
澳门九澳山海滨群落10种植物盛花期物候对极端气候事件的响应
2019-02-13 06:21张健豪付琳洪宝莹欧远雄邢福武易绮斐
热带亚热带植物学报 2019年1期
张健豪, 付琳, 洪宝莹, 欧远雄, 邢福武, 易绮斐*
澳门九澳山海滨群落10种植物盛花期物候对极端气候事件的响应
张健豪1,2, 付琳1, 洪宝莹3, 欧远雄3, 邢福武1, 易绮斐1*
(1. 中国科学院华南植物园,广州 510650;2. 中国科学院大学, 北京 100049;3. 澳门民政总署园林绿化部,澳门 999078)
为探究极端气候事件对植物的影响,对澳门九澳山海滨群落10种植物2012-2017年盛花期物候进行了观察。结果表明,植物的盛花期一般在3-9月,其中有4种植物为5月。2013和2016年早春澳门的极端强降水使植物的盛花期出现了明显的提前或者推迟。9种植物的盛花期与盛花期前0~2个月和上一年的秋冬季的月均温度或月降水存在显著相关性。温度和降水对植物盛花期的影响差异不大,但晚花植物对降水更加敏感。这为澳门和邻近岛屿的生态恢复和园林树种选材提供参考。
物候;盛花期;极端气候;澳门
物候学是研究植物或动物生命现象年周期变化与气候相互关系的学科[1]。在全球气候变暖的背景下,物候研究受到越来越多的关注,物候观察一直是研究气候变化对生物影响的重要方法[2]。植物内部调控和外部环境共同决定着其物候期。很多气候因素对植物物候有显著影响,在英国[3]、德国[4]和中国北方[5]等温带地区的研究表明,温度[6–7]和光周期[8–9]是改变植物物候最直接的非生物刺激。与此同时,在热带、亚热带地区[10–12]、干旱地带[13–14]和地中海地区[15–16]的研究表明,降水是影响植物物候的关键因素。
在南亚热带的森林中,许多物种几乎是全年萌芽,叶变色和落叶现象不会像温带的植物那样明显,而开花是最易被观察到的物候现象。对个体而言,开花是植物繁殖过程中的重要阶段,影响后续的果实成熟和种子散布等的时间,同时影响了参与这些环节的动物行为[17–18];对种群而言,不同物种间开花时间的差异影响着它们对资源的相互竞争,甚至是生态位的分化,进而关系到整个种群的生存和延续[19–20]。许多研究表明,在过去的几十年中,随着全球范围内气候变暖,世界各地植物的春季物候有明显的提前[4,21];而另一些物种的春季物候变化非常微小甚至出现了推迟[3,22]。另一方面,具有不同功能性状的物种对气候变化的敏感程度也不同,早花植物对气候变化的响应比晚花植物更敏感[22–24];不同传粉类型的物种适应气候变化的机制也是多样的[25–27];甚至物候的发生和物种间的谱系关系也有一定的相关性[19,28]。
目前,多数研究着眼于长期的、缓慢的气候变暖趋势下植物物候的变化,而关于极端气候事件对植物物候的影响研究较少[29],如短期的强降水、热浪和极端干旱等。Both等[30]认为短期的极端气候事件相比于长期缓慢的气候变化,对干扰生物体的生命活动和物候同步性有着更强的作用。由北大西洋涛动(NAO)引发的气候骤变与德国[4]、拉脱维亚和立陶宛[23]的一些物种开花时间有着密切联系;在美国卡萨斯州,长时间干旱后的突然降水会使植物开花的持续时间缩短[31];类似的,开花前突发的降水也会影响到美国安息香科植物的开花时间和数量[32]。各种类型的极端气候事件的发生时间和间隔周期是很难预测的,但它们对生态环境产生的影响不容忽视[33]。
本研究是澳门野生植物物候监测项目的一部分,该项目始于2012年1月,目前仍在持续进行中。2013年春季,中国华南地区经历了持续几周的暴雨袭击[34],期间澳门的降水量也异常偏高, 3至6月总降水量为1 274.4 mm,比第二高的年份2016年多了274 mm,比1981至2010年同期平均水平多315 mm。另外受2015年的超强厄尔尼诺事件影响,2016年上半年中国南方降水偏多,澳门在2016年1月的降水量达278.2 mm,是2015年同期(27.6 mm)的10.1倍。2013年,我们观察到九澳山海滨群落中的许多种植物,如假苹婆()、豺皮樟()等的开花时间比其他年份早,这可能与2013年春季的异常降水有关。本文利用澳门九澳山海滨群落6年来的植物开花物候数据,初步探究植物盛花期它们对异常气候事件的响应,了解九澳山海滨群落的植物开花时间与降水和温度的变化是否相关,极端的降水对一些植物盛花期的影响如何,可填补澳门地区物候研究的空白,为澳门和邻近岛屿的生态恢复研究和园林树种选材提供参考数据。
1 材料和方法
1.1 研究区概况
澳门特别行政区位于珠江口西南岸,东与香港隔海相望,北与广东省珠海市接壤,由澳门半岛、氹仔岛和路环岛三部分组成。根据《2016澳门年鉴》和地球物理暨澳门气象局(www.smg.gov.mo)的数据,2015年澳门特别行政区总面积为30.4 km2。该区域属于南亚热带,为海洋性季风气候。2012-2017年,澳门的夏季(>22℃)从4月开始持续7个月,除2012年外,最冷月平均气温均高于15℃,表明了澳门四季温暖,冬短夏长的气候特点。这6年中, 澳门从11月到翌年2月的降水较少,春夏季的降水丰沛但年际间变化很大。九澳山位于澳门路环东南海滨,植被类型为亚热带季风常绿阔叶林。
1.2 方法
本研究在九澳山海滨自然形成的森林群落中设置400 m2的样地,平均划分为16块5 m×5 m的小样方,以字母A~P排列。对样方内所有胸径≥1 cm且生长正常的乔木,以及灌木和草本层中最常见的物种进行挂牌标号。连续6年进行定点定株观察(春夏季每月2次,秋冬季每月1次)。参照《中国物候观测方法》[35]进行物候期判断和记录。
本研究采用儒略日(julian day)的统计方法,将植物物候记录的日期转化为距当年1月1日的天数。将第一次观察到植株有开花现象的儒略日作为始花期,将第一次观察到植株落花达到95%的儒略日作为开花结束期,某个个体的始花期和开花结束期的中点作为盛花期,某物种所有观察个体当年的盛花期平均值为其当年的盛花期,6年盛花期平均值为该物种平均盛花期。由于样本大小和观测频率不同对植物始花期确定的准确性影响较大,因此很多研究均采取盛花期数据来减小这种误差[22,36–38]。本研究记录了从2012年1月到2018年2月共74个月的物候观测数据(有些晚花植物开花会持续到第二年,因此2018年头两个月的数据也包括在内)。在这6年中,有少量个体消失或死亡,也有一些幼龄植株成熟后挂牌;另外由于部分挂牌个体不是每年都能观察到开花,因此每年观察到开花的植株可能不同。经统计,样地中每年都有开花记录并且每年开花个体数不少于3株的共有10种植物(表1)。本研究的气象数据来源于澳门地球物理暨气象局网站(www.smg.gov.mo)。
为了研究这6年间异常气候对植物物候的影响,将每年各月份的均温和降水量数据进行比较,采用SPSS 24.0软件中的箱形图进行异常值检验。花期和气候因子间的关系通常可以作为对不同时期的敏感性衡量指标。很多植物的花期都会和开花前某一段时期的气候因子有关,我们采用Pearson的相关系数来检验各物种6年盛花期的变化和从上年9月到盛花期当月的月均温度和月降水量相关系数的显著性。出现显著相关的月份被定义为最优时期。
2 结果和分析
10种植物的花期多集中在春季和夏季(图1), 6年平均盛花期为3月25日到11月26日。平均盛花期出现在5月的物种数最多,有台湾相思(Acacia confusa)、假苹婆、九节(Psychotria asiatica)和广东蒲桃(Syzygium kwangtungense) 4种。不同物种的盛花期年际标准差也存在很大差异,其中最大的是白楸(),达30.4 d,说明6年来白楸盛花期出现的时间最分散,年际差异大;栀子()的最小,只有4.9 d,说明其盛花期相对集中,年际波动小(表1和图1)。

表1 九澳山海滨群落10种植物6年中盛花期变化情况
在2012-2017年期间,有5种植物的盛花期在2013年出现得最早或最晚,有3种植物是在2016年。从图2和3可见,气候极端异常现象有2013年2月降水量(偏低)、2013年4月的均温(偏低)、2015年11月均温(偏高)和2016年1月降水量(偏高)。6年中异常气候主要集中在2013年春夏季和2015年冬季到2016年春季两个时段。对植物2013和2016年盛花期分别与其余年份盛花期进行单因素方差分析,结果表明2013年豺皮樟、假苹婆、九节和鸭脚木()的差异极显著(<0.01),而栀子的差异不显著。2016年白楸和豺皮樟的差异极显著(<0.01),而两面针()的差异不显著。
最早盛花期或最晚盛花期发生在2013年的植物分别有2和3种,其中鸭脚木的盛花期提前最多,达28 d,栀子只提前7 d;假苹婆推迟得最多,达39 d,其次为豺皮樟和九节,分别推迟12.9和7.7 d。在2016年,有3种植物出现了最早盛花期。白楸、豺皮樟和两面针的盛花期分别提前53.7、13.6和3.7 d。
九澳山海滨群落中有9种植物的盛花期与上年9月到盛花期当月的月均温或月降水量存在显著相关性,且这些月份几乎全部集中在2个时期:上年秋冬季(第一阶段)和自身盛花期当月或之前的0~2个月(第二阶段)。
从表2和3可以看出,在这6年中,栀子的盛花期无论是与温度和降水量的相关性,还是对异常气候的响应都不显著,表明栀子可能对降水量和温度变化相对不敏感,影响其开花的机制有待进一步探究,可能是由更复杂的内部机制调控或受到其他的环境因子如光照等的影响。两面针和台湾相思的盛花期与上年秋冬季的温度或降水呈显著正相关,即此时较高的温度或较多的降水使花期推迟,但他们对极端天气都没有明显的响应,这是由于两次极端气候事件都出现在春季。因此不论是从对极端天气的响应角度还是与气象因子的相关性角度,都反映出春季的温度与降水变化对两面针和台湾相思的盛花期影响较小。广东蒲桃和梅叶冬青的盛花期与第二阶段的均温或降水呈显著正相关, 即此时较高的温度或较多的降水使花期推迟,但他们对极端天气都没有明显的响应,可能由于异常气候出现的时期对他们盛花期影响较小。九节的盛花期与2月的降水量呈显著负相关,2013年2月的降水量异常偏低,九节的盛花期出现了显著推迟。假苹婆的盛花期与4月的温度呈显著负相关,2013年4月气温异常偏低,假苹婆盛花期出现了显著推迟。鸭脚木的盛花期与9月的降水呈显著负相关,2013年9月降水偏高,鸭脚木的盛花期提前了28 d。白楸的盛花期为6到8月,其与6和8月的降水都成显著负相关,说明春季和盛花期前0~3个月的降水可以有效促进白楸开花,而2016年春季的降水异常偏多,白楸的盛花期相应地提前了53.7 d。豺皮樟的盛花期与上年11月的均温呈显著负相关,而2015年11月的月均温异常偏高,因此豺皮樟2016年的盛花期提前13.6 d;另一方面,豺皮樟与2至4月的降水均呈一定的负相关,但2013年2月降水异常偏低,3-6月的降水又骤然增多[34],豺皮樟出现了最晚盛花期,说明其对2月的降水量更敏感, 异常少的降水使豺皮樟的开花受到较大限制。
我们将平均盛花期在6月1日之前的作为早花植物,有两面针、栀子、台湾相思、假苹婆、九节和广东蒲桃,其余的为晚花植物。晚花植物的盛花期年际标准差与早花植物有显著差异(=0.014),说明晚花植物盛花期的年际变化要比早花植物更大,早花植物的盛花期相对更稳定。从图2和3可见,月降水的年际变化明显大于月均温度的,温度相对于降水是较稳定的气象因子。在盛花期与降水呈显著相关的5种植物中有4种是晚花植物,因此晚花植物可能对降水更敏感,相应地,晚花植物也有更大的年际变化。
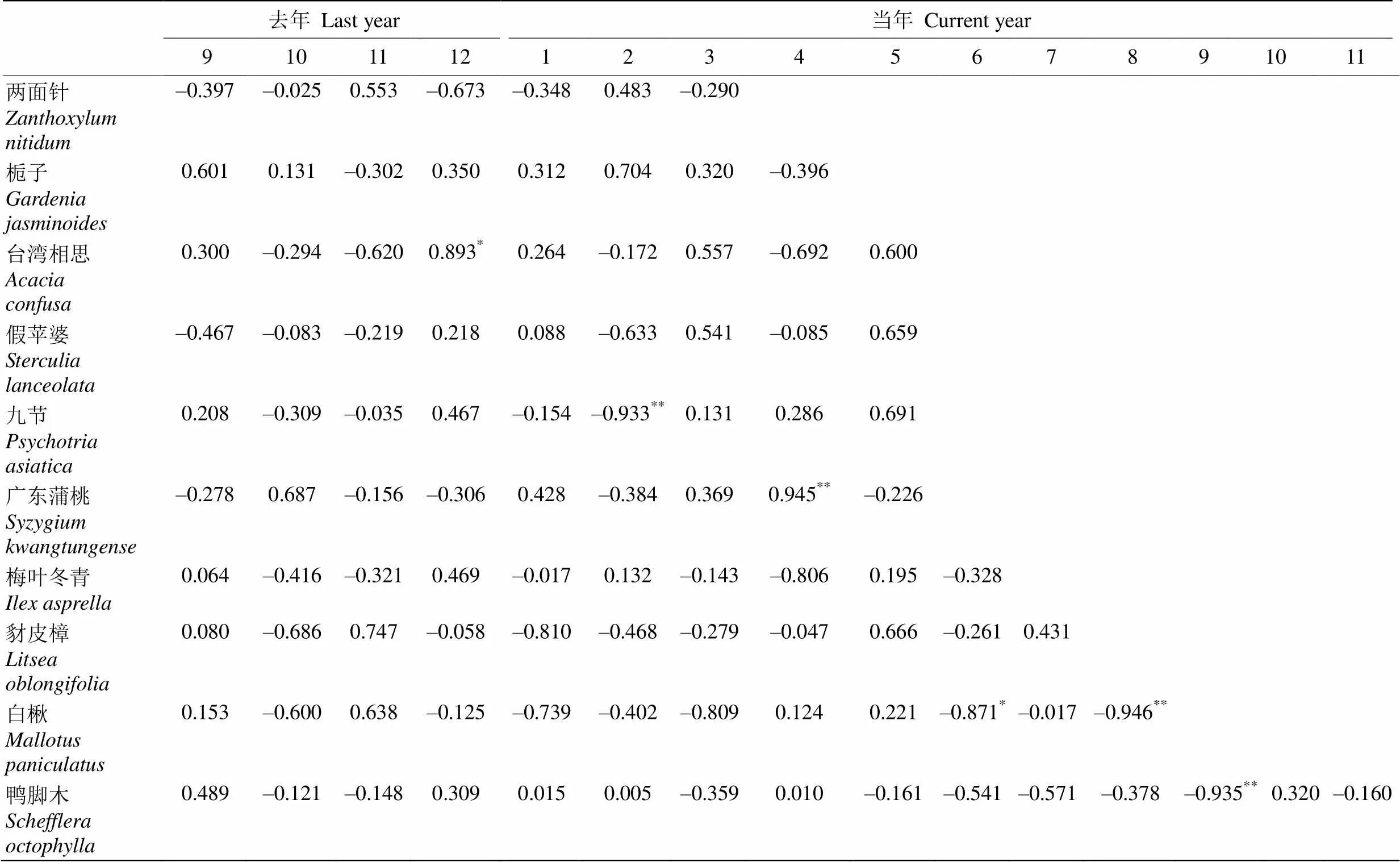
表2 盛花期和月均降水量的相关性
*:<0.05; **:<0.01

表3 盛花期和月均温度的相关性
*:<0.05; **:<0.01
3 结论和讨论
本研究结果表明,梅叶冬青、假苹婆、白楸、广东蒲桃和鸭脚木5种植物对盛花期前0~2个月的降水或者温度有显著响应,这与前人研究的结论一致[3–4,6,39],即盛花期前0~2个月的水热条件直接影响了这些植物的生理活动,并决定了开花的时机。盛花期前充足的降水能促进白楸和鸭脚木开花,较高的温度促进假苹婆的开花。然而梅叶冬青的盛花期与当月均温呈显著正相关,这可能是高温能延长一些植物的花期,从而使盛花期推迟。另外,上一年秋冬季的温度和降水也与一些植物的盛花期有显著相关性。上年秋季和冬季的气候强烈地影响了植物的变色和落叶等衰老物候,以及第二年的萌芽展叶物候,这与开花有很强的时序关系,特别是对 于热带落叶物种[40]。Lambert等报道,前一年夏季的大量降水导致了洛基山百合科植物开花提前[41]。此外,对海南[42]、贵阳[43]等地植物的研究也得出了类似的结论,认为气候因子对物候的影响具有滞后性。在温带地区温度是影响植物开花物候最主要的气象因素[6–7,44]。在热带或亚热带地区,如一些亚洲热带森林[45], 降水的变化幅度大于温度,降雨成为诱发开花的重要因素,然而也有一些研究表明,植物的始花期对温度的敏感性在亚热带地区比高纬地区更强[46]。由于物种数量有限,本研究中,月均温和月降水量这两个气象因子对这10种植物的影响没有明显差异。
早花植物始花期的年际变化比晚花植物要 大[3,24,47]。Korner[7]的研究表明,相对于光周期的变化,在一些非热带地区温度的变化幅度会更大,因此植物随温度变化而产生的生理活动是一种机会更大但更加冒险的生存策略。然而,在全球气候变暖的背景下,一些演化早期的物种和生命周期短的物种,为了取得对光敏感物种的竞争优势,会选择这一种风险更大,但是机会更多的策略。与此类似,Cleland等[44]和Willis等[45]的研究表明,对气候变化更敏感的物种在群落中的个体数和花的数量会增加,而相对不敏感的物种在群落中的多度会降低。有些外来种的始花期对温度敏感性更高,从而在群落中占据入侵优势。本研究中,晚花植物是不是也采取了一种类似的对降水更敏感的策略呢?目前本研究的物候观察只开展了6年,且仅筛选出了10种有较完整物候记录的植物,受此限制,仅对月均温和月降水量两种气候因子对盛花期的影响进行了研究。然而植物物候对气候变化的响应是一个协同变化不断耦合的过程[46],其他的物候因子如湿度和光照等和温度与降水之间有怎样的相互作用, 它们对盛花期的影响程度分别有多大?植物盛花期和其它物候期特别是萌芽期等又有怎样的关系?许格希等[42]和裴顺祥等[47]利用多年的物候数据,建立了积分回归-物候预测模型,定量分析了植物物候和气候变化的复杂动态关系过程。随着本研究的继续开展,积累更多的物候资料后,将会对澳门松山植物对气候变化的响应规律做出更深入全面的探讨。
澳门地处中国大陆南端沿海,时常遭遇极端气候事件,如台风、极端降水等。2017年8月下旬的台风“天鸽”使澳门遭受了几十年一遇的罕见风灾,造成惨重的人员伤亡和经济损失的同时,也使自然森林群落和市政绿化植物遭到严重破坏。研究极端气候事件对植物物候的影响以及其与群落格局变化的关系,能为后续的生态恢复和园林植物选材等提供参考。
综上所述,九澳山海滨群落的10种植物盛花期集中在春季和夏季,其中大多数植物的盛花期对前0~2个月的月降水量或月均温度有显著的响应,前一年秋冬季的平均气温和降水也是影响开花时间的因素。在2013或2016年,有7种植物的盛花期对异常气候有不同程度的响应,也与他们对温度和降水的相关性是一致的,月降水量和月均温度这两个气候因子对10种植物的物种盛花期的影响没有明显差异,且晚花植物的盛花期年际差异更大, 对降水更加敏感。
[1] McEWAN R W, McCARTHY B C. Reviewed work: Phenology: An integrative environmental science by Mark D. Schwartz [J]. J Torrey Bot Soc, 2005, 132(1): 170–171.
[2] SPARKS T H, JEFFREE E P, JEFFREE C E. An examination of the relationship between flowering times and temperature at the national scale using long-term phenological records from the UK [J]. Int J Biometeorol, 2000, 44(2): 82–87. doi: 10.1007/s004840000049.
[3] FITTER A H, FITTER R S R, HARRIS I T B, et al. Relationships between first flowering date and temperature in the flora of a locality in central England [J]. Funct Ecol, 1995, 9(1): 55–60. doi: 10.2307/239 0090.
[4] MENZEL A. Plant phenological anomalies in Germany and their relation to air temperature and NAO [J]. Clim Change, 2003, 57(3): 243–263. doi: 10.1023/A:1022880418362.
[5] WANG H J, ZHONG S Y, TAO Z X,et al. Changes in flowering phenology of woody plants from 1963 to 2014 in north China [J/OL]. Int J Biometeorol, 2017, [2017-05-25]. doi: 10.1007/s00484-017- 1377-2.
[6] MENZEL A, SPARKS T H, ESTRELLA N, et al. European phenological response to climate change matches the warming pattern [J]. Glob Change Biol, 2006, 12(10): 1969–1976. doi: 10.1111/j.1365-2486.2006. 01193.x.
[7] KÖRNER C, BASLER D. Phenology under global warming [J]. Science, 2010, 327(5972): 1461–1462. doi: 10.1126/science.1186473.
[8] JEONG S, CLARK S E. Photoperiod regulates flower meristem deve- lopment in[J]. Genetics, 2005, 169(2): 907–915. doi: 10.1534/genetics.104.033357.
[9] JACKSON S D. Plant Responses to Photoperiod [M]. Oxford, UK: Blackwell Publishing Ltd., 2009: 517,531.
[10] BREARLEY F Q, PROCTOR J, NAGY L,et al. Reproductive phenology over a 10-year period in a lowland evergreen rain forest of central Borneo [J]. J Ecol, 2007, 95(4): 828–839. doi: 10.1111/j.1365-2745. 2007.01258.x.
[11] CORLETT R T, LAFRANKIE J V Jr. Potential impacts of climate change on tropical Asian forests through an influence on phenology [J]. Climatic Change, 1998, 39(2/3): 439–453. doi: 10.1023/A:100532812 4567.
[12] REICH P B. Phenology of tropical forests: patterns, causes, and consequences [J]. Can J Bot, 1995, 73(2): 164–174. doi: 10.1139/b95- 020.
[13] McLAREN K P, McDONALD M A. Seasonal patterns of flowering and fruiting in a dry tropical forest in Jamaica [J]. Biotropica, 2005, 37(4): 584–590. doi: 10.1111/j.1744-7429.2005.00075.x.
[14] de LAMPE M G, BERGERON Y, MCNEIL R,et al. Seasonal flowering and fruiting patterns in tropical semi-arid vegetation of northeastern Venezuela [J]. Biotropica, 1992, 24(1): 64–76. doi: 10. 2307/2388474.
[15] PEÑUELAS J, FILELLA I, ZHANG X Y, et al. Complex spatio- temporal phenological shifts as a response to rainfall changes [J]. New Phytol, 2004, 161(3): 837–846. doi: 10.1111/j.1469-8137.2004.01003.x.
[16] PEÑUELAS J, FILELLA I, COMAS P. Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region [J]. Glob Change Biol, 2002, 8(6): 531–544. doi: 10.1046/j.1365-2486.2002.00489.x.
[17] STENSETH N C, MYSTERUD A. Climate, changing phenology, and other life history traits: Nonlinearity and match-mismatch to the environment [J]. Proc Natl Acad Sci USA, 2002, 99(21): 13379–13381. doi: 10.1073/pnas.212519399
[18] BOTH C, VISSER M E. Adjustment to climate change is constrained by arrival date in a long-distance migrant bird [J]. Nature, 2001, 411 (6835): 296–298. doi: 10.1038/35077063.
[19] DU Y J, MAO L F, QUEENBOROUGH S A,et al. Phylogenetic constraints and trait correlates of flowering phenology in the angio- sperm flora of China [J]. Glob Ecol Biogeogr, 2015, 24(8): 928–938. doi: 10.1111/geb.12303.
[20] CRAINE J M, WOLKOVICH E M, Towne E G, et al. Flowering phenology as a functional trait in a tallgrass prairie [J]. New Phytol, 2012, 193(3): 673–682. doi: 10.1111/j.1469-8137.2011.03953.x.
[21] ROOT T L, PRICE J T, HALL K R, et al. Fingerprints of global warming on wild animals and plants [J]. Nature, 2003, 421(6918): 57– 60. doi: 10.1038/nature01333.
[22] MILLER-RUSHING A J, INOUYE D W. Variation in the impact of climate change on flowering phenology and abundance: An examinationof two pairs of closely related wildflower species [J]. Amer J Bot, 2009, 96(10): 1821–1829. doi: 10.3732/ajb.0800411.
[23] KALVĀNE G, ROMANOVSKAJA D, BRIEDE A,et al. Influence of climate change on phenological phases inand[J]. Climate Res, 2009, 39: 209–219. doi: 10.3354/cr00813.
[24] McEWAN R W, BRECHA R J, GEIGER D R,et al. Flowering phenology change and climate warming in southwestern Ohio [J]. Plant Ecol, 2011, 212(1): 55–61. doi: 10.1007/s11258-010-9801-2.
[25] SPARKS T H, YATES T J. The effect of spring temperature on the appearance dates of British butterflies 1883-1993 [J]. Ecography, 1997, 20(4): 368–374. doi: 10.1111/j.1600-0587.1997.tb00381.x.
[26] BOLMGREN K, ERIKSSON O, LINDER H P. Contrasting flowering phenology and species richness in abiotically and biotically pollinated angiosperms [J]. Evolution, 2003, 57(9): 2001–2011. doi: 10.1111/j. 0014-3820.2003.tb00380.x.
[27] FITTER A H, FITTER R S R. Rapid changes in flowering time in British plants [J]. Science, 2002, 296(5573): 1689–1691. doi: 10.1126/ science.1071617.
[28] DAVIES T J, WOLKOVICH E M, KRAFT N J B,et al. Phylogenetic conservatism in plant phenology [J]. J Ecol, 2013, 101(6): 1520–1530. doi: 10.1111/1365-2745.12154.
[29] JENTSCH A, KREYLING J, BEIERKUHNLEIN C. A new generation of climate-change experiments: Events, not trends [J]. Front Ecol Environ, 2007, 5(7): 365–374. doi: 10.1890/1540-9295(2007)5[365: ANGOCE]2.0.CO;2.
[30] BOTH C, BOUWHUIS S, LESSELLS C M,et al. Climate change and population declines in a long-distance migratory bird [J]. Nature, 2006, 441(7089): 81–83. doi: 10.1038/nature04539.
[31] FAY P A, CARLISLE J D, KNAPP A Ket al. Altering rainfall timing and quantity in a mesic grassland ecosystem: Design and performance of rainfall manipulation shelters [J]. Ecosystems, 2000, 3(3): 308–319. doi: 10.1007/s100210000028.
[32] MAHALL B E, THWING L K, TYLER C M. A quantitative comparison of two extremes in chaparral shrub phenology [J]. Flora, 2010, 205(8): 513–526. doi: 10.1016/j.flora.2009.12.011.
[33] JENTSCH A, KREYLING J, BOETTCHER-TRESCHKOW J,et al. Beyond gradual warming: Extreme weather events alter flower phenology of European grassland and heath species [J]. Glob Change Biol, 2009, 15(4): 837–849. doi: 10.1111/j.1365-2486.2008.01690.x.
[34] HU Y M, ZHAI P M, LUO X L,et al. Large scale circulation and low frequency signal characteristics for the persistent extreme precipitation in the first rainy season over south China in 2013 [J]. Acta Meteor Sin, 2014, 72(3): 465–477. doi: 10.11676/qxxb2014.042.胡娅敏, 翟盘茂, 罗晓玲, 等. 2013年华南前汛期持续性强降水的大尺度环流与低频信号特征 [J]. 气象学报, 2014, 72(3): 465–477. doi: 10.11676/qxxb2014.042.
[35] WAN M W, LIU X Z. Method of Phenology Observation of China [M]. Beijing: Science Press, 1979: 1–22.宛敏渭, 刘秀珍. 中国物候观测方法[M]. 北京: 科学出版社, 1979: 1–22.
[36] BERTIN R I. Climate change and flowering phenology in Worcester County, Massachusetts [J]. Int J Plant Sci, 2015, 176(2): 107–119. doi: 10.1086/679619.
[37] MILLER-RUSHING A J, PRIMACK R B, PRIMACK D,et al. Photographs and herbarium specimens as tools to document phenological changes in response to global warming [J]. Amer J Bot, 2006, 93(11): 1667–1674. doi: 10.3732/ajb.93.11.1667.
[38] CARADONNA P J, ILER A M, INOUYE D W. Shifts in flowering phenology reshape a subalpine plant community [J]. Proc Natl Acad Sci USA, 2014, 111(13): 4916–4921. doi: 10.1073/pnas.1323073111.
[39] PRIMACK D, IMBRES C, PRIMACK R B, et al. Herbarium specimens demonstrate earlier flowering times in response to warming in Boston [J]. Amer J Bot, 2004, 91(8): 1260–1264. doi: 10.3732/ajb.91.8.1260.
[40] RIVERA G, ELLIOTT S, CALDAS L S, et al. Increasing day-length induces spring flushing of tropical dry forest trees in the absence of rain [J]. Trees, 2002, 16(7): 445–456. doi: 10.1007/s00468-002-0185-3.
[41] LAMBERT A M, MILLER-RUSHING A J, INOUYE D W. Changes in snowmelt date and summer precipitation affect the flowering phenology of(glacier lily; Liliaceae) [J]. Amer J Bot, 2010, 97(9): 1431–1437. doi: 10.3732/ajb.1000095.
[42] XU G X, LUO S X, GUO Q S, et al. Responses of leaf unfolding and flowering to climate change in 12 tropical evergreen broadleaf tree species in Jianfengling, Hainan Island [J]. Chin J Plant Ecol, 2014, 38(6): 585–598. doi: 10.3724/SP.J.1258.2014.00054.许格希, 罗水兴, 郭泉水, 等. 海南岛尖峰岭12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应 [J]. 植物生态学报, 2014, 38(6): 585–598. doi: 10.3724/SP.J.1258.2014.00054.
[43] BAI J, GE Q S, DAI J H. Response of woody plant phenophases to climate change for recent 30 years in Guiyang [J]. Geograph Res, 2009, 28(6): 1606–1614.白洁, 葛全胜, 戴君虎. 贵阳木本植物物候对气候变化的响应 [J]. 地理研究, 2009, 28(6): 1606–1614.
[44] CLELAND E E, ALLEN J M, CRIMMINS T M,et al. Phenological tracking enables positive species responses to climate change [J]. Ecology, 2012, 93(8): 1765–1771. doi: 10.1890/11-1912.1.
[45] WILLIS C G, RUHFEL B, PRIMACK R B,et al. Phylogenetic patterns of species loss in thoreau’s woods are driven by climate change [J]. Proc Natl Acad Sci USA, 2008, 105(44): 17029–17033. doi: 10.1073/ pnas.0806446105.
[46] LI M C, ZHU J J, SUN Y R. Uncertainty analysis in physio-ecological response of plant to climate change [J]. Acta Bot Boreali-Occid Sin, 2009, 29(1): 207–214. doi: 10.3321/j.issn:1000-4025.2009.01.034.李明财, 朱教君, 孙一荣. 植物对气候变化生理生态响应的不确定性分析 [J]. 西北植物学报, 2009, 29(1): 207–214. doi: 10.3321/j. issn:1000-4025.2009.01.034.
[47] PEI S X, GUO Q S, JIA Y B, et al. Integral regressive analysis on the responses of first flowering date of eight woody species to climate change in Baoding, China [J]. J Beijing For Univ, 2015, 37(7): 11–18. doi: 10.13332/j.1000-1522.20130423.裴顺祥, 郭泉水, 贾渝彬, 等. 保定市8种乔灌木开花始期对气候变化响应的积分回归分析 [J]. 北京林业大学学报, 2015, 37(7): 11– 18. doi: 10.13332/j.1000-1522.20130423.
Effects of Extreme Climate Events on Phenology at Flowering Period of 10 Species in Ka Ho Hill, Macao
ZHANG Jian-hao1,2, FU Lin1, HONG Pou-leng3, AO Un-hong3, XING Fu-wu1, YI Qi-fei1*
(1. South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China; 2. University of Chinese Academy Sciences,Beijing 100049, China; 3. Department of Garden and Green Area, Civil and Municipal Affairs Bureau of Macao Special Administrative Region, Macao 999078, China)
In order to understand the effect ofextreme climate events on plants, the phenology at flowering period of 10 species in Ka Ho Hill, Macao were observed from 2012 to 2017. The results showed that the flowering period of 10 species was usually from March to September, in which that of 4 species was in May. The flowering period of the plants has been significantly advanced or delayed by extreme heavy rainfall in early spring of 2013 and 2016. There were significant relations between flowering period of 9 species and monthly mean temperature or monthly precipitation in autumn and winter of last year and 0–2 months before flowering period. In addition, there was not distinct difference in flowering period of plants between temperature and precipitation, but late-flowering species were relatively sensitive to precipitation. Therefore, these would provide references for ecological restoration and selection of garden tree species in Macao and neighboring islands.
Phenology; Flowering period; Extreme climate; Macao
10.11926/jtsb.3921
2018-04-04
2018-06-05
澳门野生植物物候监测项目资助
This work was supported by the Project for Phenological Monitoring of Wild Plants in Macao.
张健豪(1992~ ), 男,硕士研究生,主要从事植物物候研究。E-mail: zhangjh@scbg.ac.cn
E-mail: yiqifei@scbg.ac.cn
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
农产品市场周刊(2022年4期)2022-05-21
农产品市场周刊(2022年4期)2022-03-23
成都信息工程大学学报(2021年4期)2021-11-22
——以长白山区为例
东北林业大学学报(2020年9期)2020-10-21
农业灾害研究(2020年8期)2020-05-11
疯狂英语·新读写(2018年3期)2018-11-29
山西果树(2015年3期)2015-05-29
