
不同香稻品种苗期耐高温的形态生理响应
2019-01-24 03:53:10冯活仪江浩林王孟唐湘如段美洋潘圣刚田华王树丽莫钊文
中国水稻科学 2019年1期
冯活仪 江浩林 王孟 唐湘如,2,3 段美洋,2,3 潘圣刚,2,3 田华,2,3 王树丽,2,3 莫钊文,2,3,*
不同香稻品种苗期耐高温的形态生理响应
冯活仪1江浩林1王孟1唐湘如1,2,3段美洋1,2,3潘圣刚1,2,3田华1,2,3王树丽1,2,3莫钊文1,2,3,*
(1华南农业大学 农学院,广州 510642;2农业部华南地区作物栽培科学观测实验站,广州 510642;3广东省丝苗香米科技创新中心,广州 510642;*通讯联系人,E-mail: scaumozhw@126.com)
【目的】本研究旨在分析不同香稻品种苗期耐高温的形态生理响应特征,筛选出耐热性较好的香稻品种。【方法】在盆栽条件下,于光照培养箱内,以超级常规香稻玉香油占为对照,研究了常规香稻品种巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号、美香占2号在苗期高温处理下的形态生理响应特征。于高温处理前后测定了8个香稻品种苗期各项形态生理指标。【结果】与玉香油占相比,高温胁迫下,巴斯马蒂的株高、单株鲜质量、单株干质量增幅更大,成活率和过氧化物酶活性更高,丙二醛含量反而下降了33.39%,表明巴斯马蒂的耐高温胁迫能力较好。湘晚籼13在高温处理下的单株干质量增幅与玉香油占相近,株成活率较玉香油占低,胞间二氧化碳浓度变幅较小,SOD活性和丙二醛含量的变幅在8个品种中均为最低,说明其耐高温胁迫的能力略低于玉香油占。农香18、象牙香占和美香占2号的成活率分别为13.33%、12.67%和16.00%。桂香占(4.33%)、中香1号(1.33%)在高温处理后的成活率最低,表明两者的耐高温能力均较差。【结论】8个品种耐热性从强到弱依次为巴斯马蒂、玉香油占、湘晚籼13、农香18、象牙香占、美香占2号、桂香占和中香1号。
香稻;耐高温;形态生理;幼苗
水稻是世界上主要粮食作物之一。水稻发源于低纬度地区,形成了适应高温生态环境的特性,但水稻的正常生长发育需要适宜的温度范围[1-3]。随着温室效应不断加剧,全球变暖也不断加剧,并且全球变暖这种现象将持续到21世纪末[4]。局部短期异常的高温频繁发生,高温胁迫作为一种自然逆境对植物的影响是复杂的[5]。相关研究表明,高温影响水稻的水分和养分吸收利用、膜系统的稳定性、酶促反应等,高温下膜系统的稳定性直接影响水稻的光合能力,成为限制水稻产量和品质的因素之一[6]。目前来说,在农业生产上应对高温的措施主要有化学调控、合理肥水管理、调整播期、加强田间管理、选育和培育耐高温品种[5,7-11]。在水稻对高温胁迫的响应上,国内外学者已就水稻不同生育时期的耐高温胁迫鉴定评价方法与指标、生理及遗传机理开展了大量的研究,但研究对象多为普通水稻品种,对香稻耐热性研究的相关报道较少[12]。黄显波等[6]发现杂交籼稻幼苗在38℃高温胁迫下抗氧化酶活性降低,丙二醛(MDA)含量升高。张晓晶等[13]通过对两个不同耐性的水稻幼苗进行7 d 40℃高温处理,发现高温处理促进了耐热性水稻幼苗的生长,表现为株高、根长以及地上部干质量显著增加。香稻品种对高温响应的形态生理基础目前鲜有报道。
朱雪梅等[14]对不同穗型水稻品种的耐高温性进行研究,发现高温耐性从高到低为重穗型品种、中穗型品种、轻穗型品种。本研究以超级常规香稻品种玉香油占为对照,比较另外7个常规香稻品种与玉香油占在苗期高温胁迫下形态生理响应,旨在对不同品种香稻苗期耐热性能进行初步的鉴定和形态生理基础分析评价,筛选出耐热性能较强的香稻品种并为香稻苗期的高温耐性机理研究提供依据。
1 材料与方法
1.1 供试材料
试验材料常规超级香稻玉香油占、常规香稻巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号、美香占2号均由华南农业大学农学院水稻研究室提供。巴斯马蒂和桂香占由华南农业大学农学院引进并繁留种子。玉香油占由广东省农业科学院水稻研究所以水稻中间材料TY36×IR100为母本,以水稻优质品系IR100为父本进行人工温汤杀雄杂交,经5代系谱选育而成。农香18是由湖南省水稻研究所以农香16为母本,以三合占为父本杂交选育而成。湘晚籼13是湖南省水稻研究所等单位选育的迟熟香型优质常规晚籼品种,父母本分别为湘晚籼6号和湘晚籼5号。象牙香占由台山市农业科学研究所以香丝苗126为母本,以象牙软占为父本杂交选育而成。中香1号是中国水稻研究所以80-66为母本,以矮黑为父本杂交选育的迟熟优质晚籼稻品种。美香占2号为由广东省农业科学院水稻研究所以Lement为母本,以丰澳占为父本经三次轮回杂交选育而成的感温型常规籼稻品种。
1.2 试验设计
试验于2015年6月在华南农业大学农学院院楼进行,所用的水稻种子进行浸种催芽后,每个品种选取长势一致的谷芽100粒播于上口径为16.0 cm,下口径为11.5 cm,高为15.5 cm的塑料桶内,桶内已提前装好2.5 kg的水稻泥。每个品种重复8次。置于光照培养箱中,光照16 h/黑暗8 h,温度28℃/25℃。待秧苗长至3叶期后,移入光照16 h/黑暗8 h,40℃培养箱中培养8 d。高温处理前后分别取样1次并测定相关形态生理指标。
1.3 各项指标的测定方法
1.3.1 秧苗素质的测定
1.3.1.1 株高的测定
采用随机取样法,每个重复选取10株秧苗,测定自茎基部至叶子最顶端的长度,取平均值。
1.3.1.2 单株鲜质量的测定
把测完株高的秧苗剪去根部,将10株秧苗用千分之一天平上称量鲜质量,计算单株鲜质量,重复4次。
1.3.1.3 单株干质量的测定
将测完鲜质量的秧苗放入鼓风干燥箱,于105℃下杀青30 min,80℃下烘干至恒质量,然后用千分之一分析天平称量每个重复的秧苗干质量。
1.3.1.4 株成活率的测定
高温处理后,测定剩余秧苗的株成活率。
1.3.2 光合参数的测定
于晴朗天气上午9:00-11:00,采用美国产LI-6400型便携式光合测定系统(Li-cor公司)测定光合参数净光合速率(n)、气孔导度(s)、胞间CO2浓度(i)和蒸腾速率(r)。大气条件为CO2浓度为400 μmol/mol,30℃,光照度1000 μmol/(m2·s)。测定最上面一片展开叶的中部,每个处理重复测定4次。
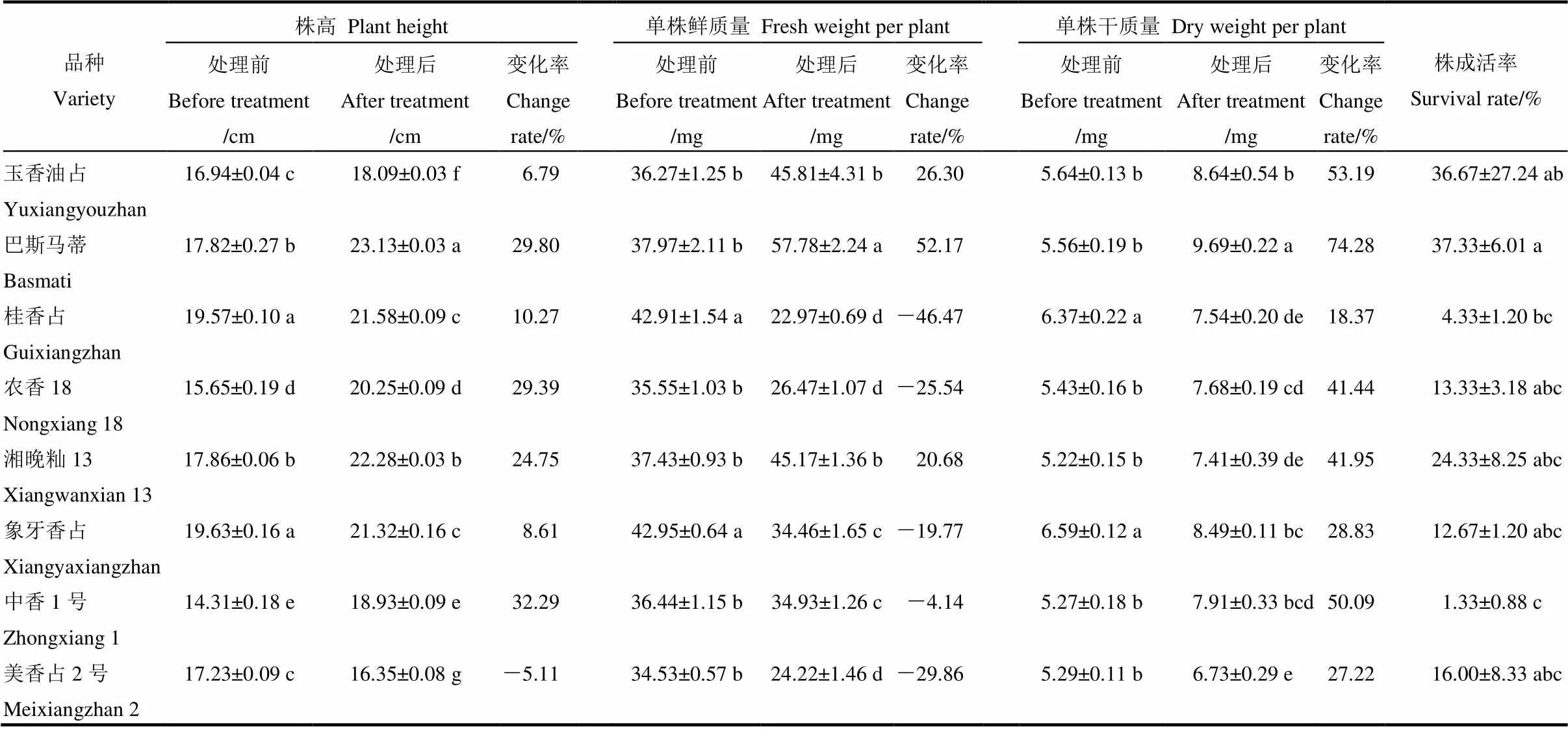
表1 高温胁迫对秧苗素质的影响
同列数据后不同小写字母表示差异显著(<0.05)。下同。
Different lowercase letters in the same column mean significant difference(<0.05). The same as below.
1.3.3 叶绿素荧光特性的测定
叶绿素荧光参数的测定使用TEACHING-PAM叶绿素荧光仪进行,测定所用秧苗经过黑暗处理30min后,选取植株最上面的第一片展开叶对其最小荧光强度(O)、最大荧光强度(m)、最大可变荧光强度(v)和最大光化学效率(v/m)等叶绿素荧光参数进行测定,每个处理重复测定4次。
1.3.4 抗氧化酶活性及丙二醛含量的测定
于高温处理前后各取一次样,每个处理取存活的地上部植株,液氮固定之后保存于-80℃冰箱中。参照李合生[15]测定秧苗体内抗逆性酶活性:超氧化物歧化酶(SOD)活性测定采用氯化硝基四氮蓝唑(NBT)光化学还原法,以抑制50%为1个酶活力单位U;过氧化物酶(POD)活性测定采用愈创木酚法,以470的变化值达0.1为一个相对酶活力单位U;过氧化氢酶(CAT)活性测定采用双氧水法,以240每1min的变化值达0.1为一个相对酶活力单位U;丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)氧化法。
1.4 数据分析
所有数据使用Microsoft Excel 2010处理,使用Statistix 8数据处理系统进行统计。各数值为4次重复的平均数,多重比较采用LSD法,结果采用字母标记法标记,相同小写字母的两组数据之间表示差异未达到5%显著水平。
2 结果与分析
2.1 高温胁迫对秧苗素质的影响
从表1可以看出,高温处理后,除美香占2号的株高下降了5.11%,其余品种的株高均有所提高,玉香油占、巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占和中香1号的株高分别增加了6.79%、29.80%、10.27%、29.39%、24.75%、8.61%和32.29%。在单株鲜质量上,高温处理后,桂香占、农香18、象牙香占、中香1号和美香占2号的单株鲜质量分别下降了46.47%、25.54%、19.77%、4.14%和29.86%,而玉香油占、巴斯马蒂和湘晚籼13的单株鲜质量分别增加了26.30%、52.17%和20.68%,其中,巴斯马蒂的增幅最大。在单株干质量方面,8个品种在高温胁迫下干质量均有所增加,其中,巴斯马蒂的增幅最大,为74.28%,其次是玉香油占,为53.19%。在株成活率上,巴斯马蒂的株成活率最高,桂香占和中香1号的株成活率最低,其余品种的成活率无显著差异。
2.2 高温胁迫对秧苗光合速率以及气体交换的影响
如表2所示,高温处理后,供试香稻品种秧苗的净光合速率(n)均显著下降,玉香油占、巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号、美香占2号的净光合速率分别下降了66.89%、83.85%、69.67%、89.73%、77.76%、96.43%、94.40%和93.43%,由此可见,玉香油占的降幅最小,其次是桂香占。在气孔导度(s)上,8个品种的秧苗在高温处理后气孔导度(s)均显著降低,玉香油占、巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号和美香占2号的气孔导度分别下降了41.84%、72.03%、67.59%、70.94%、58.01%、69.43%、94.35%和35.05%,可见,美香占2号的降幅是最小的,其次是玉香油占。在胞间二氧化碳浓度(i)方面,除桂香占在高温处理后降低2.78%外,其余品种在高温处理后均有所增加;玉香油占、巴斯马蒂、农香18、湘晚籼13、象牙香占、中香1号和美香占2号的i分别增加了46.85%、18.28%、8.94%、11.50%、25.94%、5.43%和59.79%,美香占2号的增幅是最大的,其次是玉香油占。在蒸腾速率(r)上,高温胁迫降低了秧苗的蒸腾速率;玉香油占、巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号和美香占2号的蒸腾速率分别下降了35.99%、68.55%、60.91%、65.49%、51.89%、65.86%、93.68%和35.55%,其中,美香占2号的降幅最小,其次是玉香油占。
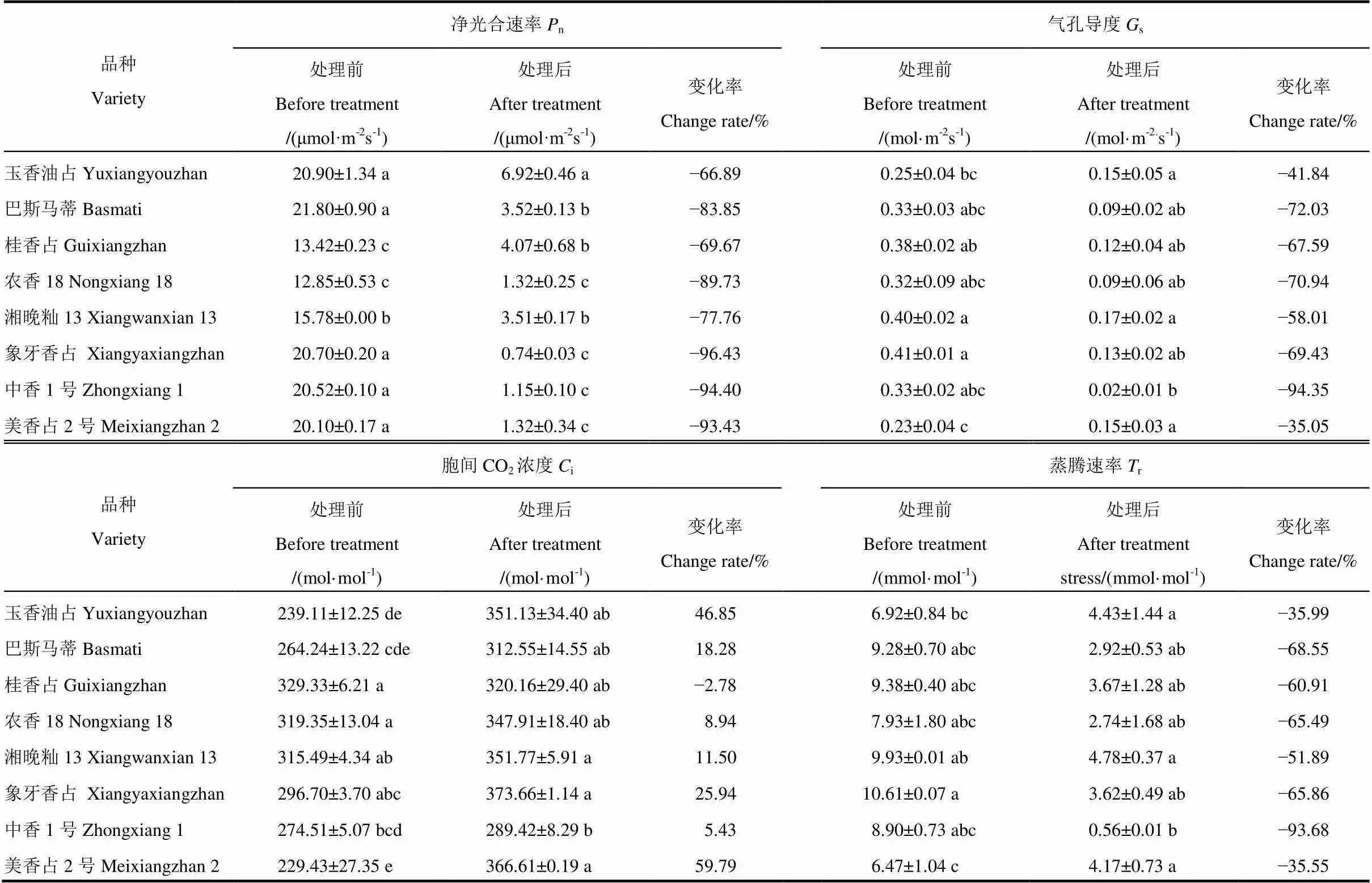
表2 高温胁迫对秧苗光合速率以及气体交换的影响
2.3 高温胁迫对秧苗叶绿素荧光特性的影响
从表3可以看出,高温处理后,8个品种间初始荧光产量(o)、最大荧光产量(m)以及最大可变荧光产量均无显著差异。在PSⅡ最大光化学量子产量(v/m)上,高温处理前,巴斯马蒂v/m值最高且显著高于象牙香占,而与其余品种无显著差异。高温处理后,8个品种的v/m均有所下降,但品种间无显著差异。
2.4 高温胁迫对秧苗抗氧化酶活性以及丙二醛含量的影响
如表4所示,高温处理后,除玉香油占的POD活性有所下降外,其余品种的POD活性均有所提高,POD活性增幅从大到小依次为桂香占、中香1号、湘晚籼13、巴斯马蒂、象牙香占、农香18、美香占2号。高温处理后,桂香占、象牙香占和中香1号的SOD活性分别提高了183.61%、9.27%和43.98%,而玉香油占、巴斯马蒂、农香18、湘晚籼13和美香占2号的SOD活性下降了83.66%、72.83%、13.20%、0.33%和66.68%。高温处理后8个品种的CAT活性均有所降低,其中,玉香油占的CAT活性减幅最大,为68.15%,巴斯马蒂、桂香占、农香18、湘晚籼13、象牙香占、中香1号和美香占2号的CAT活性分别降低了62.65%、65.23%、50.56%、40.85%、64.06%、54.40%和65.31%。在丙二醛(MDA)含量上,高温处理后,除巴斯马蒂降低33.39%外,其余品种的MDA含量均有所提高,按MDA增幅从大到小依次为桂香占、中香1号、象牙香占、农香18、湘晚籼13、美香占2号、玉香油占。
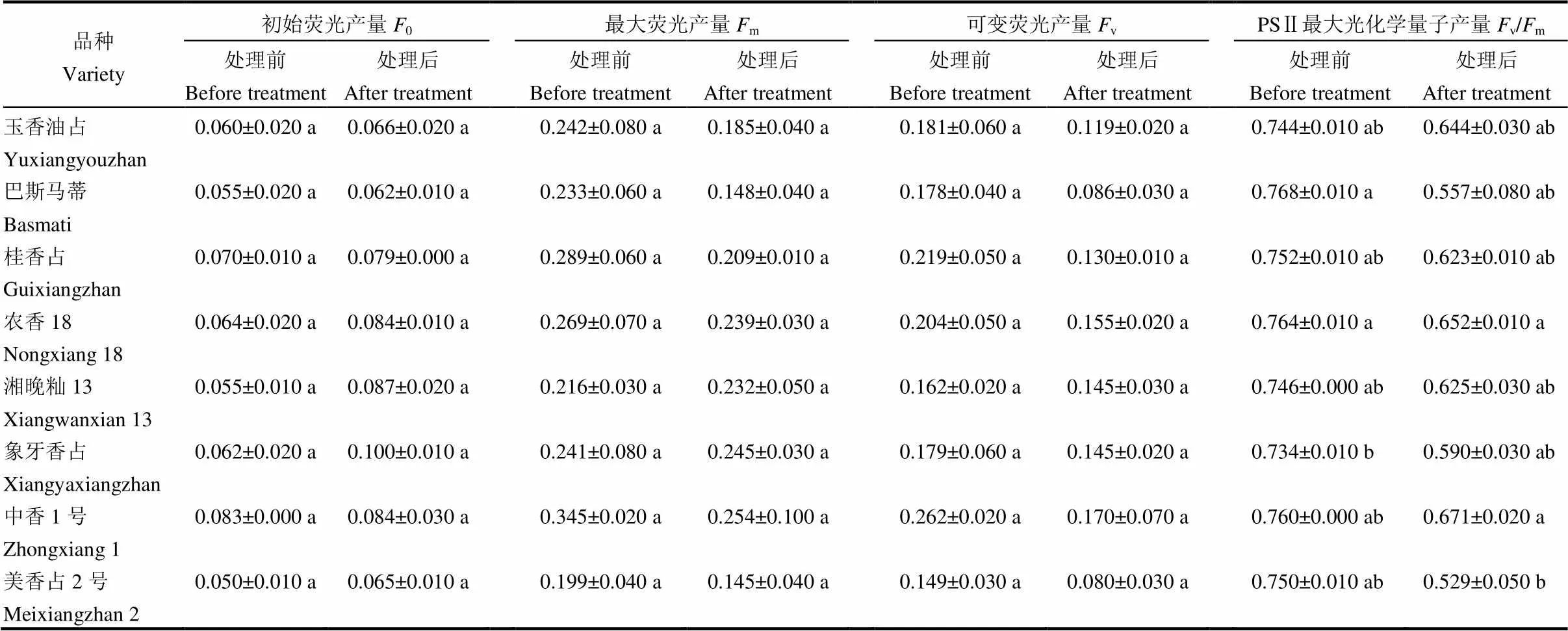
表3 高温胁迫对秧苗叶绿素荧光特性的影响
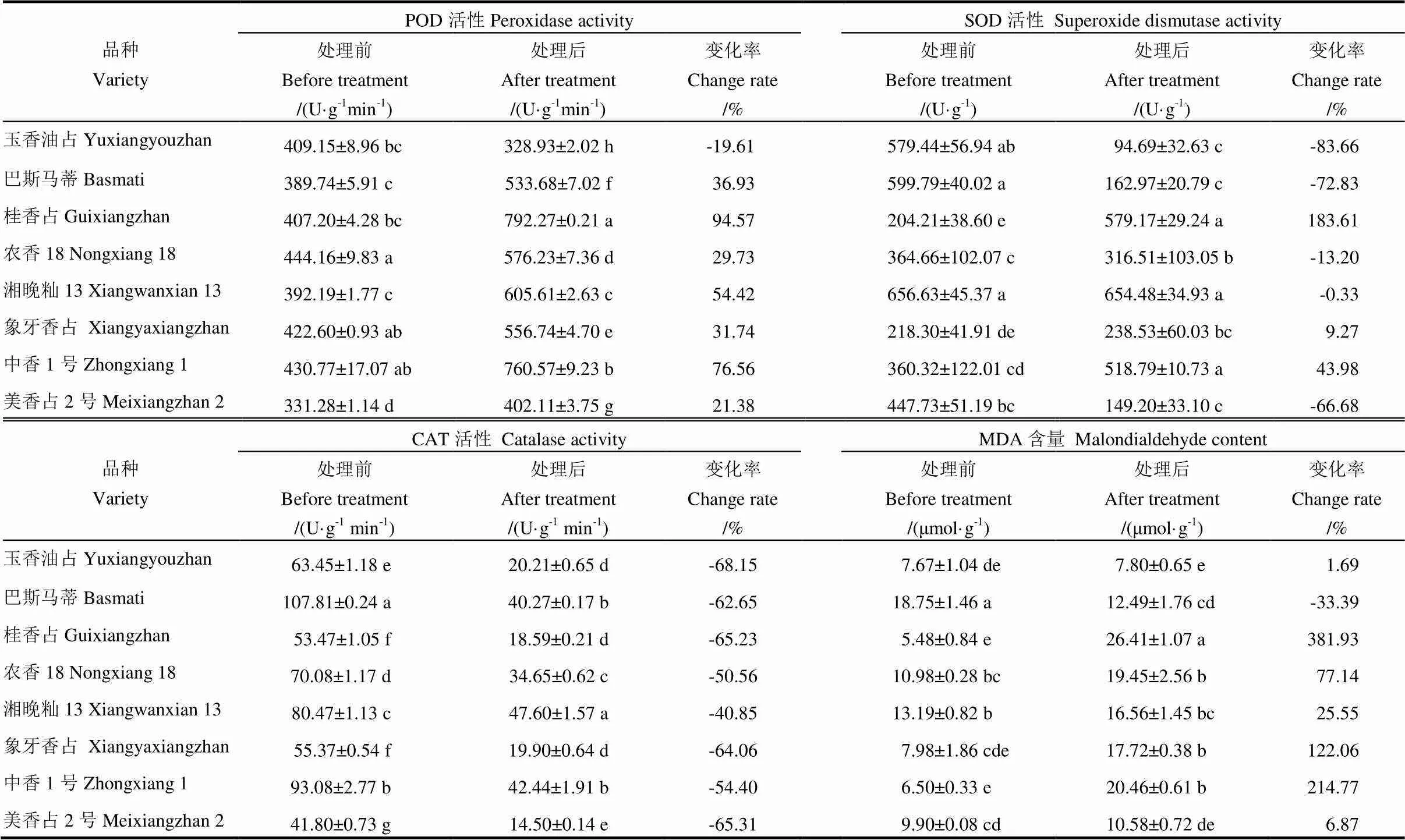
表4 高温胁迫对秧苗抗氧化酶活性以及丙二醛含量的影响
3 讨论与结论
高温胁迫能对作物生长和发育造成不可逆转的影响[16]。有研究表明,水稻在营养生长期遇35℃高温,地上部和地下部的生长受到抑制,株高增加缓慢[17]。本研究结果显示,高温处理后,不同水稻品种的株高和单株鲜质量的增长率存在明显差异甚至出现负增长,单株干质量都有所增加但相互之间有明显差异,这表明高温胁迫降低了秧苗的物质积累速度,阻碍株高的增加甚至使株高下降,降低成活率。其中,巴斯马蒂的株高增幅、单株鲜质量增幅、单株干质量增幅和株成活率均比玉香油占高,这表明巴斯马蒂的耐热性比玉香油占更强。湘晚籼13各方面均次于玉香油占,故其耐热性亦次之。桂香占、中香1号在高温处理后的成活率只有4.33%和1.33%,在8个品种中是最低的,表明两者的耐高温能力均较差。
高温处理会导致植物光合作用受阻,净光合速率和气孔导度降低,胞间CO2浓度上升,这与张桂莲等[18]的研究结果相符。气孔导度影响秧苗的光合作用和蒸腾作用。当高温处理后,秧苗的气孔导度下降,因而净光合速率和蒸腾速率都下降,光合作用受阻导致秧苗不能高效利用细胞间的二氧化碳,导致胞间二氧化碳浓度上升。综合净光合速率、气孔导率、蒸腾作用和细胞间二氧化碳的浓度等因素来看,玉香油占在光合速率以及气体交换方面受到高温胁迫的影响是最小的,其他品种均次于玉香油占。
PSⅡ最大光化学量子产量(v/m)可反映PSⅡ反应中心的光能转化效率,是叶绿素荧光特性中用来表示环境胁迫的指标之一[19]。高温胁迫均能导致8个香稻品种的v/m值降低,说明高温均导致PSⅡ潜在活性中心受损,光合作用的原初反应受到抑制,这与杨卫丽等[20]的研究结论相符。然而,8个香稻品种v/m值的变幅无明显差异,说明8个香稻品种在叶绿素荧光特性方面对高温的胁迫的响应较为一致。
在抗氧化性能上,高温处理后绝大多数品种秧苗的POD活性升高,SOD活性下降,MDA含量积累增多,这与曹云英等[21]的研究结果相符。不同的是,曹云英的试验喷施了油菜素内酯,而本研究没有施用油菜素内酯,这表明香稻本身具有的耐热性与水稻通过喷施油菜素内酯增强耐热性的响应机制是一致的。郑小林等研究表明高温胁迫下,POD活性升高能提高水稻幼苗抗膜脂过氧化的能力,是一种适应高温胁迫的生理反应[22]。POD、SOD、CAT均是抗氧化酶,可维持细胞活性,延缓衰老。每种酶都有其发挥作用最适宜的温度范围,高温(40℃)影响CAT的活性,故8个品种的CAT活性均下降。除玉香油占外,其余7个品种的POD活性均增加,有些品种秧苗的SOD活性增强而有些却减弱,这表明秧苗面对逆境能产生抗逆性,并且由于各品种之间的复杂性和差异性而导致它们的作用机制不一样。除玉香油占外,其余7个香稻品种的耐热性可通过增强POD活性来实现。桂香占、象牙香占和中香1号的耐热性机制通过增强SOD活性来响应。MDA是膜脂过氧化作用的产物,是反映膜脂过氧化反应强弱的指标[23]。MDA对膜和细胞有毒害性,MDA积累的量越大,表明其受到的伤害越大[24]。MDA的含量变化能间接反映膜系统受损程度和植物的抗逆性。因此,高温处理后,只有巴斯马蒂的MDA含量减少,其余品种的MDA含量均增加,可能原因是巴斯马蒂的耐热性机制通过减少MDA含量来响应。结果表明巴斯马蒂受到高温胁迫的伤害最小,其高温耐性最好,其余品种均受到不同程度的伤害,它们的耐热性均比巴斯马蒂弱。
总的来说,巴斯马蒂的耐高温胁迫能力最强,玉香油占次之,接下来依次为湘晚籼13、农香18、象牙香占、美香占2号、桂香占和中香1号。
[1] 罗昊文, 钟卓君, 聂俊, 唐湘如. 超声波处理对粤晶丝苗生理特性及产量、品质的影响. 中国稻米, 2017, 23(2): 64-67.
Luo H W, Zhong Z J, Nie J, Tang X R. Effects of ultrasound on physiological characters, yield and quality of rice Yuejingsimiao., 2017, 23(2): 64-67(in Chinese)
[2] 张洪程, 龚金龙. 中国水稻种植机械化高产农艺研究现状及发展探讨. 中国农业科学, 2014, 47(07): 1273-1289.
Zhang H C, Gong J L. Research status and development discussion on high-yielding agronomy of mechanized planting rice in China., 2014, 47(7): 1273-1289. (in Chinese with English abstract)
[3] 贾士荣, 袁潜华, 王丰, 姚克敏, 裴新梧, 胡凝, 王志兴, 王旭静, 柳武革, 钱前. 转基因水稻基因飘流研究十年回顾. 中国农业科学, 2014, 47(1): 1-10.
Jia S R, Yuan Q H, Wang F, Yao K M, Pei X W, Hu N, Wang Z X, Wang X J, Liu W G, Qian Q. What we have learnt in ten years’ study of rice transgene flow., 2014, 47(1): 1-10. (in Chinese with English abstract)
[4] 沈永平, 王国亚. IPCC第一工作组第五次评估报告对全球气候变化认知的最新科学要点. 冰川冻土, 2013, 35(5): 1068-1076.
Shen Y P, Wang G Y. Key findings and assessment results of IPCC WGI Fifth assessment report., 2013, 35(5): 1068-1076. (in Chinese)
[5] 张桂莲, 陈立云, 雷东阳, 张顺堂. 水稻耐热性研究进展. 杂交水稻, 2005, 20(1): 1-5.
Zhang G L, Chen L Y, Lei D Y, Zhang S T. Progresses in research on heat tolerance in rice.2005, 20(1): 1-5. (in Chinese with English abstract)
[6] 黄显波, 严寒, 胡建林, 田志宏. 高温胁迫下水稻幼苗几个相关生理指标的变化. 长江大学学报: 自然科学版农学卷, 2008, 5(2): 50-53.
Huang X B, Yan H, Hu J L, Tian Z H. Changes of several related physiological indexes of rice seedlings under high temperature stress.:,, 2008, 5(2): 50-53. (in Chinese with English abstract)
[7] 李仁忠, 刘海英, 李建业, 叶海龙, 余久红, 金志凤. 浙江省早稻高温热害发生规律及防御措施. 浙江气象, 2011, 32(1): 23-26, 45.
Li R Z, Liu H Y, Li J Y, Ye H L, Yu J H, Jin Z F. Occurrence law and prevention measures of high temperature heat damage of early rice in Zhejiang Province., 2011, 32(1): 23-26, 45. (in Chinese)
[8] 熊玉江, 乔伟, 姚付启, 罗文兵, 袁念念, 付浩龙. 气候变化和灌溉方式对稻田需水量的影响. 长江科学院院报, 2018, 35(4): 76-80.
Xiong Y J, Qiao W, Yao F Q, Luo W B, Yuan N N, Fu H L. Effect of climate change and irrigation pattern on water demand of paddy field., 2018, 35(4): 76-80. (in Chinese with English abstract)
[9] 赵庆雷, 吴修, 王瑜, 李曰鹏, 陈博聪, 王佳, 马加清. 扬花期高温条件下施肥量对水稻高位分蘖及产量性状的影响. 山东农业科学, 2016, 48(8): 61-66.
Zhao Q L, Wu X, Wang Y, Li Y P, Chen B C, Wang J, Ma J Q. Effects of high temperature at flowering stage and fertilization on high-node tillering and yield traits of rice.. 2016, 48(8): 61-66. (in Chinese)
[10] 陈蔚. 高温对水稻生育特性的影响及其防御对策. 湖北农业科学, 2010, 49(8): 1817-1818.
Chen W. Effects of high temperature on growth and development characteristics of rice and its preventive measures., 2010, 49(8): 1817-1818. (in Chinese with English abstract)
[11] 段骅, 杨建昌. 高温对水稻的影响及其机制的研究进展. 中国水稻科学, 2012, 26(4): 393-400.
Duan H, Yang J C. Research Advances in the effect of high temperature on rice and its mechanism.. 2012, 26(4): 393-400. (in Chinese with English abstract)
[12] 王小菁, 萧浪涛, 董爱武, 王台, 钱前, 漆小泉, 陈凡, 左建儒, 杨淑华, 顾红雅, 陈之端, 姜里文, 白永飞, 孔宏智, 种康. 2016年中国植物科学若干领域重要研究进展. 植物学报, 2017, 52(4): 394-452.
Wang X J, Xiao L T, Dong A W, Wang T, Qian Q, Qi X Q, Chen F, Zuo J R, Yang S H, Gu H Y, Chen Z D, Jiang L W, Bai Y F, Kong H Z, Zhong K. Research progress in several fields of Chinese plant science in 2016., 2017, 52(4): 394-452. (in Chinese)
[13] Zhang X J, Zhang S Y, Li C Q, Zhang W H. Effects of high temperature stress on growth of stress-tolerant rice seedlings with resistibility.&, 2014, 15(4): 576-578, 584.
[14] 朱雪梅, 柯永培, 邵继荣, 林立金, 杨远祥.高温胁迫对重穗型水稻品种叶片活性氧代谢的影响. 种子, 2005, 24(3): 25-27, 32.
Zhu X M, Ke Y P, Shao J R, Lin L J, Yang Y X. Effects of high temperature stress on activated oxygen metabolism of the heavy panicle type of medium indica hybrid rice., 2005, 24(3): 25-27, 32. (in Chinese with English abstract)
[15] 李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000.
Li H S. Principles and Techniques of Plant Physiology and Biochemistry Experiments. Beijing: Higher Education Press, 2000. (in Chinese)
[16] Wahid A, Gelani S, Ashraf M, Foolad M R. Heat tolerance in plants: An overview., 2007, 61(3): 199-223.
[17] 杨纯明, Heilman L J. 谢国禄译. 短期高温对水稻生长发育和产量的影响. 国外作物育种, 1994, 13(2): 4-5.
Yang C M, Heilman L J. Xie G L Trans. Effects of short-term high temperature on rice growth and yield., 1994, 13(2): 4-5. (in Chinese)
[18] 张桂莲, 陈立云, 张顺堂, 刘国华, 唐文邦, 贺治洲, 王明. 抽穗开花期高温对水稻剑叶理化特性的影响. 中国农业科学, 2007, 40(7): 1345-1352.
Zhang G L, Chen L Y, Zhang S T, Liu G H, Tang W B, He Z Z, Wang M. Effects of high temperature on physiological and biochemical characteristics in flag leaf of rice during heading and flowering period., 2007, 40(7): 1345-1352. (in Chinese with English abstract)
[19] Barnabás B, Jäger K, Fehér A. The effect of drought and heat stress on reproductive processes in cereals.&, 2008, 31(1): 11-38.
[20] 杨卫丽, 黄福灯, 曹珍珍, 雷炳婷, 胡东维, 程方民. 高温胁迫对水稻光合PSⅡ系统伤害及其与叶绿体D1蛋白间关系. 作物学报, 2013, 39(6): 1060-1068.
Yang W L, Huang F D, Cao Z Z, Lei B T, Hu D W, Cheng F M. Effects of high temperature stress on PSⅡ function and its relation to D1 protein in chloroplast thylakoid in rice flag leaves., 2013, 39(6): 1060-1068. (in Chinese with English abstract)
[21] 曹云英, 赵华. 高温胁迫下油菜素内酯对水稻幼苗的保护作用. 中国水稻科学, 2007, 21(5): 525-529.
Cao Y Y, Zhao H. Protective roles of brassinolide in rice seedlings under heat stress., 2007, 21(5): 525-529. (in Chinese with English abstract)
[22] 郑小林, 董任瑞. 水稻热激反应的研究: Ⅲ.高温对水稻幼苗叶片过氧化物酶的影响. 湖南农业大学学报, 1998, 24(6): 13-16.
Zheng X L, Dong R R.A Study on the response of rice to heat shock: Ⅲ. High temperature’s effect on peroxidase in seedling leaves of rice., 1998, 24(06): 13-16. (in Chinese with English abstract)
[23] 陈贵, 胡文玉, 谢甫绨, 张立军. 提取植物体内MDA的溶剂及MDA作为衰老指标的探讨. 植物生理学通讯, 1991, 27(1): 44-46.
Chen G, Hu W Y, Xie P T, Zhang L J. Solvent for extracting malondialdehyde in plant as an index of senescence., 1997, 27(1): 44-46. (in Chinese)
[24] 陈少裕. 膜脂过氧化对植物细胞的伤害. 植物生理学通讯, 1991, 27(2): 84-90.
Chen S Y. Membrane-lipid peroxidation and plant stress., 1991, 27(2): 84-90. (in Chinese with English abstract)
Morphophysiological Responses of Different Scented Rice Varieties to High Temperature at Seedling Stage
FENG Huoyi1, JIANG Haolin1, WANG Meng1, TANG Xiangru1,2,3, DUAN Meiyang1,2,3, PAN Shenggang1,2,3, TIAN Hua1,2,3, WANG Shuli1,2,3, MO Zhaowen1,2,3,*
(College of Agriculture,,,;Scientific bserving and Experimental Station of Crop Cultivation in South China,,..,,;Technology Innovation Center of Fragrant Long Grain Rice in Guangdong,,;Corresponding author,:.)
【Objective】The purpose of this research is to study the morphophysiological responses of different scented rice varieties to high temperature at seedling stage and to screen out heat-resistant varieties.【Method】With the super inbred fragrant rice Yuxiangyouzhan as control, a pot experiment was conducted in a light incubator, to reveal morphophysiological responses of seven inbred fragrant rice varieties (Basmati, Guixiangzhan, Nongxiang 18, Xiangwanxian 13, Xiangyaxiangzhan, Zhongxiang 1, Meixiangzhan 2) to high temperature stress. The morphophysiological indexes of the eight scented rice varieties were determined before and after high temperature treatment.【Result】Compared with Yuxiangyouzhan, the plant height, fresh weight, dry weight increased with a langer rate, the survival rate and POD activity in Basmati were higher under high temperature treatment. The malondialdehyde content was decreased by 33.39% for Basmati under high temperature treatment. It indicated that Basmati had higher tolerance to high temperature stress. The dry weight growth rate per plant of Xiangwanxian 13 under high temperature stress was equal to that of Yuxiangyouzhan with lower survival rate than Yuxiangyouzhan, and the change rate of intercellular CO2 concentration was lower after high temperature treatment, the SOD activity and the change rate of malondialdehyde content was the lowest among the eight varieties, indicating that its ability to withstand high temperature stress was slightly lower than that of Yuxiangyouzhan. The survival rates of Nongxiang18, Xiangyaxiangzhan and Meixiangzhan 2 were 13.33%, 12.67% and 16.00%, respectively. The survival rates of Guixiangzhan (4.33%) and Zhongxiang 1 (1.33%) under high temperature stress were the lowest among the eight varieties, indicating their weak high temperature tolerance.【Conclusion】In general, the heat resistance of the eight varieties from strong to weak was Basmati>Yuxiangyouzhan>Xiangwanxian 13>Nongxiang 18>Xiangyaxiangzhan>Meixiangzhan>Guixiangzhan>Zhongxiang 1.
scented rice; high temperature resistance; morphophysiology; seedling
10.16819/j.1001-7216.2019.8022
Q948.112+.2; S511.01
A
1001-7216(2019)01-0068-07
2018-03-08;
2018-05-06。
国家自然科学基金资助项目(31601244,31271646);广州市科技计划资助项目(201707010413)。
猜你喜欢
农业与技术(2024年7期)2024-04-19 00:54:46
农业科技通讯(2023年1期)2023-02-12 07:09:58
福建稻麦科技(2022年3期)2022-09-14 02:17:10
青年文学家(2022年24期)2022-07-04 12:36:01
鸭绿江(2021年35期)2021-04-19 12:24:08
东坡赤壁诗词(2020年3期)2020-07-04 02:50:05
风流一代·经典文摘(2018年5期)2018-05-16 12:05:08
小小说月刊(2017年6期)2017-05-16 17:30:47
疯狂英语·初中天地(2017年9期)2017-04-15 04:30:01
热带农业科学(2015年8期)2015-09-10 12:54:53
