5 cm的土沉香个体基因型进行了检测。结果表明,五桂山土沉香种群的观测杂合度和期望杂合度均为0.534;近交系数为0.000 1,说明五桂山土沉香遗传多样性并不低,且种群处于随机交配状况。空间遗传结构分析结果表明,五桂山土沉香缺乏空间遗传结构,不同遗传背景的个体相互混杂,导致相邻个体遗传差异较大。这可能是五桂山土沉香种群发育过程中自疏所致,也可能是这一地区土沉香为人工种植,而非自然发育形成而造成的。
土沉香;微卫星位点; 遗传多样性; 遗传景观图; 空间主成分分析法; 空间自相关法
土沉香[(Lour.) Gilg],又称白木香、香树、女儿香、牙香树,为瑞香科(Thyme- laeaceae)沉香属植物,分布于我国热带、亚热带地区,为多年生常绿乔木,中国特有。土沉香是一种珍贵的香料和中药资源[1]。作为香料,自古以来就有“沉檀龙麝”之说,其中的“沉”,就是指沉香,其香品高雅,被列为众香之首。作为药材,其功效主要有理气调中、壮阳除痹、行气止痛和纳气平喘,被列为十大广药之一。
土沉香浑身都是宝,用途非常广泛。除了作为名贵药材和天然香料,其种子可制肥皂、润滑油,有重要的工业用途;树皮可用来造纸和生产人造棉。同时,沉香树为主根向下生长之深根系弱阳性树种,是最好的水土保护者。沉香生长过程中,不需喷洒农药,不会危害土质和人体的健康,有益于生态维护。沉香又是中国悠久历史文化的产物,它起源于唐之前,经历各朝代,从香案、香囊、香袋、香木雕品及薰香等,处处体现了土沉香包含的文化底蕴。因此,土沉香具有十分重要的科研价值、经济价值、生态价值和文化价值。
由于沉香的重要经济价值,致使其野生资源被掠夺性采伐,破坏严重。为保护这一珍贵野生种质资源,土沉香被《国家重点保护野生植物名录》(1999)列为国家二级保护植物,同时被《濒危野生动植物种国际贸易公约(CITES)》列为禁止贸易的物种资源。
土沉香的种植与沉香文化具有悠久的历史,源远流长。广东省中山市是土沉香种植和文化产业的重要区域。中山市古称香山,地属东莞县文顺乡, 这一地区出产的沉香被称为“莞香”,是上乘进贡极品,因而中山市享有“中国沉香之乡”美誉[2–3]。作为古代莞香主要产地,中山市五桂山地区土壤和气候条件俱佳,至今仍遗留有丰富的土沉香种质资源,极具保护和利用价值[4]。
遗传多样性是生物多样性的重要组成,是物种适应环境和进化发展的源泉。因此,了解、保护和维持物种的遗传多样性具有十分重要的理论和实践意义。空间遗传结构是物种遗传多样性在空间分布的状况,是物种生活史特征(交配系统、扩散)和环境因素(种间竞争、生境选择)多方面作用下形成的特定结构形式,在反映物种进化历史、生境适应方面具有重要作用[5–7]。
为此,本文在前期五桂山地区土沉香遗传多样性研究[8]的基础上,通过增加采样量,对中山市五桂山的土沉香空间遗传结构进行研究,为这一地区的土沉香种质资源保护和开发提供相关指导。
1 材料和方法
1.1 研究地点和样品采集
土沉香[(Lour.) Gilg]采样地点位于广东省中山市五桂山生态保护区内[8], 在非常熟悉五桂山保护区内土沉香分布的当地护林员帮助下,沿着林中小路进行样品采集,基本覆盖土沉香主要分布区,对胸径(diameter at breast height, DBH)大于5 cm的土沉香个体,采集2~3片叶,放入装有硅胶的密封袋内带回实验室处理,所采集的土沉香植株数量占现有分布数量的90%以上。野外观察表明,DBH大于5 cm的土沉香已可以开花结实。对每株采样个体进行GPS定位(图1),并测定DBH, 共采集143株土沉香的叶片,DBH主要为10~30 cm (图2)。为了解年龄级对空间遗传结构的影响,按照土沉香胸高直径分布,把样株分成2个年龄级:年龄级I,胸高直径小于20 cm (76株); 年龄级II,胸高直径大于20 cm (67株)。
1.2 遗传多样性检测
参考蒋谦才等[8]的18个微卫星位点和方法对所有土沉香进行基因型分析。
1.3 数据分析
利用GenAlEx 6.501[9]和Genepop-4.5.1[10]软件分别计算五桂山土沉香各位点和种群等位基因数、观测杂合度和期望杂合度和近交系数()[11]。采用Bonferroni纠正的方法进行多重比较下的显著性水平调整[12]。其中,近交系数值反映位点或种群是否处于哈迪-温伯格平衡(Hardy-Weinbergequilibrium,一种自由交配状况),值为0表明位点或种群处于平衡状态。
空间遗传结构分析首先采用遗传景观图(genetic landscape shapes, GLS)方法[13], 先构建所有个体间的德劳内三角网格图,然后对每个网格图计算个体间的遗传距离,最后应用统计学方法把每个网格对应的遗传距离反映在空间格局上,反映局部空间个体间的相似性。
再采用Jombart等[14]开发的sPCA (spatial principal component analysis)方法对数据进行分析, 其整合了空间和主成分分析(principle component analysis, PCA)方法,可有效揭示不同尺度空间上个体间的遗传相似程度。sPCA分析同时可以对个体在空间上的正相关和负相关进行显著程度的检测, 正相关被称为全局检验,负相关被称为局部检验。两者的显著度检验采用对比999次随机重复结果获得。
最后利用GenAlEx 6.501进行遗传多样性的空间自相关分析(spatial autocorrelation analysis),探讨个体间的遗传相似性随地理距离变化的状况。由于个体间地理距离变化较大(0.9~1 638.3 m),采用非均匀地理尺度,包括了大小不同地理尺度,以全面了解不同尺度下五桂山土沉香空间自相关状况。
2 结果和分析
18个微卫星位点在五桂山土沉香种群中分别扩增出2~14个等位基因,观测杂合度为0.063~ 0.797,期望杂合度为0.075~0.802 (表1),近交系数表明所有位点均符合哈迪-温伯格平衡。种群整体观测杂合度和期望杂合度都为0.534,近交系数也接近0,表明种群整体处于哈迪-温伯格平衡。
从年龄级来看,年龄级I的整体观测杂合度为0.526,期望杂合度为0.530;年龄级II的整体观测杂合度为0.542,期望杂合度为0.539。两个年龄级的近交系数分别为0.0075和-0.0070,都接近0。
GLS分析结果表明,五桂山土沉香只有东北和西南部小部分个体间有较小的遗传距离(图3)。不同年龄级的GLS结果(图略)均和整体结果一致。
根据sPCA计算的特征向量值(图4),前两个正向量分别解释了10.380%和6.682%的遗传变异。从第一向量来看(图5: A),除东北方向的土沉香个体间遗传相似性较大,形成比较单纯的聚集外,其他地区没有明显的遗传相似度较大的个体间形成地理聚集状况,并出现遗传差异较大的个体在同一地区混杂;第二向量(图5: B)也表明遗传差异较大的个体相互混杂分布,没有明显的地理聚集群。正相关和负相关显著度检测值()分别为0.094和0.907,表明不存在明显的空间相关性。不同年龄级第一向量和第二向量中的第一轴和第二轴均反映出不同遗传差异个体的混杂(图略),与整体分析结果一致。
空间自相关分析结果表明,五桂山土沉香整体上缺乏空间遗传结构,在不同空间尺度上表现为随机状况(图6)。两个年龄级也均表现出缺乏空间遗传结构的结果(图略)。
3 讨论
我们前期对中山市五桂山土沉香遗传多样性进行了初步研究,观察杂合度和期望杂合度分别为0.523和0.522[8],本次增加了采集量,基本覆盖了五桂山土沉香生长分布区。相比之下,随着样品量的增加,整体遗传多样性略有增加,观察杂合度和期望杂合度都为0.534。但两次的近交系数都接近0,表明五桂山土沉香种群整体处于随机交配状况。
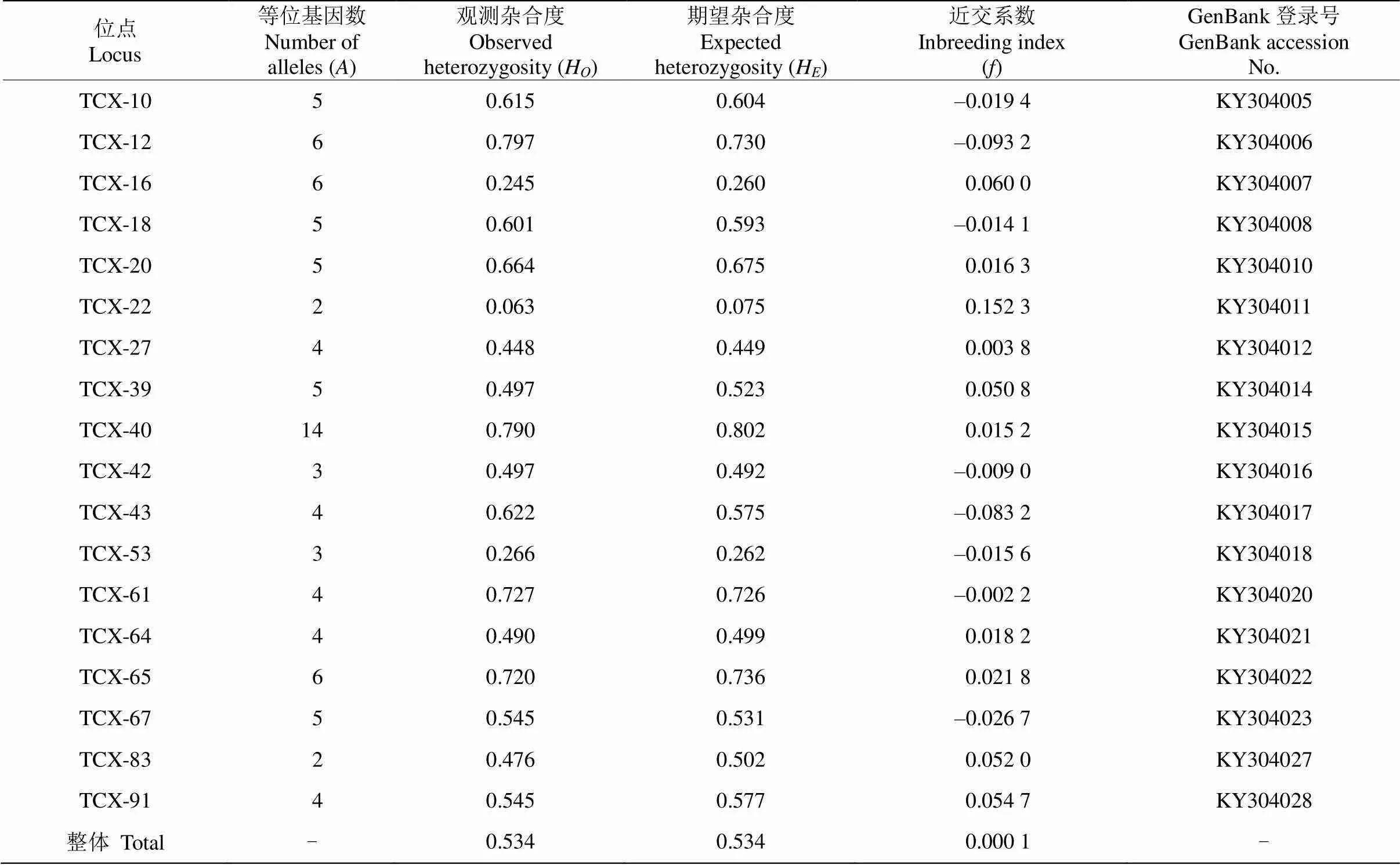
表1 土沉香18个微卫星位点遗传多样性
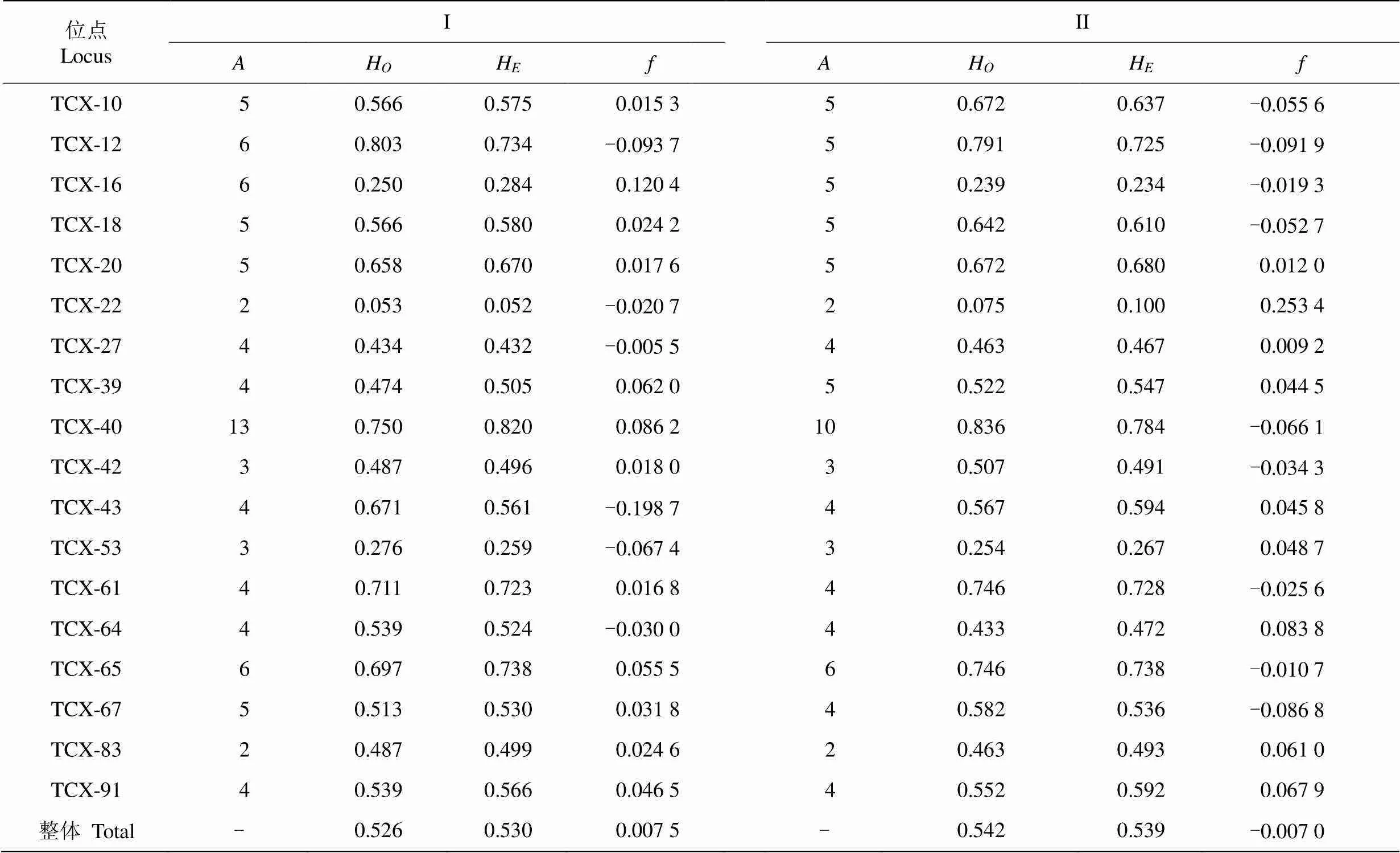
表2 五桂山土沉香两个年龄级的遗传多样性
: 等位基因数;H: 观测杂合度;H: 期望杂合度;: 近交系数。
: Numble of alleles;H: Observed heterozygosity;H: Expected heterozygosity;: Inbreeding index.
除了遗传多样性,空间遗传结构也是反映种群维持状况的一个重要方面。遗传多样性大小是物种生活史特征及其进化历史过程的综合反映,其大小与物种的适应进化、濒危状况等有着一定的相关性[15–16]。但物种由不同地理种群组成,而种群又是由不同个体组成。这些种群或者个体分布在不同地理位置上,所处的景观格局、生境状况可能千差万别,不同的生物和非生物因子作用于不同的种群或个体上,会造成不同地理来源、分布点的种群或个体遗传变异的差异。因此,仅仅进行物种或种群遗传多样性大小的研究不能全面把握物种遗传变异状况。而空间遗传结构作为了解遗传变异与种群或个体地理位置相关状况,有助于更深入探讨物种遗传多样性形成,为物种的合理保护和经营提供指导。对于植物而言,花粉和种子扩散状况是决定其空间遗传结构的首要方面,其中种子扩散更具有主导性[7]。
土沉香是虫媒花,传粉者主要是蛾类,如螟蛾(Pyralids)和夜蛾(Noctuids)[17]。虽然这些昆虫在土沉香中的传粉距离还没有研究,但有研究表明这些昆虫的传粉距离可能较长(1 000 m或更远)[18–21]。但由于虫媒花需要昆虫等的帮助,花粉流状况有更多不确定性, 如受昆虫种群大小、昆虫本身运动能力、景观格局的影响[21–23]。土沉香的种子为卵圆形,黑褐色,长度约14 mm,宽度约5 mm,重约0.14 g[24], 除种子本体还附着一附属物,附属物可以分泌化学信号,引诱多种胡蜂前来取食, 胡蜂取食过程中一定程度上会协助土沉香种子传播。但在野外观察,能被胡蜂成功携带远距离传播的种子很有限,大部分种子还是依靠重力传播,散落在母树周围,因此土沉香的种子有重力和摄食传播两种方式[1,24]。胡超[24]曾认为胡蜂可以协助土沉香扩散到500 m远。对同属马来沉香(.)的研究表明,种子唯一传播者黄腰胡蜂()可以最远扩散到500 m以外[25]。除了胡蜂,是否还有其他动物或鸟类协助土沉香种子扩散还需进一步研究。土沉香种子的传播距离应在100 m以内,野外观察到土沉香母树的种子多是直接落在其附近,并在母树旁发芽,表明土沉香种子的扩散以重力居多。土沉香花粉和种子虽然有长距离传播的可能或情形,但多数情况下扩散有限,易于形成一定的空间遗传结构,即遗传相似性的个体呈聚集分布。
同时,利用不同分子遗传标记进行的土沉香不同种群遗传分化研究表明,不同地区土沉香种群间遗传分化较大[26–27]。黄久香等[26]采用ISSR (inter-simple sequence repeat)分子遗传标记对广东东莞、电白和广西玉林的土沉香种群遗传多样性进行了研究,结果表明这些种群间遗传分化为0.227;贾文杰等[1]采用ISSR分子标记对广东、广西、云南和海南地区8个种群的遗传多样性进行了研究,结果表明土沉香种群间的遗传分化达0.443。当遗传分化为0.15~0.25时,种群间的遗传分化程度较大; 当遗传分化大于0.25时,种群间的遗传分化程度很大[27]。因此,土沉香种群间出现了较大的遗传分化,说明土沉香花粉或种子扩散能力有限。
本研究结果表明,五桂山土沉香种群缺乏空间遗传结构,并呈现一些遗传差异较大的个体混杂的情况。在种群发育过程中,种群不同年龄级个体表现出不同的遗传结构[28–34]。祁彩虹等[28]对浙江省天台上甜槠()种群遗传结构进行了研究,结果表明有3个低年龄级都在小距离尺度上表现出较强的空间遗传结构,而大年龄级个体在不同空间距离尺度上均没有表现出空间遗传结构,并认为这是甜槠种子扩散有限造成,即有限的种子扩散距离导致遗传相似度大的低龄个体聚集在母树周围,引起空间遗传结构的发生;而在幼苗或小树长大后,个体间的竞争导致自疏作用发生,从而降低了大年龄级个体的空间遗传结构。Berens等[29]在肯尼亚对非洲李()的研究中也有类似的结果。但受到种子远距离扩散能力强、人类干扰等因素的影响,这种随年龄级而空间遗传结构逐渐变小的趋势并不一定会出现[32–34]。五桂山土沉香2个年龄级同样缺乏遗传结构,由于我们采集的土沉香为胸径大于5 cm的成年个体,可能是种群个体生长过程中自疏作用导致缺乏空间遗传结构,即围绕母树具有较高遗传相似度的个体(如全同胞或半同胞)在成长为大树过程中存在相互竞争,致使具有遗传差异较大的个体能相互共存。
同时,由于缺乏历史资料,虽然现有五桂山土沉香种群散布在次生林中,且有些个体胸径较大(58.2 cm),但种群整体起源状况并不明晰。经过咨询当地林业管理部门、农户,对于五桂山土沉香来源也没有相关资料凭证,大致认为这一地区土沉香资源很丰富,大树又很多,分布也很分散,一般不会是人工种植的。但考虑到中山市及其五桂山地区历史上是土沉香主产区,人工种植广泛,而且建国以来也开展过一些国土绿化工程,土沉香作为优良树种也会被用于进行林地改造等。因此,如果存在不同种源的人为种植,也会导致五桂山地区土沉香种群空间遗传结构的缺乏。
在空间遗传结构研究中,不同的空间采样方式会对结果有一定的影响[35–37]。采样方式大致包括了全部取样法、随机取样法(包括整体随机取样法、分组后组内随机法)、均匀取样法(按照网格等距离取样)、线性取样以及混合取样(包括上述各种取样)。通常,由于物种种群个体数量较大,能全部采样的状况非常少。而对于只能部分采样的状况,Schwartz等[36]的研究表明,除非物种种群个体遗传变异在空间上完全随机分布,否则没有任何一种部分取样方式的研究结果上没有偏差。Zeng等[37]在青藏高原对垫状点地梅()空间遗传结构的研究表明,对于整体随机取样法、分组后组内随机法以及线性取样3种方法,线性取样的研究结果略好,一是误差小,二是采样方式开支较少。
本研究的目的在于反映整个五桂山土沉香的空间遗传结构状况,因此采用沿山林小路全部取样的方式,保证了每个有土沉香的分布点均有样品采集。我们的采样类似于线性取样(图1),与随机取样相比,保证取样密度较大,可以反映更细致的空间遗传结构状况。而对于均匀取样,需要清楚掌握分布区内种群个体分布状况后才能很好设计网格进行采样,均匀取样虽然保证了样品分布的均匀性,但也会牺牲一定的细致度,取样密度低于全部取样。目前对五桂山土沉香的分布了解还局限在林道周围,对于群落中是否包含更多的土沉香资源有待进一步踏查和研究。
[1] JIA W J, LI E X, YANG B Y, et al. Studies on genetic diversity of[J]. J Trop Subtrop Bot, 2010, 18(2): 159–164. doi: 10.3969/j.issn.1005-3395.2010.02.008.贾文杰, 李恩香, 杨柏云, 等. 白木香遗传多样性研究 [J]. 热带亚热带植物学报, 2010, 18(2): 159–164. doi: 10.3969/j.issn.1005-3395. 2010.02.008.
[2] MEI Q X, LI H C, WANG K Y. Certificate of origin and development conception ofin Zhongshan [J]. Lishizhen Med Mat Med Res, 2007, 18(8): 2049–2051. doi: 10.3969/j.issn.1008-0805.2007. 08.166.梅全喜, 李汉超, 汪科元. 南药中山沉香的产地考证与发展构想[J]. 时珍国医国药, 2007, 18(8): 2049–2051. doi: 10.3969/j.issn.1008- 805.2007.08.166.
[3] MEI Q X, LI H C, WANG K Y, et al. Medical history and origin certificate of[J]. Pharm Today, 2011, 21(1): 3–5.梅全喜, 李汉超, 汪科元, 等. 南药沉香的药用历史与产地考证 [J]. 今日药学, 2011, 21(1): 3–5.
[4] MEI Q X. Fragrant Medicines:[M]. Beijing: China Press of Traditional Chinese Medicine, 2016: 1–240.梅全喜. 香药: 沉香[M]. 北京: 中国中医药出版社, 2016: 1–240.
[5] Wang Z F, Lian J Y, Huang G M, et al. Genetic groups in the common plant speciesand their associations with topographic habitats [J]. Oikos, 2012, 121(12): 2044–2051. doi: 10.1111/j.1600-0706.2012.20483.x.
[6] Wang Z F, Lian J y, Ye W H, et al. The spatial genetic pattern ofin a large forest plot with complex topography [J]. For Ecol Manage, 2014, 318: 318–325. doi: 10.1016/j.foreco.2014. 01.042.
[7] Wang Z F, Lian J y, Ye W h, et al. Pollen and seed flow under different predominant winds in wind-pollinated and wind-dispersed species[J]. Tree Genet Genomes, 2016, 12 (2): 19. doi: 10.1007/s11295-016-0973-3.
[8] JIAN Q C, TAN Z J, LIAO H B, et al. The genetic diversity ofin Wuguishan, Zhongshan, Guangdong Province [J]. Guihaia, 2018, 38(6): 804–811. doi: 10.11931/guihaia.gxzw201709027.蒋谦才, 谭宗健, 廖浩斌, 等. 广东省中山市五桂山土沉香()遗传多样性 [J]. 广西植物, 2018, 38(6): 804–811. doi: 10.11931/guihaia.gxzw201709027.
[9] PEAKALL R, SMOUSE P E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research: An update [J]. Bioinformatics, 2012, 28(19): 2537–2539. doi: 10.1093/bioinformatics/ bts460.
[10] ROUSSET F. GENEPOP'007: A complete re-implementation of the GENEPOP software for Windows and Linux [J]. Mol Ecol Resour, 2008, 8(1): 103–106. doi: 10.1111/j.1471-8286.2007.01931.x.
[11] WEIR B, COCKERHAM C. Estimating F-statistics for the analysis of population structure [J]. Evolution, 1984, 38(6): 1358–1370. doi: 10. 2307/2408641.
[12] Holm S. A simple sequentially rejective multiple test procedure [J]. Scand J Statist, 1979, 6(2): 65–70.
[13] Miller M P. Alleles In Space (AIS): Computer software for the joint analysis of interindividual spatial and genetic information [J]. J Hered, 2005, 96(6): 722–724. doi: 10.1093/jhered/esi119.
[14] Jombart T, Devillard S, Dufour A B, et al. Revealing cryptic spatial patterns in genetic variability by a new multivariate method [J]. Heredity, 2008, 101(1): 92–103. doi: 10.1038/hdy.2008.34.
[15] COLE C T. Genetic variation in rare and common plants [J]. Annu Rev Ecol Evol Syst, 2003, 34: 213–237. doi: 10.1146/annurev.ecolsys.34. 030102.151717.
[16] NYBOM H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants [J]. Mol Ecol, 2004, 13(5): 1143–1155. doi: 10.1111/j.1365-294X.2004.02141.x.
[17] Chen G, Liu C Q, Sun W B. Pollination and seed dispersal of(Lour.) Gilg (Thymelaeaceae): An economic plant species with extremely small populations in China [J]. Plant Divers, 2016, 38(5): 227–232. doi: 10.1016/j.pld.2016.09.006.
[18] Ward M, Dick C W, Gribel R, et al. To self, or not to self: A review of outcrossing and pollen-mediated gene flow in neotropical trees [J]. Heredity, 2005, 95(4): 246–254. doi: 10.1038/sj.hdy.6800712.
[19] Ismail S A, Ghazoul J, Ravikanth G, et al. Does long-distance pollen dispersal preclude inbreeding in tropical trees? Fragmentation genetics ofin an agro-forest landscape [J]. Mol Ecol, 2012, 21(22): 5484–5496. doi: 10.1111/mec.12054.
[20] Finger A, Kaiser-Bunbury C N, Kettle C J, et al. Genetic connectivity of the moth pollinated treein a highly fragmented habitat [J]. PLoS One, 2014, 9(10): e111111. doi: 10.1371/ journal.pone.0111111.
[21] Noreen A M E, Niissalo M A, Lum S K Y, et al. Persistence of long-distance, insect-mediated pollen movement for a tropical canopy tree species in remnant forest patches in an urban landscape [J]. Heredity, 2016, 117(6): 472–480. doi: 10.1038/hdy.2016.64.
[22] Hanson T R, Brunsfeld S J, Finegan B, et al. Pollen dispersal and genetic structure of the tropical treein a fragmented Costa Rican landscape [J]. Mol Ecol, 2008, 17(8): 2060– 2073. doi: 10.1111/j.1365-294X.2008.03726.x.
[23] Chávez-Pesqueira M, Suárez-Montes P, Castillo G, et al. Habitat fragmentation threatens wild populations of(Caricaceae) in a lowland rainforest [J]. Amer J Bot, 2014, 101(7): 1092–1101. doi: 10.3732/ajb.1400051.
[24] HU C. Seed dispersal inby paper wasp [D]. Nan- chang: Nanchang University, 2012: 1–57.胡超. 胡蜂对白木香种子传播的作用 [D]. 南昌: 南昌大学, 2012: 1–57.
[25] MANOHARA T N. Wasp-mediated seed dispersal in agarwood plant (), a critically endangered and overexploited species of north east India [J]. Curr Sci, 2013, 105(3): 298–299.
[26] HUANG J X, LIU X K, YE Y C, et al. Leaf morphology and genetic diversity ofin Dongguan and its surrounding areas [J]. Guangdong Agric Sci, 2014, 41(3): 153–158. doi: 10.3969/j.issn. 1004-874X.2014.03.037.黄久香, 刘宪宽, 叶永昌, 等. 东莞与邻近地区土沉香居群的叶形态和遗传多样性分析 [J]. 广东农业科学, 2014, 41(3): 153–158. doi: 10.3969/j.issn.1004-874X.2014.03.037.
[27] WRIGHT S. Evolution and the Genetics of Populations [M]. Chicago: University of Chicago Press, 1978: 1–590.
[28] QI C H, JIN Z X, LI J M. Small-scale spatial patterns of genetic structure inpopulations based on autocorrelation analysis in the Tiantai Mountain of Zhejiang Province [J]. Acta Ecol Sin, 2011, 31(18): 5130–5137.祁彩虹, 金则新, 李钧敏. 浙江天台山甜槠种群遗传结构的空间自相关分析 [J]. 生态学报, 2011, 31(18): 5130–5137.
[29] Berens D G, Braun C, GonzÁlez-MartÍnez S C, et al. Fine-scale spatial genetic dynamics over the life cycle of the tropical tree[J]. Heredity, 2014, 113(5): 401–407. doi: 10. 1038/hdy.2014.40.
[30] Chung M Y, Epperson B K, Chung M G. Genetic structure of age classes in(Theaceae) [J]. Evolution, 2003, 57 (1): 62–73. doi: 10.1111/j.0014-3820.2003.tb00216.x.
[31] Murrell D J. On the emergent spatial structure of size-structured populations: When does self-thinning lead to a reduction in clustering? [J]. J Ecol, 2009, 97(2): 256–266. doi: 10.1111/j.1365-2745.2008.01475.x.
[32] Roberts D G, Ottewell K M, Whelan R J, et al. Is the post- disturbance composition of a plant population determined by selection for outcrossed seedlings or by the composition of the seedbank? [J]. Heredity, 2014, 112(4): 409–414. doi: 10.1038/hdy.2013.119.
[33] González-Díaz P, Jump A S, Perry A, et al. Ecology and management history drive spatial genetic structure in Scots pine [J]. For Ecol Manage, 2017, 400: 68–76. doi: 10.1016/j.foreco.2017.05.035.
[34] Guidugli M C, Nazareno A G, Feres J M, et al. Small but not isolated: A population genetic survey of the tropical tree(Lecythidaceae) in a highly fragmented habitat [J]. Heredity, 2016, 116(3): 339–347. doi: 10.1038/hdy.2015.108.
[35] VEKEMANS X, HARDY O J. New insights from fine-scale spatial genetic structure analyses in plant populations [J]. Mol Ecol, 2004, 13 (4): 921–935. doi: 10.1046/j.1365-294X.2004.02076.x.
[36] Schwartz M K, McKelvey K S. Why sampling scheme matters: the effect of sampling scheme on landscape genetic results [J]. Conserv Genet, 2009, 10(2): 441–452. doi: 10.1007/s10592-008-9622-1.
[37] ZENG L Y, XU L L, TANG S Q, et al. Effect of sampling strategy on estimation of fine-scale spatial genetic structure in(Primulaceae), an alpine plant endemic to Qinghai-Tibetan Plateau [J]. J Syst Evol, 2010, 48(4): 257–264. doi: 10.1111/j.1759-6831.2010. 00084.x.
Spatial Genetic Structure ofin Wuguishan, Zhongshan, Guangdong Province
LIU Pan-pan1, LIAO Hao-bin1, JIANG Qian-cai1, TAN Zong-jian1, SUN Hong-mei1, WANG Rui-jiang2, CAO Hong-lin2, HUANG Xiang-xu2, Lai Si-ru2, Liang Dan2, WANG Zheng-feng2*
(1. Zhongshan State-owned Forest Resources Protected Center, Zhongshan 528407, Guangdong, China; 2. Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China)
In order to understand the spatial genetic structure ofin Wuguishan, Zhongshan City, Guangdong Province, the genotypes of 143 individuals of.with DBH (diameter at breast height) more than 5 cm were determined by using 18 microsatellite loci. The results showed that the observed heterozygosity and expected heterozygosity of.populationwere all 0.534, and inbreeding coefficient was 0.000 1, which indicated that the genetic diversity of.in Wuguishan was not low, and the whole population was under random mating situation. The genetic landscape shapes, spatial principal components analysis (sPCA) and spatial autocorrelation analysis showed.in Wuguishan lack spatial genetic structure. The individuals with different genetic backgrounds mixed, so that the neighbors had high genetic differences. So, it was suggested that the self thinning and artificial planting of.could be the two possible reasons.
; Microsatellite loci; Genetic diversity; Genetic landscape shape; Spatial principle component analysis; Spatial autocorrelation analysis
10.11926/jtsb.3917
2018-03-30
2018-06-01
中山市财政局资金项目; 东莞市科技项目(2015108101002); 广东省科技计划项目(2017A030303060); 广东省林业厅中药材保护项目; 广东省林业厅野生动植物保护和湿地保护专项资金(2130299); 广东省野生动植物保护管理项目资助
This work was supported by the Project in Zhongshan Finance Bureau, the Project for Science and Technology in Dongguan City (Grant No. 2015108101002), the Project for Science and Technology in Guangdong Province (Grant No. 2017A030303060), the Project for Conservation of Traditional Chinese Medicine in Forestry Administration of Guangdong, the Special Project for Conservation of Wild Animals, Plants and Wetlands in Forestry Administration of Guangdong (Grant No. 2130299), and the Project for Conservation and Management of Wild Animals and Plants in Guangdong Province.
刘盼盼(1986~ ), 女, 硕士, 林业工程师, 主要研究方向林业保护。E-mail: 769936661@qq.com
E-mail: wzf@scib.ac.cn
猜你喜欢
今日农业(2022年15期)2022-09-20
种子(2021年3期)2021-04-12
汽车维修与保养(2021年12期)2021-03-08
汽车维修与保养(2021年12期)2021-03-08
汽车维修与保养(2020年10期)2021-01-22
汽车维修与保养(2020年4期)2020-07-18
生物学教学(2018年3期)2018-08-08
中学生物学(2018年8期)2018-03-01
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
