
一个水稻低温移栽白条纹突变体wltt的鉴定和基因定位
2019-01-24 03:52林添资孙立亭龚红兵王益华刘玲珑赵志刚江玲万建民
中国水稻科学 2019年1期
林添资 孙立亭 龚红兵 王益华 刘玲珑 赵志刚 江玲 万建民, *
一个水稻低温移栽白条纹突变体的鉴定和基因定位
林添资1, 2孙立亭2龚红兵2王益华1刘玲珑1赵志刚1江玲1万建民1, *
(1南京农业大学 作物遗传与种质创新国家重点实验室/农业部长江中下游粳稻生物学与遗传育种重点实验室/长江流域杂交水稻协同创新中心/江苏省现代作物生产协同创新中心, 南京 210095;2江苏丘陵地区镇江农业科学研究所, 江苏 句容 212400;*通讯联系人, E-mail: wanjm@njau.edu.cn)
【目的】叶色突变相关基因的鉴定与克隆为研究叶绿体发育、叶绿素合成和光合作用等分子机制提供理论基础。【方法】从常规粳稻镇糯19杂交后代中分离出一个低温移栽后叶色转成白条纹的自然变异突变体, 命名为。成熟期测定野生型和的主要农艺性状,分别在苗期、移栽后15 d和同时期直播条件下测定新生叶片的色素含量并观察叶绿体的超微结构;将和野生型正反交进行遗传分析;用与籼稻9311杂交产生的F2作为定位群体进行基因定位;采用RT-qPCR 分析叶绿体发育、叶绿素合成和光合作用相关基因在野生型和中的表达水平。【结果】突变体在苗期表现正常绿色,移栽15 d后心叶出现白条纹叶表型,至分蘖末期心叶叶色恢复;而不经移栽,突变体不会出现白条纹叶。人工模拟实验表明该表型是由低温条件下根损伤引起的。与野生型相比,突变体移栽后的新生叶色素含量显著降低,光合速率下降;同时株高变矮,穗长、剑叶长和每穗粒数均显著降低。叶绿体的超微结构显示,突变体的叶肉细胞中,仅少数细胞含有正常的叶绿体,其余大部分叶肉细胞不含叶绿体。进一步研究发现,突变体中部分光合系统相关基因和叶绿体发育相关基因表达下调,叶绿素生物合成相关的14个基因表达也下调。遗传分析表明,该突变性状受一对隐性核基因控制。利用突变体/9311的F2群体,将该基因定位于水稻第2染色体着丝粒附近853 kb区间内。目前,该区间内没有叶色相关基因的报道。【结论】是低温条件下移栽调控叶片转色的关键基因,在叶绿体发育过程中发挥重要作用。
水稻;移栽叶片转色;叶绿体发育;基因定位
叶绿体是植物进行光合作用的场所,也是植物体内很多有机大分子如淀粉、脂类和氨基酸的合成场所[1],因此叶绿体的正常发育对植物至关重要。叶绿体存在缺陷的突变体往往表现出肉眼可见的异常叶色表型,如叶色淡绿、黄化、白条纹、斑马叶等,并通常伴随有一系列生长发育缺陷,严重的会导致植株死亡[2-4]。叶色突变与植物光合作用[5]、激素生理[6]和光形态建成[7]等密切相关,对它们的研究有助于阐明光合作用机制、叶绿素生物合成及代谢途径以及叶绿体发育的调控机理,为开展水稻高光效育种提供理论基础。此外,水稻的叶色变异可用作标记性状剔除假杂种,提高种子纯度,降低制种风险[8]。因此,水稻叶色变异相关基因的克隆及功能分析具有重要的理论意义和生产应用价值。
迄今为止,关于叶色突变的研究很多。突变基因在水稻12条染色体上均有分布,大多数突变性状受单隐性核基因控制[9],而由细胞质基因和显性基因控制的叶色突变很少[10-11]。引起叶色变异的机理主要有叶绿素代谢途径受阻[12-13]和叶绿体发育进程受阻[14-15]两类。高等植物中,叶绿素的生物合成途径已基本明确[16-17],水稻中,已克隆的参与叶绿素合成途径的基因分别为编码镁离子螯合酶D、I、H三个亚基的、和基因[18-19],编码镁离子原卟啉Ⅸ单酯环化酶的基因[2],编码脱酯基叶绿素酸酯a乙烯基还原酶的基因[20],编码NADPH-原叶绿素酸酯氧化还原酶A和B的基因[21]和[22],编码叶绿素合成酶的基因[23],编码叶绿素酸酯a加氧酶的基因[12]和[13],参与这个途径的任何一个基因发生改变都会导致叶色变异。
叶绿体发育受阻不仅直接影响叶绿体的发育,同时会间接影响叶绿素的合成。控制叶绿体发育的基因主要有以下几类:一是质体转录复合物,如[24]、[25]、[3]、[26];二是PPR蛋白,如[8]、[27]、[28];三是叶绿体蛋白酶,如[29]、[30];四是信号转导相关基因,如[31]。由于叶色形成过程十分复杂,涉及到多方面的生理生化反应,因此叶色变异的分子机制仍然不是十分清楚,挖掘、鉴定新的叶色突变相关基因对进一步阐明水稻叶色调控的分子机制具有十分重要的意义。
本研究从常规粳稻镇糯19后代中发现一个移栽后叶色转成白条纹的自然变异突变体。对该突变体移栽前后的叶色表型、叶色转变条件、光合特性等进行分析,并构建群体对突变性状进行了遗传分析和基因定位,同时从植物生理、细胞和分子水平阐述的突变机理,以期为该基因克隆和功能分析奠定基础,揭示调控叶色的分子机制。
1 材料与方法
1.1 供试材料
来源于镇糯19后代自然突变,经10代连续自交获得,移栽后出现的白条纹性状在江苏句容、海南陵水都能稳定遗传。
1.2 wltt表型特征及主要农艺性状调查
在江苏丘陵地区镇江农业科学研究所行香基地种植和野生型亲本。5月15日播种,5叶1心期移栽,单本栽插,常规田间管理,观察在不同时期的叶色变化。成熟期分别取和野生型亲本各5株,考查株高、单株有效穗、剑叶长、穗长、每穗总粒数、千粒重和结实率等主要农艺性状。
1.3 剪根、光照和温度处理
水稻幼苗在水培条件下生长,营养液配制采用国际水稻所营养液配方[32],放置在光照培养箱(HP400GS-C)中,温度设置为20℃和30℃,光照强度设置为25 000 lx和6 250 lx,光照和黑暗各12 h,温度和光照强度两两组合,利用4台光照培养箱设置不同温度和光照强度。幼苗生长3叶1心时,剪根,留1 cm左右,继续生长2个叶龄,进行拍照和色素含量测定。
1.4 色素含量测定
取突变体与其野生型新完全展开叶,去掉中间叶脉,测定方法参照Wu等[23]。后续实验均在分蘖期移栽后表型出现时期进行。
1.5 光合速率测定
利用光合速率测定仪LI-6400XTOPEN 6.1测定自然条件下野生型和突变体正常移栽后分蘖期新生叶片的净光合速率,选取红蓝光源,叶室内的光照强度是1500 μmol/(m2·s),CO2浓度为400~420 μmol/mol, 叶片温度为25℃~28℃;叶室内空气湿度为40%~60%,将叶片夹入叶室后,数据稳定进行测量,在上午9:00−11:00进行测定,测定方法详见说明书。野生型和突变体各测5株,每株测3次。
1.6 叶绿体透射电镜分析
利用透射电镜(Transmission Electron Microcope, TEM)进行叶绿体超微结构观察。移栽后15 d,分别取突变体和野生型新完全展开叶的中上部,去掉中间叶脉,固定于含有2.5%的戊二醛溶液中,抽真空,4℃下固定12 h,冲洗后用1%锇酸溶液(pH 7.2)后固定4 h,经30%、50%、70%、80%、90%、100%酒精梯度脱水后,环氧树脂SPURR包埋、聚合。之后用LKB-V型切片机将样品切成1 μm厚的切片,醋酸铀染色后,经透射电镜(JEM-1230)观察、照相。
1.7 遗传分析及作图群体构建
用突变体和野生型亲本正反交得到F1,自交获得F2种子,并在江苏丘陵地区镇江农业科学研究所行香基地种植,移栽后统计F2群体中突变表型和正常表型的植株数,计算分离比,并进行χ2测验。用与籼稻9311杂交产生的F2作为定位群体,从F2群体中选取移栽后表现白条纹的单株(共3364份),进行基因定位。
1.8 基因定位
采用SDS法[33]提取水稻基因组DNA。利用均匀分布于水稻12条染色体上的InDel标记(引物由南京金斯瑞生物科技有限公司合成)进行亲本多态性分析。用有多态的引物对/9311的F2群体中选取的移栽后表现白条纹的单株进行连锁分析及精细定位,所用定位引物见表1。PCR扩增产物用8%聚丙烯酰胺凝胶电泳进行检测。
1.9 叶绿素合成和叶绿体发育相关基因的表达分析
使用天根公司的植物RNA小量提取试剂盒,对野生型和突变体移栽后15 d的新完全展开叶提取RNA,提取步骤详见说明书。反转录步骤参照TaKaRa公司(日本)的说明书。实时PCR 20 μL反应体系如下:反转录产物2 μL,10 µmol/L的正、反向引物0.4 μL,SYBRGreen混合物(TaKaRa公司) 10 μL,最后补去离子水至20 μL。反应在ABI 7500 实时 PCR仪上进行,反应程序参照Applied Biosystems操作手册,水稻的基因作为内参。每个样品设置3个重复。按2方法分析基因的相对表达量[34]。
2 结果与分析
2.1 突变体wltt的表型鉴定
通过对突变体的叶色观察,发现在苗期,突变体与野生型无显著差异,叶片表现出正常绿色(图1-A)。6月10日移栽,15 d(大田日均温均值24.5℃)后,突变体新生叶片表现出白条纹症状,而老叶维持叶色不变(图1-B);至成熟期,突变体叶色恢复正常(图1-D);7月10日移栽,15 d后(大田日均温31.7℃)突变体表型与野生型无差异;在齐穗期进行移栽,15 d后(大田日均温24.3℃)再生分蘖同样表现出白化(图1-C);而采用直播(不移栽)的方式种植的野生型和突变体植株全生育期叶色无明显差异(图1-E)。上述结果表明,表型的产生与移栽有关,且移栽后的相对低温对于白条纹表型的形成起着重要的作用。与野生型相比,正常移栽的突变体(6月10日移栽)新生叶片净光合速率比野生型显著降低(图1-F)。成熟期的农艺性状测定表明,株高明显下降(图1-D, 表2),同时穗长、剑叶长和每穗粒数亦明显降低,表明突变体的叶色变异显著影响了植株的生长发育。

表1 基因定位引物
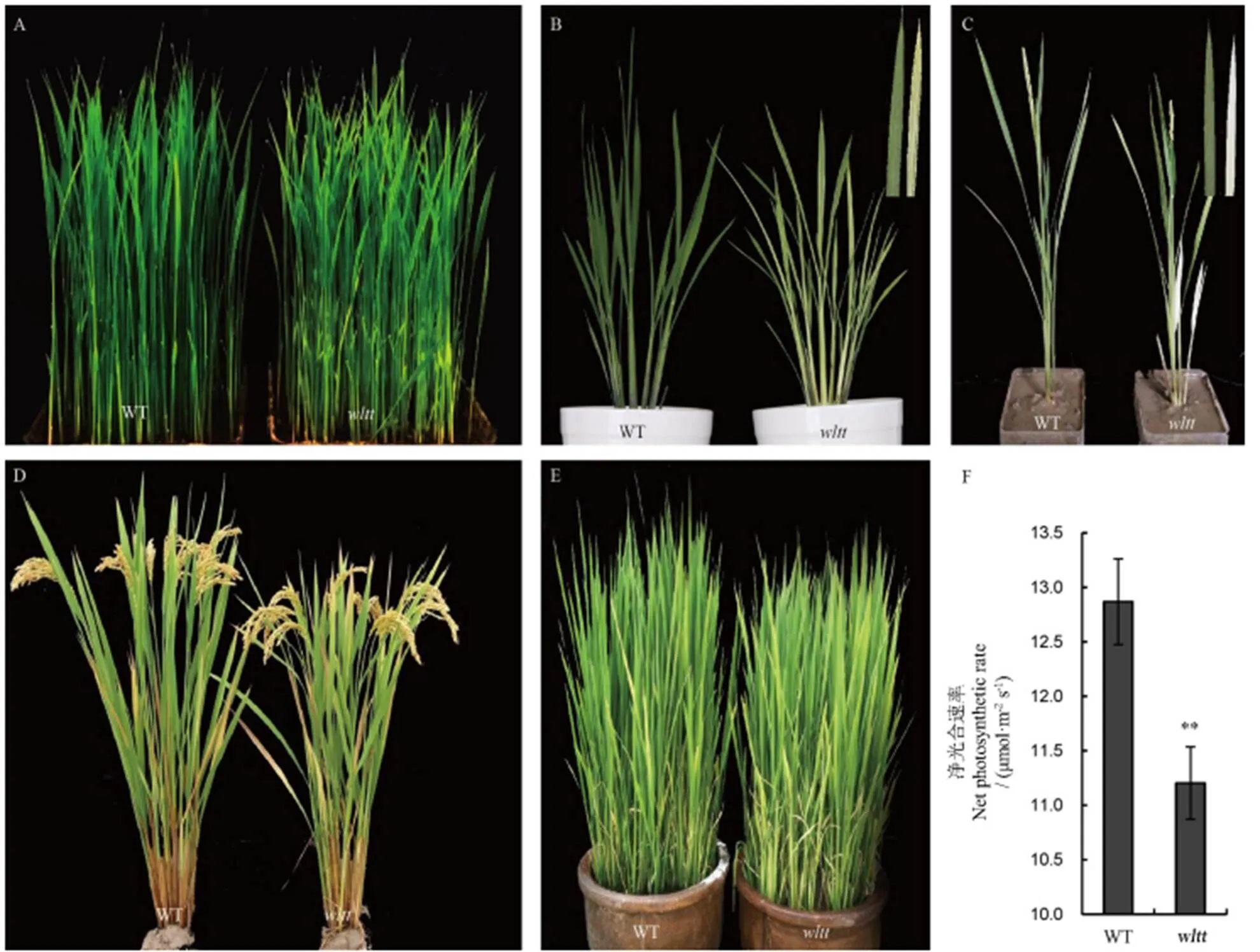
A−苗期;B−分蘖期,移栽后15 d,右上角为野生型(左)和突变体(右)的新完全展开叶;C−抽穗期移栽后15 d,右上角为野生型(左)和突变体(右)的再生分蘖的叶片;D−6月10日正常移栽的成熟期植株;E−直播条件下分蘖期的植株;F−正常移栽分蘖期植株新生叶片的净光合速率。WT−野生型。**表示经t测验后,P<0.01下差异显著。
Fig. 1. Phenotypic characterization of themutant and its wild type(WT).
2.2 突变体与野生型的色素含量测定
通过测定突变体及其野生型叶片的叶绿素和类胡萝卜素含量发现,苗期的各色素含量与野生型无显著差异;分蘖期移栽15 d后,的叶绿素a、叶绿素b、类胡萝卜素含量及总的色素含量明显下降,与野生型相比,差异极显著;而直播条件下,分蘖期的突变体的各色素含量与野生型相比,差异不显著(图2)。
2.3 突变体中叶绿体的超微结构变化
利用透射电镜(TEM)观察了移栽转色后野生型和突变体的叶绿体超微结构,结果显示,突变体的叶肉细胞与野生型相比(图3-A, C),大部分叶肉细胞不正常,观察不到叶绿体(图3-D),但也有少数叶肉细胞能观察到叶绿体(图3-E),且叶绿体中含有丰富的类囊体,其片层结构与野生型无显著差异(图3-F)。这一结果与白条纹突变性状(白绿相间)一致。
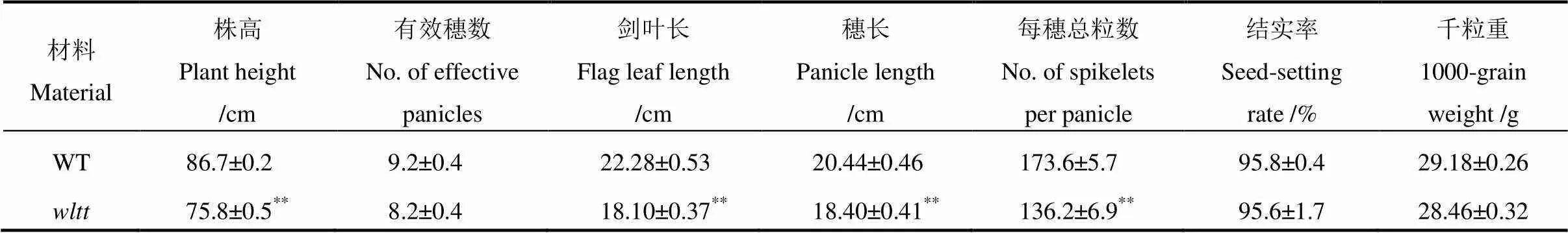
表2 野生型与突变体wltt的主要农艺性状比较
**表示经测验后,在<0.01下差异显著。
**The difference between the wild type andis significant at0.01 level according to Student’stest.
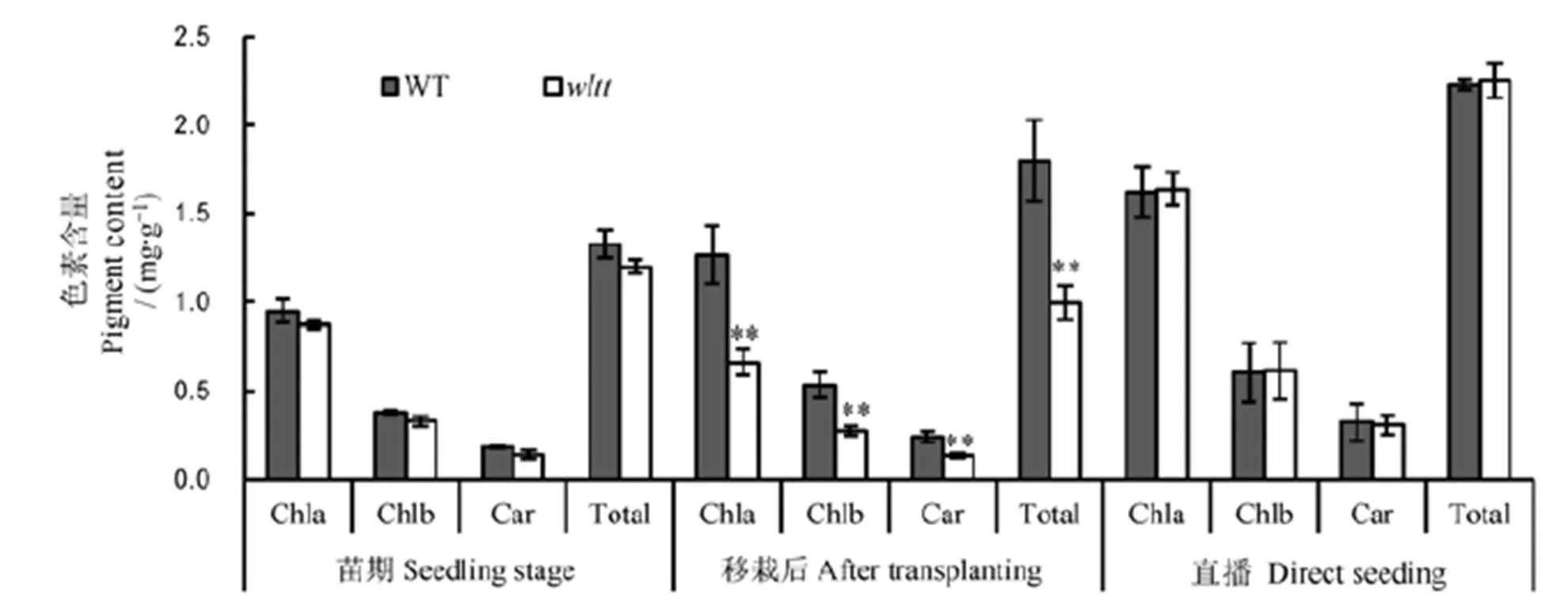
Chla-叶绿素a;Chlb-叶绿素b;Car-类胡萝卜素;Total-总色素含量。误差线表示3次独立实验的标准差。**表示野生型与突变体间差异达0.01显著水平(t测验)。
Fig. 2. Pigment contents in leaves ofmutant and its wild type(WT).
2.4 光合系统和叶绿体发育相关基因的表达分析
叶绿体是植物进行光合作用的场所,为研究该突变是否影响叶绿体发育,我们分析了移栽前后叶绿体发育和部分光合系统相关基因的表达(图4)。结果表明,在突变体中,叶绿体发育相关基因,如编码转运RNA聚合酶的家族的基因(、和),低温移栽后只有表达极显著上调表达(图4-A);而移栽前,、和全部上调表达,均达到极显著水平(图4-B)。叶绿体能量代谢相关酶编码基因,如编码1,5-二磷酸核酮糖羧化酶/加氧酶的大小亚基和;编码NADH脱氢酶的和及编码ATP合酶的和在突变体中低温移栽后全部下调(图4-A),而移栽前只有和在突变体中上调表达,其他基因无显著差异(图4-B)。在突变体中,光合系统相关基因,如编码光系统Ⅱ和Ⅰ蛋白复合物的基因家族的和基因在低温移栽后全部下调表达,而移栽前这些基因的表达无显著差异。这些结果表明,叶绿体发育相关基因的表达模式在移栽前后差异显著,移栽后大量的基因表达量下调,这很可能导致叶绿体发育缺陷。
2.5 叶绿素合成相关基因的表达分析
高等植物中叶绿素的合成过程十分复杂,突变体的色素含量比野生型显著降低,叶绿素的合成受到抑制,我们进一步分析了移栽前后突变体中叶绿素合成相关基因的表达水平(图5)。结果表明,突变体中,叶绿素生物合成的三个限速酶基因,如(编码谷氨酸-tRNA还原酶)、(编码镁离子螯合酶D亚基)、(编码镁离子螯合酶Ⅰ亚基)、(编码叶绿素合成酶)在低温移栽后均下调表达(图5-A);而移栽前和表达下调,和表达无差异(图5-B)。其他的叶绿素生物合成基因,如(编码谷氨酸-1-半缩醛氨基转移酶)、(编码δ-氨基酮戊酸脱水酶)、(编码原脱氨酶)、(编码尿卟啉原Ⅲ脱羧酶)、(编码粪卟啉Ⅲ氧化酶)、(编码镁离子原卟啉Ⅸ甲基转移酶)、(编码镁离子原卟啉Ⅸ单甲基脂环氧化酶)、(编码脱酯基叶绿素酸酯a乙烯基还原酶)、(编码NADPH-原叶绿素酸酯氧化还原酶)、(编码叶绿素合成酶),低温移栽后均下调表达(图5-A);移栽前基因和的表达水平在突变体中下调(图5-B)。这些结果表明,突变体的部分叶绿素合成相关基因的表达在移栽前后均有所下调,但似乎对移栽后的基因表达影响更大。

A~C为野生型的叶绿体超微结构;D~F为突变体wltt中同时含有异常结构的叶绿体(D)和正常结构的叶绿体(E~F)。Cp-叶绿体; Thy−类囊体; OB-嗜锇体。
Fig. 3. Transmission electron microscopic (TEM) images of chloroplast ultrastructure in the wild type and themutant.
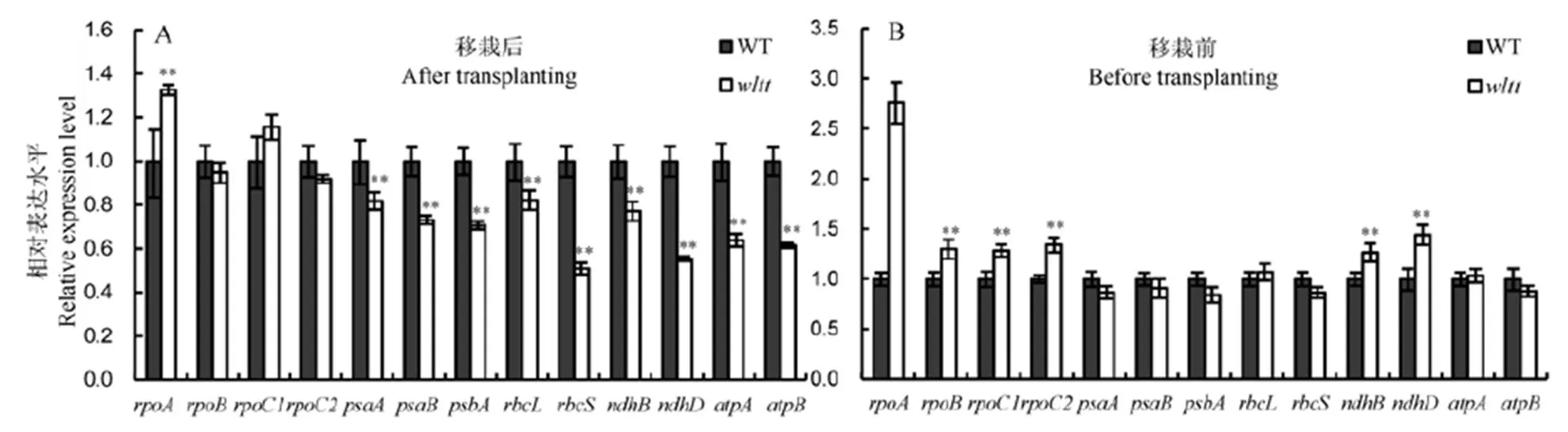
误差线表示3次独立实验的标准差。**表示野生型与突变体间差异达0.05和0.01显著水平(t测验)。
Fig. 4. Expression analysis of genes associated with chloroplast development and photosynthetic system in the wild type(WT) andmutant.
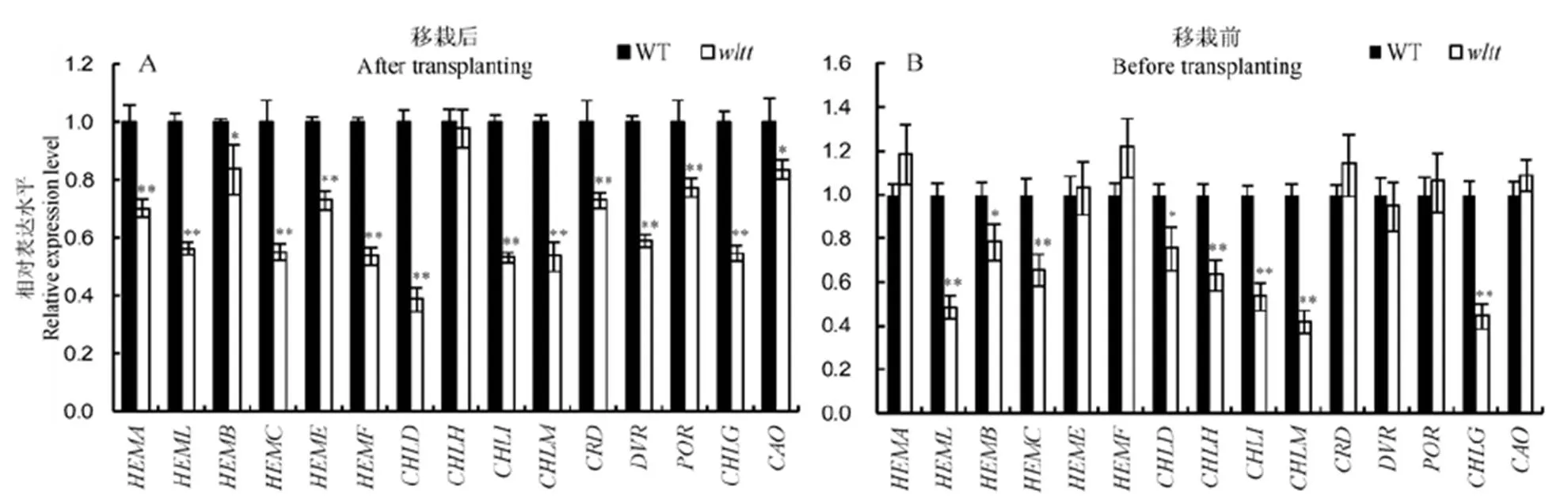
误差线表示3次独立实验的标准差。*和**分别表示野生型与突变体间差异达0.05和0.01显著水平(t测验)。
Fig. 5. Expression analysis of genes associated with chlorophyll biogenesis in the wild type(WT) andmutant.
2.6 突变体叶片转色的影响因素
连续两年观察发现,在梅雨季节结束之前(相对低温,弱光照)移栽心叶出现白条纹表型,而在梅雨季节后(高温,强光照)移栽无此表型。为研究叶片转色与机械损伤、光照、温度的关系,对幼苗进行剪根处理。结果表明,不进行剪根处理,在不同光照(25 000 lx和6 250 lx)和温度(20℃和30℃)条件下(两两组合),表型与野生型无差异(图6-A);在20℃下,光照强度在25 000 lx(图6-B)和6520 lx(图6-C)条件下,新生叶片均出现白条纹症状,同时,的叶绿素a、叶绿素b、类胡萝卜素含量及总色素含量明显下降,与野生型相比,差异极显著(图7);在30℃下,光照强度在25 000 lx和6250 lx条件下,突变体新生叶片均未转色,相应的的各色素含量与野生型相比无显著差异(图7)。这些结果表明,突变体新生叶片出现白条纹表型是由根系损伤和低温(20℃)引起的,与光照强度无关。

A-不进行剪根处理;B, C-剪根处理10 d后,20℃条件下,光照强度为25 000 lx(B)和6250 lx(C);D, E-剪根处理10 d后,30℃条件下,光照强度为25 000 lx(D)和6250 lx(E)。
Fig. 6. Effects of environmental conditions on leaf color variation.
2.7 突变体的遗传模式及基因定位
为确定突变体的遗传特性,我们进行了正反交实验并统计了后代的分离比例。结果表明,无论突变体用作母本还是父本,相应的F1植株移栽后新生叶片均表现出正常绿色,其自交后代F2群体移栽后新生叶片颜色表现出明显分离,且正常绿色与白条纹植株数比值符合3∶1(表3),这说明该性状是由一对隐性核基因控制的。
将突变体与籼稻9311杂交,从其F2代挑取移栽后表现为白条纹的单株共3364株作为定位群体,利用均匀分布在水稻12条染色体上的InDel标记将基因初定位在第2染色体着丝粒附近区域,两端标记为I2-5和I2-8,遗传距离分别是50.0 cM和58.4 cM(图8)。为精细定位,利用Primer 5.0软件,开发出两亲本之间有多态的引物11对(表1),最终将基因定位于标记L22和L26之间,物理距离为853 kb(图8),标记I2-6为共分离标记。
3 讨论
本研究中的叶色突变体的叶色表型为绿-白-绿,与之前Shi等[35]获得的移栽敏感叶绿体发育缺陷突变体的表型有相似之处,即在移栽15 d后,新生的3~4片叶表现出白化表型,随后的叶片恢复绿色,表型均在移栽后出现。不同的是,基因位于第11染色体,编码ABC转运蛋白,在黑暗条件下(移栽后的叶原基处于土壤中)的质体发育中发挥重要的作用;突变体即使经剪根处理在水培(光照补偿)条件下,移栽后也不出现白条纹叶的表型,与光照有关,与温度无关。本研究中,突变体经剪根处理在水培条件下移栽,低温(20℃)生长15 d,叶片出现白条纹症状;而在较高温度(30℃)无突变表型,并与光照强度无关;在大田条件下(叶原基处于黑暗条件),也表现出移栽低温敏感特性。因此,突变体的叶色变异是由低温(20℃)条件下根系损伤造成的,与光照无关。然而,突变体的低温敏特性,又有别于其他的叶色低温敏突变体;已成功克隆的5个叶色低温敏突变基因,即[36]、[37]、[38]、[39]和[40],这些基因突变后,对应的突变体均是在低温条件下出现白条纹表型,在高温条件下叶色恢复正常;突变体只有根系损伤后才表现出低温敏特性。基于本材料的特殊性,该突变体为研究水稻叶片生长发育、叶绿体发育及移栽后根系重建等机理提供了理想材料。
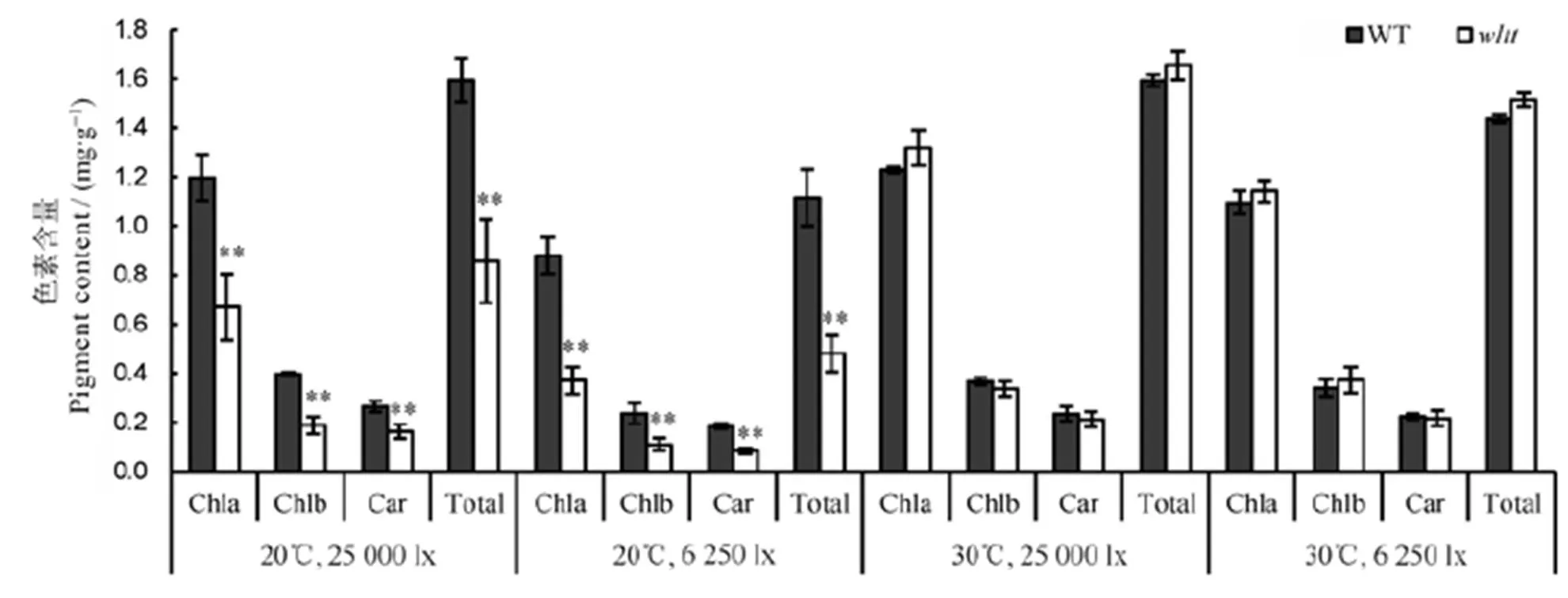
误差线表示3次独立实验的标准差。**表示野生型与突变体间差异达0.01显著水平(t测验)。
Fig. 7. Effects of environmental conditions on pigment contents after root cutting treatment.

表3 野生型与wltt突变体的F2代分离比
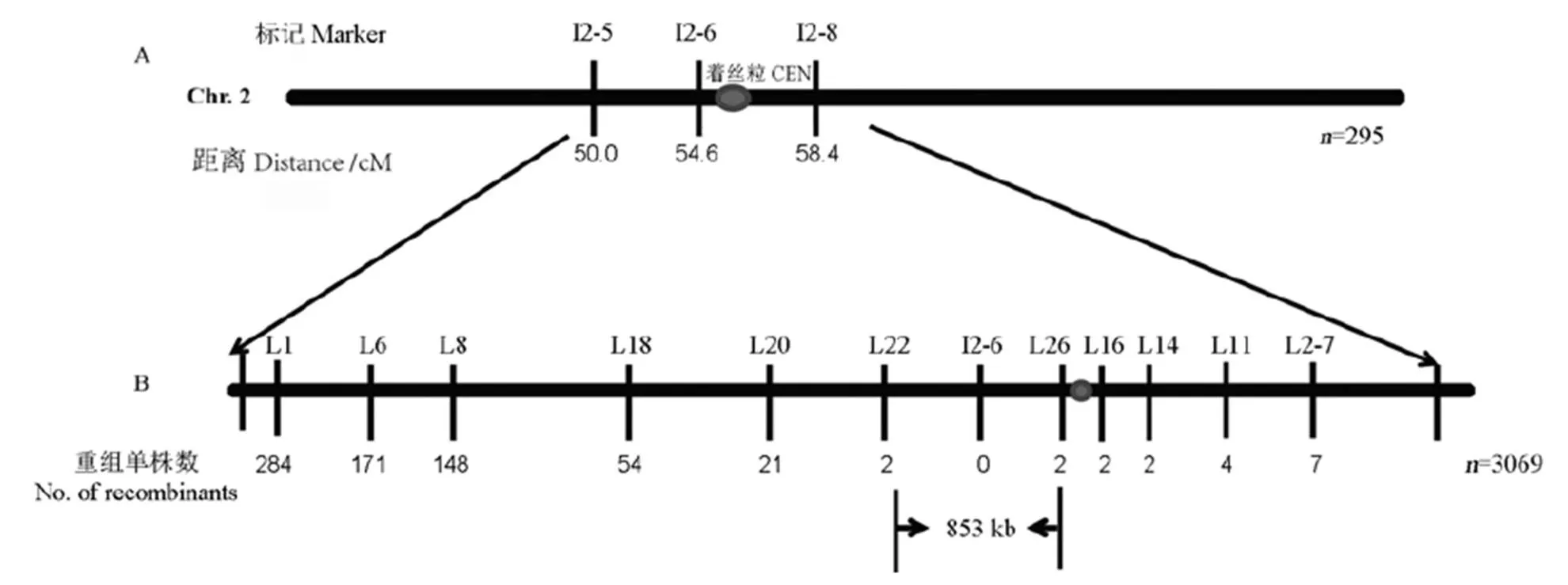
A-基因位点初定位在第2染色体着丝粒附近InDel标记I2-5和I2-8之间;B-基因位点精细定位到标记L22和L26之间,物理距离853 kb。n为定位所用的白条纹叶植株数。
Fig. 8. Location ofon rice chromosome 2.
叶绿体是植物特有的细胞器,也是植物进行光合作用的主要场所。通过对叶绿体透射电镜观察,发现突变体中既含有正常的叶绿体结构,也含有只存在嗜锇体、无类囊体片层结构的叶绿体,说明突变体的叶绿体发育受到影响。光反应相关蛋白编码基因表达量的改变会影响叶绿体光系统的形成,、编码PSⅠ合成相关亚基[41],其中PsaA和PsaB蛋白结合形成光合电子传递链的最初电子供体P700[42],基因用于编码光系统Ⅱ核心复合物;在突变体中,低温移栽后这些基因表达均下调,说明叶绿体的光系统受到影响。叶绿体中mRNA的转录对其发育至关重要,mRNA转录需要RNA聚合酶的参与,植物叶绿体内有4个基因编码RNA聚合酶:、、和;RNA聚合酶基因缺失后,植株色素含量降低,叶绿体内膜系统发育不完整[43];移栽前,突变体中上述4个基因全部上调,推测基因突变后反馈增强了RNA聚合酶的转录,以弥补的缺陷对叶绿体发育的影响;而低温移栽后只有表达略有上调,其他3个基因的表达量与野生型无差异,因而与移栽前相比,对叶绿体发育缺陷的补偿效应减弱甚至消失,从而造成了叶绿体发育的缺陷。核糖体大、小亚基参与叶绿体基因蛋白翻译过程,敲除叶绿体核糖体小亚基编码基因后,叶片表现出白化,植株不能存活[44];核糖体大亚基编码基因缺失后,叶片发育受到影响,植株光合速率下降[45];在突变体中,低温移栽后核糖体大、小亚基均表达下调,这与其表现出白条纹症状及光合速率下降相吻合。ATP合酶与NADH脱氢酶参与叶绿体内的能量代谢,小麦叶绿素缺失突变体中,白化株的家族基因全部表达下调[46];在突变体中,低温移栽后基因、、和均表达下调,则推测光合电子传递中的耗能过程不能正常进行,而移栽前和表达上调,推测基因突变后首先激活了NADH脱氢酶的转录,随着表型的出现,该基因转录水平下降。因此,控制突变体叶片转色的基因可能是叶绿体发育的关键基因。叶绿体发育受阻间接影响叶绿素合成,在突变体中,低温移栽后参与叶绿素生物合成的14种酶的编码基因表达大部分下调,叶绿素的生物合成受到抑制,这与突变体色素含量下降一致。而移栽前,部分叶绿素合成相关基因转录水平下调,这表明基因突变抑制了部分叶绿素合成相关基因的表达,与表型出现与否无直接关联。
本研究通过/9311的F2群体,将基因定位于第2染色体着丝粒附近,物理距离相距853 kb。由于临近着丝粒区交换冷点,尽管使用了较大的定位群体,进一步缩小区间仍存在一定困难。目前,在水稻第2染色体上已经报道了12个与叶色相关的基因,其中表现白化或白条纹性状的有3个,分别是(t)[47]、[48]和[49]。(t)突变体的表型是在3叶期白化,4叶期恢复为正常绿色,突变体表现为苗期叶片白色和淡绿色横向相间的斑马叶,突变体幼苗白化死亡,这3个基因均不在突变体基因定位的区间内;而且,该区间没有叶色相关基因的报道,表明为一个新的叶色基因。进一步克隆和鉴定的功能,将有助于提高我们对环境因子(如温度、植伤诱导等)影响叶绿体发育的机制的深入理解。
[1] Leister D. Chloroplast research in the genomic age., 2003, 19(1): 47-56.
[2] Kong W, Yu X, Chen H, Liu L, Xiao Y, Wang Y, Wang C, Lin Y, Yu Y, Wang C, Jiang L, Zhai H, Zhao Z, Wan J. The catalytic subunit of magnesium-protoporphyrin IX monomethyl ester cyclase forms a chloroplast complex to regulate chlorophyll biosynthesis in rice., 2016, 92(1-2): 177-191.
[3] Wang L, Wang C, Wang Y, Niu M, Ren Y, Zhou K, Zhang H, Lin Q, Wu F, Cheng Z, Wang J, Zhang X, Guo X, Jiang L, Lei C, Wang J, Zhu S, Zhao Z, Wan J. WSL3, a component of the plastid-encoded plastid RNA polymerase, is essential for early chloroplast development in rice., 2016, 92(4/5): 581-595.
[4] Zhang Z, Tan J, Shi Z, Xie Q, Xing Y, Liu C, Chen Q, Zhu H, Wang J, Zhang J, Zhang G.that encodes the sole octotricopeptide repeat protein is responsible for chloroplast development., 2016, 171(2): 1182-1191.
[5] Fambrini M, Castagna A, Vecchia F D, Degl Innocenti E, Ranieri A, Vernieri P, Pardossi A, Guidi L, Rascio N, Pugliesi C. Characterization of a pigment-deficient mutant of sunflower (L.) with abnormal chloroplast biogenesis, reduced PSII activity and low endogenous level of abscisic acid., 2004, 6: 645-650.
[6] Agrawal G K, Yamazaki M, Kobayashi M, Hirochika R, Miyao A, Hirochika H. Screening of the rice viviparous mutants generated by endogenous retrotransposoninsertion. Tagging of a zeaxanthin epoxidase gene and a novel ostatc gene., 2001, 125(3): 1248-1257.
[7] Parks B M, Quail P H. Phytochrome-deficientandlong hypocotyl mutants ofare defective in phytochrome chromophore biosynthesis., 1991, 3(11): 1177-1186.
[8] Su N, Hu M L, Wu D X, Wu F Q, Fei G L, Lan Y, Chen X L, Shu X L, Zhang X, Guo X P, Cheng Z J, Lei C L, Qi C K, Jiang L, Wang H, Wan J M. Disruption of a rice pentatricopeptide repeat protein causes a seedling- specific albino phenotype and its utilization to enhance seed purity in hybrid rice production., 2012, 159(1): 227-238.
[9] 谭炎宁, 孙学武, 袁定阳, 孙志忠, 余东, 何强, 段美娟, 邓华凤, 袁隆平. 水稻单叶独立转绿型黄化突变体的鉴定与基因精细定位. 作物学报, 2015, 41(6): 831-837.
Tan Y N, Sun X W, Yuan D Y, Sun Z Z, Yu D, He Q, Duan M J, Deng H F, Yuan L P. Identification and fine mapping ofgenein rice (L.)., 2015, 41(6): 831-837. (in Chinese with English abstract)
[10] 钱前, 朱旭东, 曾大力, 张小惠, 严学强, 熊振民. 细胞质基因控制的新特异材料白绿苗的研究. 作物品种资源, 1996(4): 11-12.
Qian Q, Zhu X D, Zeng D L, Zhang X H, Yan X Q, Xiong Z M. The study on a new special material, white-green rice which controlled by plasma gene., 1996(4): 11-12. (in Chinese).
[11] 李贤勇, 王楚桃, 李顺武, 何永歆, 陈世全. 一个水稻高叶绿素含量基因的发现. 西南农业学报, 2002, 15(4): 122-123.
Li X Y, Wang C T, Li S W, He Y X, Chen S Q. The discovery of a high chlorophyll content gene in rice., 2002, 15(4): 122-123. (in Chinese with English abstract)
[12] Lee S, Kim J H, Yoo E S, Lee C H, Hirochika H, An G. Differential regulation of chlorophyll a oxygenase genes in rice., 2005, 57(6): 805-818.
[13] Yang Y L, Xu J, Huang L C, Leng Y J, Dai L P, Rao Y C, Chen L, Wang Y Q, Tu Z J, Hu J, Ren D Y, Zhang G H, Zhu L, Guo L B, Qian Q, Zeng D L., encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice., 2016, 67(5): 1297-1310.
[14] Kusumi K, Yara A, Mitsui N, Tozawa Y, Iba K. Characterization of a rice nuclear-encoded plastid RNA polymerase gene., 2004, 45(9): 1194-1201.
[15] Sugimoto H, Kusumi K, Tozawa Y, Yazaki J, Kishimoto N, Kikuchi S, Iba K. Themutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation., 2004, 45(8): 985-996.
[16] Beale S I. Green genes gleaned., 2005, 10(7): 309-312.
[17] Nagata N, Tanaka R, Satoh S, Tanaka A. Identification of a vinyl reductase gene for chlorophyll synthesis inand implications for the evolution of., 2005, 17(1): 233-240.
[18] Goh C H, Satoh K, Kikuchi S, Kim S C, Ko S M, Kang H G, Jeon J S, Kim C S, Park Y. Mitochondrial activity in illuminated leaves of chlorophyll-deficient mutant rice () seedlings., 2010, 4(4): 281-291.
[19] Zhang H T, Li J J, Yoo J H, Yoo S C, Cho S H, Koh H J, Seo H S, Paek N C. Riceandencode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development., 2006, 62(3): 325-337.
[20] Wang P R, Gao J X, Wan C M, Zhang F T, Xu Z J, Huang X Q, Sun X Q, Deng X J. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice., 2010, 153(3): 994-1003.
[21] Sakuraba Y, Rahman M L, Cho S H, Kim Y S, Koh H J, Yoo S C, Paek N C. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions., 2013, 74(1): 122-133.
[22] Yang Q S, He H, Li H Y, Tian H, Zhang J J, Zhai L G, Chen J D, Wu H, Yi G J, He Z H, Peng X X. NOA1 functions in a temperature-dependent manner to regulate chlorophyll biosynthesis and rubisco formation in rice., 2011, 6(5): e20015.
[23] Wu Z M, Zhang X, He B, Diao L P, Sheng S L, Wang J L, Guo X P, Su N, Wang L F, Jiang L, Wang C M, Zhai H Q, Wan J M. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis., 2007, 145(1): 29-40.
[24] Yagi Y, Ishizaki Y, Nakahira Y, Tozawa Y, Shiina T. Eukaryotic-type plastid nucleoid protein pTAC3 is essential for transcription by the bacterial-type plastid RNA polymerase., 2012, 109(19): 7541-7546.
[25] Arsova B, Hoja U, Wimmelbacher M, Greiner E, Ustun S, Melzer M, Petersen K, Lein W, Bornke F Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: Evidence for an essential role in chloroplast development inand., 2010, 22: 1498-1515.
[26] Wang Y, Wang C , Zheng M, Lyu J, Xu Y, Li X, Niu M, Long W, Wang D, Wang H Y, William T, Wang Y, Wan J. WHITE PANICLE1, a Val-tRNA synthetase regulating chloroplast ribosome biogenesis in rice, is essential for early chloroplast development., 2016, 170(4): 2110-2123.
[27] Wu L, Wu J, Liu Y, Gong X, Xu J, Lin D, Dong Y. The rice pentatricopeptide repeat geneis needed for chloroplast development under cold stress., 2016, 9: 67.
[28] Tang J, Zhang W, Wen K, Chen G, Sun J, Tian Y, Tang W, Yu J, An H, Wu T, Kong F, Terzaghi W, Wang C, Wan J. OsPPR6, a pentatricopeptide repeat protein involved in editing and splicing chloroplast RNA, is required for chloroplast biogenesis in rice., 2017, 95(4/5): 345-357.
[29] Yue R, Wang X, Chen J, Ma X, Zhang H, Mao C, Wu P. A rice stromal processing peptidase regulates chloroplast and root development., 2010, 51(3): 475-485.
[30] Dong H, Fei G L, Wu C Y, Wu F Q, Sun Y Y, Chen M J, Ren Y L, Zhou K N, Cheng Z J, Wang J L, Jiang L, Zhang X, Guo X P, Lei C L, Su N, Wang H, Wan J M. A ricemutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants., 2013, 162(4): 1867-1880.
[31] Zhou S, Sawicki A, Willows R D, Luo M. C-terminal residues ofGUN4 are required for the activation of the ChlH subunit of magnesium chelatase in chlorophyll synthesis., 2012,586 (3): 205-210.
[32] Yoshida S, Forno D A, Cock J A H, Gomez K A. Laboratory Manual for Physiological Studies of Rice. Los Banos, Philippines: The International Rice Research Institute, 1976: 61.
[33] McCouch S R, Kochert G, Yu Z H, Wang Z Y, Khush G S, Coffman W R, Tanksley S D. Molecular mapping of rice chromosome., 1998, 76: 815-829.
[34] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method., 2001, 25(4): 402-408.
[35] Shi X, Chen S, Peng Y, Wang Y, Chen J, Hu Z, Wang B, Li A, Chao D, Li Y, Teng S. TSC1 enables plastid development under dark conditions, contributing to rice adaptation to transplantation shock., 2018, 60 (2): 112-129.
[36] Kensuke K, Shoko H, Hiroshi S, Yoko C, Osanu M, Koh I. Contribution of chloroplast biogenesis to carbon- nitrogen balance during early leaf development in rice., 2010, 123(4): 617-622.
[37] Hiroki S, Kensuke K, Ko N, Masahiro Y, Atsushi Y, Koh I. The rice nuclear gene,, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria., 2007, 52(3): 512-527.
[38] Gong X D, Su Q Q, Lin D Z, Jiang Q, Xu J L, Zhang J H, Teng S, Dong Y J. The riceencoding a novel pentatricopeptide repeat protein is required for chloroplast development during the early leaf stage under cold stress., 2014, 56(4): 400-410
[39] Jiang Q, Mei J, Gong X D, Xu J L, Zhang J H, Teng S, Lin D Z, Dong Y J. Importance of the riceencoding subunit of chaperonin protein 60 (Cpn60) for the chloroplast development during the early leaf stage., 2014, 215/216: 172-179.
[40] Song J, Wei X J, Shao G N, Sheng Z H, Chen D B, Liu C L, Jiao G A, Xie L L, Tang S Q, Hu P S. The rice nuclear geneencoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions., 2014, 84(3): 301-314
[41] Pakrasi H B. Genetic analysis of the form and function of photosystem Ⅰ and photosystem Ⅱ., 1995, 29: 755-776.
[42] Boudreau E, Takahashi Y, Lemieux C, Turmel M, Rochaix J D. The chloroplastandopen reading frames ofare required for the accumulation of the photosystem I complex., 1997, 16(20): 6095-6104.
[43] Santis-Maciossek G D, Kofer W, Bock A, Schoch S, Maier R M, Wanner G, Rüdiger W, Hans-Ulrich K, Herrmann R G. Targeted disruption of the plastid RNA polymerase genes,and: Molecular biology biochemistry and ultrastructure., 1999,18: 477-489.
[44] Rogalski M, Ruf S, Bock R. Tobacco plastid ribosomal protein S18 is essential for cell survival., 2006, 34: 4537-4545.
[45] Fleischmann T T, Scharff L B, Alkatib S, Hasdorf S, Schottler M A, Bock R. Nonessential plastid-encoded ribosomal proteins in tobacco: A developmental role for plastid translation and implications for reductive genome evolution., 2011, 23(9): 3137-3155.
[46] 夏家平, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 李军辉, 刘录祥. 小麦叶绿素缺失突变体的叶绿体基因差异表达分析. 中国水稻科学, 2012, 38 (11):2122-2130.
Xia J P, Guo H J, Xie Y D, Zhao L S, Gu J Y, Zhao S R, Li J H, Liu L X. Differential expression of chloroplast genes in chlorophyll-deficient wheat mutantderived from space mutagenesis., 2012(11): 2122-2130.
[47] Chen T, Zhang Y, Zhao L, Zhu Z, Lin J, Zhang S, Wang C. Fine mapping and candidate gene analysis of a green-revertible albino gene(t) in rice.2009, 36(2): 117-123.
[48] 李燕群, 钟萍, 高志艳, 朱柏羊, 陈丹, 孙昌辉, 王平荣, 邓晓建. 水稻斑马叶突变体的表型鉴定及候选基因分析.中国农业科学, 2014, 47(15): 2907-2915.
Li Y Q, Zhong P, Gao Z Y, Zhu B Y, Chen D, Sun C H, Wang P R, Deng X J. Morphological characterization and candidate gene analysis of zebra leaf mutantin rice., 2014, 47(15): 2907-2915.
[49] Lin D, Jiang Q, Zheng K, Chen S, Zhou H, Gong X, Xu J, Teng S, Dong Y. Mutation of the ricegene encoding plastid ribosomal protein L21 causes chloroplast developmental defects and seedling death., 2015, 17(3): 599-607.
Identification and Gene Mapping of aMutant in Rice
LIN Tianzi1, 2, SUN Liting2, GONG Hongbin2, WANG Yihua1, LIU Linglong1, ZHAO Zhigang1, JIANG Ling1, WAN Jianmin1,*
(State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University/Key Laboratory of Biology, Genetics and Breeding of japonica Rice in Mid-lower Yangtze River, Ministry of Agriculture/The Yangtze River Valley Hybrid Rice Collaboration Innovation Center/Jiangsu Collaboration Innovation Center for Modern Crop Production, Nanjing 210095, China;Zhenjiang Agricultural Research Institute, Jurong 212400, China;*)
【Objective】Isolation and characterization of leaf-color mutation related genes lays a firm theoretical foundation for dissecting the molecular mechanism underlying chloroplast development, chlorophyll biosynthesis, and photosynthesis in rice. 【Method】A spontaneous leaf-color mutant, termed aswas obtained from the progeny ofcultivar Zhennuo 19. The main agronomic traits of the wild type andwere determined at maturity. The pigment contents and ultrastructure of chloroplast of newly emerged leaves were analyzed at the seedling stage, fifteen days after transplanting, at the tillering stage under direct seeding. Genetic analysis was carried out by reciprocal cross of the wild type and. An F2population derived from the cross×9311 was used for gene mapping. Quantitative RT-PCR was carried out to analyze the relative expression of genes associated with chloroplast development and chlorophyll biogenesis in the wild type and themutant. 【Result】The white-striped leaves in themutant only emerged at 15 days after transplanting at low temperature such as 20℃. No white-striped leaf was observed under direct seeding treatment. However, leaves of the mutant developed normally at the late tillering stage. Simulation experiments showed that the mutant phenotype was caused by root injury at low temperature. Compared with the wild type, the pigment contents in white-stripe leaves of themutant were significantly decreased, accompanying by reduced photosynthetic rate. Simultaneously, most of the mesophyll cells had no chloroplasts. The expression levels of genes associated with chloroplast development, chlorophyll biosynthesis, and photosynthetic system were all down-regulated in the mutant. At maturity, the mutant was featured with reduced plant height, panicle length, flag leaf length and number of spikelets per panicle relative to its wild type. Genetic analysis revealed that the mutant phenotype was controlled by a single recessive nuclear gene. Moreover, thegene was mapped within an 853 kb region near the centromere on chromosome 2, between InDel markers L22 and L26, in which no gene related to leaf color was reported. 【Conclusion】is a key gene regulating leaf color after transplanting at low temperature, which plays an important role in chloroplast development.
rice; white-stripe leaf after transplanting; chloroplast development; gene mapping
10.16819/j.1001-7216.2018.8026
Q343.5; S511.034
A
1001-7216(2019)01-0001-11
2018-03-13;
2018-05-29。
国家重点研发计划资助项目(2017YFD0100400);江苏省重点研发计划资助项目(BE2015363);江苏省农业科技自主创新资金资助项目[CX(16)1029]。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
语文世界(小学版)(2021年5期)2021-06-04
农业科技通讯(2021年1期)2021-03-06
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
江苏农业科学(2016年2期)2016-04-11
