
齐穗至乳熟期不同温度对水稻南粳9108籽粒激素含量、淀粉积累及其合成关键酶活性的影响
2019-01-23 07:53成臣曾勇军程慧煌谭雪明商庆银曾研华石庆华
中国水稻科学 2019年1期
成臣 曾勇军 程慧煌 谭雪明 商庆银 曾研华 石庆华
齐穗至乳熟期不同温度对水稻南粳9108籽粒激素含量、淀粉积累及其合成关键酶活性的影响
成臣 曾勇军*程慧煌 谭雪明 商庆银 曾研华 石庆华
(作物生理生态与遗传育种教育部重点实验室/江西省作物生理生态与遗传育种重点实验室/江西农业大学 双季稻现代化生产协同创新中心/南方粮油作物协同创新中心,南昌 330045;*通讯联系人,E-mail: zengyj2002@163.com)
【目的】为了明确灌浆结实前中期不同温度对晚粳稻籽粒灌浆、总淀粉及其组分含量的影响特征及其机理,【方法】以优质食味粳稻南粳9108为材料,利用植物培养箱控温,于水稻灌浆结实期设置高温(HT,日均温度31℃)、中温(MT,日均温度26℃)和低温(LT,日均温度21℃)三个处理,以自然温度为对照(CK),研究灌浆结实前中期持续高低温对晚粳稻籽粒灌浆、内源激素含量、淀粉及其及合成关键酶活性的影响。【结果】随温度上升(21℃~31℃),晚粳稻结实率和千粒重降低;灌浆前中期籽粒总淀粉积累速率加快,但持续时间短,总含量降低,而直链淀粉含量及其所占比例在整个灌浆期均呈降低趋势;灌浆前中期脱落酸(ABA)含量显著增加,赤霉素(GA3)含量降低,玉米素核苷(ZR)含量呈“先增先降”趋势;MT处理有利于提高灌浆结实期籽粒生长素(IAA)含量,HT和LT处理均降低籽粒IAA含量,LT处理下,IAA含量先降后升,而HT处理籽粒IAA含量一直处于较低水平;籽粒可溶性淀粉合成酶(SSS)活性与ZR变化规律相似,也呈“先增先降”趋势,但二者在灌浆结实期并不同步;淀粉分支酶(SBE)活性增加而淀粉脱分支酶(DBE)活性降低。【结论】不同温度显著影响晚粳稻籽粒内源激素含量和淀粉合成关键酶活性的变化,进而影响晚粳稻籽粒灌浆、总淀粉及其组分的积累。
晚粳稻;温度;籽粒;内源激素;淀粉合成
随着我国经济的快速发展和人民生活水平的不断提高,兼顾水稻产量的同时加大发展优质稻米的生产模式是我国粮食生产的重要走向,并且也是目前缓解我国粮食质量与数量间矛盾问题的有效措施之一[1-2]。气候生态条件是影响水稻产量和品质的首要因子,其中,灌浆结实期是决定水稻高产和优质的关键阶段[3]。若灌浆结实期发生高、低温胁迫,籽粒灌浆过程中生理生化的差异显著,从而直接影响水稻籽粒生长发育。
温度胁迫对水稻籽粒形成的生理生化机制已有大量研究。前人主要从淀粉形成[4]、氮代谢[5]、籽粒灌浆[6]、胚乳淀粉结构[7]等角度进行较为深入的探讨。而有关温度对水稻灌浆结实与籽粒中内源激素含量及淀粉合成关键酶活性变化的关系也有一些报道。李木英等[8]认为,高温下通过淀粉合成关键酶可以调控淀粉积累速率及籽粒灌浆,且不同水稻品种间胚乳淀粉合成酶对高温的反应也存在差异。王丰等[9]认为,在灌浆初期高温显著降低水稻籽粒IAA、ZR和GA3含量,特别是籽粒ABA含量剧增,是导致其早期灌浆速率增加、灌浆持续时间缩短及灌浆进程提前的重要原因。但上述均是对灌浆结实期短期不同温度下有关籽粒灌浆、淀粉形成与其内源激素含量变化关系的研究,而对长期不同温度胁迫的相关研究较少。
南方双季稻区是我国最重要的粮食主产区之一,在保障我国粮食安全中起着不可替代的作用[10-11]。近年来,南方双季稻区“籼改粳”得到迅速发展,“晚粳”种植模式成为当地推广粳稻的主要模式[12]。与当地晚籼稻相比,晚粳稻的产量及品质均表现较好[13]。然而不同纬度的生态气候条件存在较大差异,晚粳稻的推广区域也存在一定限制,其中灌浆结实期日均温的差异是导致南方晚粳稻产量和品质形成差异的主要原因。因此,为了模拟晚粳稻在不同纬度灌浆结实期籽粒发育过程中的生理变化特征,以优质食味水稻南粳9108为材料,研究灌浆结实前中期(长期)不同日均温度处理下晚粳稻籽粒内源激素、淀粉合成及其关键酶活性的变化规律,以期为南方晚粳稻抗逆栽培和高产优质生产提供理论依据。
1 材料与方法
1.1 试验地点与供试品种
试验于2017年晚季在江西省上高县泗溪镇曾家村(115°6'52″E,28°16'42″N)开展。采用盆栽模拟方式开展,盆栽内土壤取自当地农田0~20 cm耕层,土壤基础肥力如下:有机碳含量23.1 g/kg,全氮2.37 g/kg,速效钾80.7 mg/kg,速效磷23.4 mg/kg,pH 5.22。供试品种为优质食味粳稻南粳9108。
1.2 试验设计
根据江西双季稻地区近30年来9-10月份的气候特征以及上高近5年晚稻灌浆结实期温度的变化规律,预计2017年上高生态点在晚稻灌浆前中期日平均温度为26℃±4℃,即最高温与最低温相差8℃。因此,本研究设置三种温度处理,高温(HT,日平均气温31℃,27℃~35℃)、中温(MT,日均温26℃,22℃~30℃)和低温(LT,21℃,17℃~25℃)。晚粳稻齐穗后,将盆栽从田间移入室内植物光照培养箱[日本三洋(松下),型号为MLR-352-PC],其有效容积为294 L(宽、深和高分别为0.76 m、0.70 m和1.84 m)。温度处理持续17 d,湿度设置为80%,光照最强档位,其光合有效光子通量约为1 000 μmol/(m2∙s),光照时间为6:00-18:00。以水稻始终在室外正常生长为对照,未对其温湿光等自然气候条件进行任何人为干预。每个处理3个重复,每重复5盆,4个处理总共60盆。具体各处理气温日变化如图1-A所示。
试验所用盆钵的高度与直径为30 cm×30 cm,每盆栽插1穴,每穴4根苗。水稻于6月27日播种,7月20日移栽,9月6日齐穗,当天将盆栽移入不同温度处理的人工气候箱,9月24日移至室外,10月15日左右成熟。对照在9月6日至9月23日期间的日平均温度为26.6℃(31.9℃~22.5℃),与MT处理所设置温度特征基本一致(图1-B)。
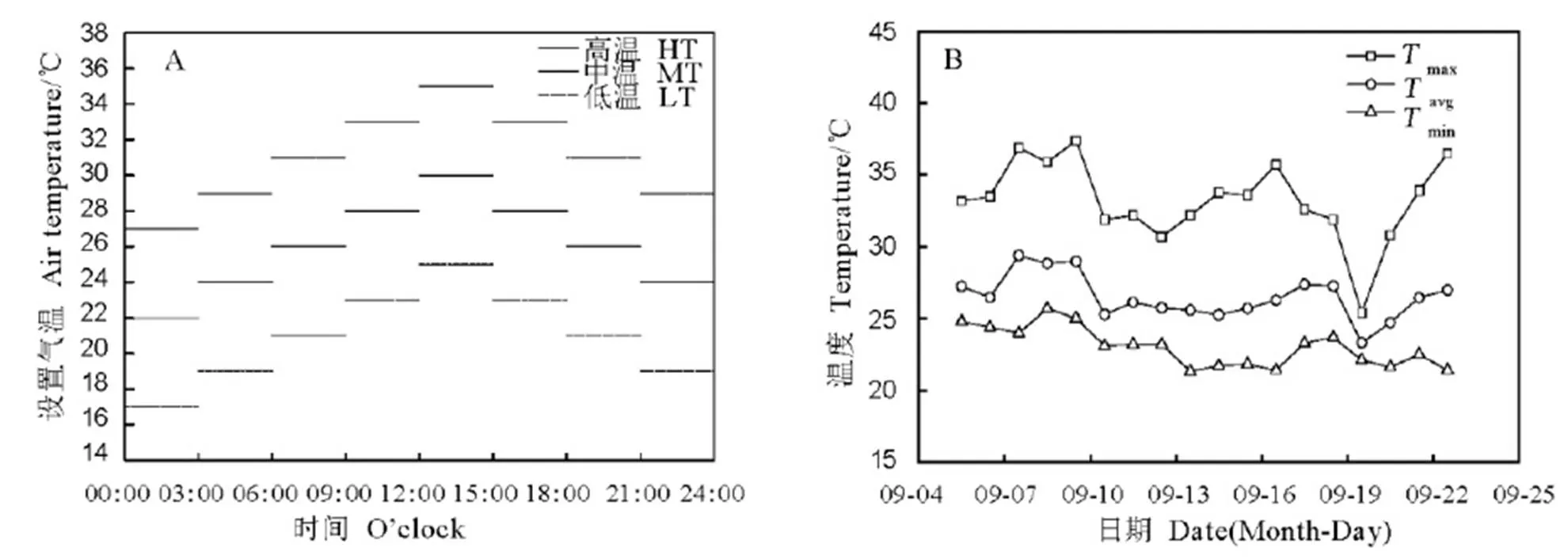
max、avg和min分别指室外一天最高气温、平均气温和最低气温。CK代表与MT处理日平均温度较为一致的对照处理;LT代表低温21℃(17℃~25℃);MT处理代表中温26℃(22℃~30℃);HT处理代表高温31℃(27℃~35℃)。
max,avgandminrefer to the maximum temperature, average temperature and minimum temperature of a day, respectively. CK, A control treatment consistent with MT treatment in daily average temperature; LT, Low temperature of 21℃(17℃~25℃); MT treatment, Medium temperature of 26℃ (22℃~30℃); HT treatment , High temperature of 31℃(27℃~35℃). The same as below.
图1 不同处理的温度日变化及温度处理期间(9月6日至9月23日)室外气温变化
Fig.1. Daily change patterns of temperature in different treatments and outdoor temperature changes during experiment (from 6th to 23rd, September).
1.3 样品采集与测定方法
1.3.1 样品采集
于水稻抽穗期,对同日抽穗的稻穗挂牌标记,分别于晚粳稻齐穗后10、16、23和30 d的上午9:00-10:00对各处理挂有标签牌的稻穗进行取样,摘取穗中部灌浆籽粒,并立即将籽粒放入液氮中处理1 min,随后放入并保存于−80℃的超低温冰箱中,用于淀粉及内源激素含量等指标的测定。
1.3.2 样品测定
籽粒淀粉合成关键酶、总淀粉含量及其组分等测定均采用购买的相关试剂盒(苏州科铭生物技术有限公司生产)并按照相应测定方法进行[8,14]。籽粒内源激素ABA、ZR、GA3和IAA含量测定采用高效液相色谱法(HPLC)[15]。每份样品取籽粒30粒左右,加入80%预冷甲醇置弱光下冰浴研磨至匀浆,在4℃(低温)下过夜浸提16 h,8000×下冷冻离心10 min取出上清液,残渣用80%预冷甲醇重复浸提2 h。将石油醚用于色素除杂及调节溶液的pH,用有机相萃取进一步除杂提纯,随后进行上机测试。通过不同的流动相配比、柱温及流速等调节,将目标峰与杂质峰区分,从而得到各测定指标的峰面积及含量。每样品测定3次重复,取平均值。IAA、ZR、GA3和ABA的回收率分别为87.1%、89.3%、95.4%和96.8%。
1.4 数据分析
采用Microsoft Excel 2010和DPS 7.05软件分析和处理数据;Origin 9.0软件制图;LSD 法进行差异显著性检验(<0.05)。
2 结果与分析
2.1 晚粳稻结实率及千粒重
灌浆结实前中期不同温度对晚粳稻结实率和千粒重均有显著的影响(表1)。方差分析表明,LT处理结实率和千粒重均显著高于MT和HT处理,而MT处理与CK间在结实率和千粒重上均无显著差异。与LT处理相比,MT和HT处理结实率分别降低3.8%和27.2%,千粒重分别降低4.2%和14.9%,而与MT处理相比,CK处理结实率与千粒重略有增加。表明随着灌浆结实前中期温度上升,晚粳稻结实率及千粒重均呈降低趋势,其中日平均温度在26℃~31℃之间二者的降幅较大。
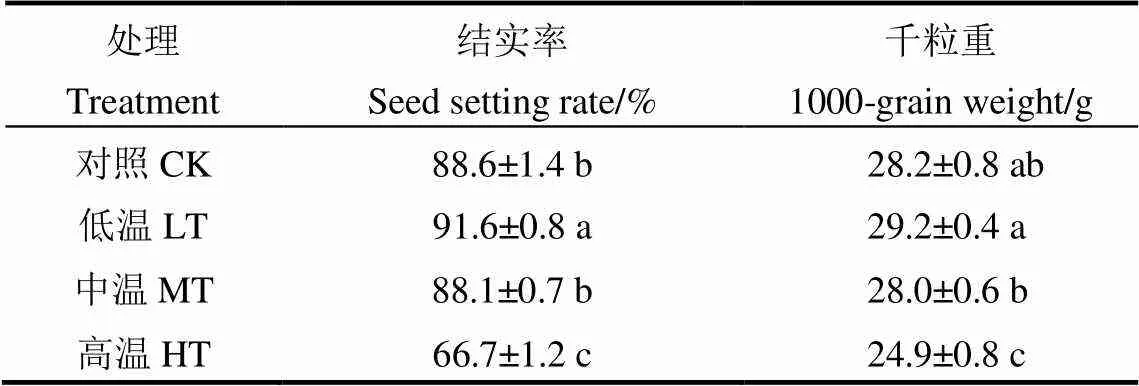
表1 灌浆结实前中期温度对晚粳稻产量构成因素的影响
表中数据均采用平均值±标准差表示(=4);表中同列数据后跟不同小写字母表示差异达5%显著水平(﹤0.05)。下同。
Data are shown as mean±SD,= 4. Different lowercase letters in the same column indicate significant difference at 5% level. The same as below.
2.2 籽粒总淀粉及其组分含量
2.2.1 总淀粉含量
HT处理总淀粉含量在灌浆前中期积累速率较快,至齐穗16 d后籽粒总淀粉含量增速放缓,LT处理在籽粒灌浆前中期总淀粉含量积累速率却较为缓慢,但持续时间较长,至齐穗后23 d总淀粉含量基本稳定,而MT和CK介于二者之间(图2-A)。籽粒总淀粉含量在齐穗后10 d和16 d表现为HT>MT(CK)>LT,而在齐穗后第23 d和30 d却呈相反的变化规律。与MT处理相比,LT处理在齐穗后30 d总淀粉含量增加1.8%,而HT和CK处理分别降低6.8%和2.7%,其中HT处理与MT处理间达差异显著水平。这说明随温度升高,灌浆前期总淀粉含量积累速率加快,积累时间缩短,灌浆进程提前,最终总含量降低。
2.2.2 直链淀粉含量及其占总淀粉含量比值
由图2-B可知,各处理直链淀粉含量在灌浆前中期积累速率均较快,中后期积累速率较为缓慢,至齐穗后16 d基本达到稳定。在籽粒灌浆整个过程中,直链淀粉含量均表现为LT>MT(CK)>HT,至齐穗后30 d,与MT处理相比,LT和CK处理直链淀粉含量分别增加16.1%和0.5%,而HT处理降低10.8%,其中LT处理与MT处理间差异达到显著水平。各处理直链淀粉含量占总淀粉含量的比值表现为先增后降或逐渐增加的趋势,至齐穗后23 d基本变化不大(图2-C)。其比值在籽粒灌浆整个过程中均表现为LT>MT(CK)>HT;至齐穗后30 d,与MT处理相比,LT和CK处理直链淀粉含量所占比值分别增加14.0%和3.4%,而HT处理降低4.4%。说明随温度升高,直链淀粉含量及其所占比例在整个灌浆期均呈降低趋势。
2.2.3 支链淀粉含量及其占总淀粉含量比值
各处理支链淀粉含量与总淀粉含量的变异规律基本一致(图2-D)。支链淀粉含量在籽粒灌浆前中期(齐穗后10 d和16 d)表现为HT>MT(CK)>LT,而在齐穗后23 d和30 d表现为相反的变化趋势。在齐穗后30 d,与MT处理相比,LT处理支链淀粉含量增加0.6%,而HT和CK处理分别显著降低6.4%和3.0%。在整个籽粒灌浆过程中,各处理支链淀粉含量占总淀粉含量的比值表现为先降后增或逐渐降低的趋势,至齐穗后23 d基本稳定;不同处理支链淀粉含量与总淀粉含量的比值在灌浆期均表现为HT>MT(CK)>LT,与直链淀粉含量所占比值的变化规律基本相反(图2-E)。至齐穗后30d,与MT处理相比,LT和CK处理支链淀粉含量所占比例分别降低1.2%和0.3%,而HT处理增加0.4%。
2.3 内源激素含量
2.3.1 生长素(IAA)含量
由图3-A可知,MT和CK处理IAA含量变化规律相似,在齐穗后10-30 d均高于HT处理。而LT处理IAA含量在齐穗后10-16 d下降,随后逐渐增加;LT处理IAA含量在齐穗后第10 d和30 d均略高于其他处理,而在灌浆中期处于较低水平。表明在灌浆结实期,LT处理籽粒IAA含量先降后升,而HT处理籽粒IAA含量一直处于较低水平。因此,MT处理有利于提高灌浆结实期籽粒IAA含量,高温或低温均有效降低籽粒IAA含量。
2.3.2 玉米素核苷(ZR)含量
由图3-B可知,HT、MT和CK处理ZR含量均呈单峰型变化特征,其中HT处理齐穗后16d达到峰值且其峰值高于其他处理,而MT和CK处理ZR含量变化趋势基本一致,在齐穗后23 d达到峰值,而LT处理ZR含量在齐穗后10-30 d内一直呈增加趋势。说明高温促进籽粒ZR含量快速增加并提前达到峰值,温度越高,达到峰值时间越短,下降时间也提前,呈“先增先降”变化规律。
2.3.3 赤霉素(GA3)含量
由图3-C可知,各处理籽粒GA3含量在齐穗后10 d至30 d均逐渐降低的趋势,但下降幅度存在差异。各处理在整个灌浆过程中GA3含量均表现为LT>MT>HT,而CK处理GA3含量降幅最大,其值在灌浆前中期高于LT处理,灌浆中后期与MT处理无显著性差异。表明在籽粒灌浆过程中HT处理降低GA3含量,LT处理有利于提高GA3含量,CK处理与MT处理在灌浆前中期存在差异。
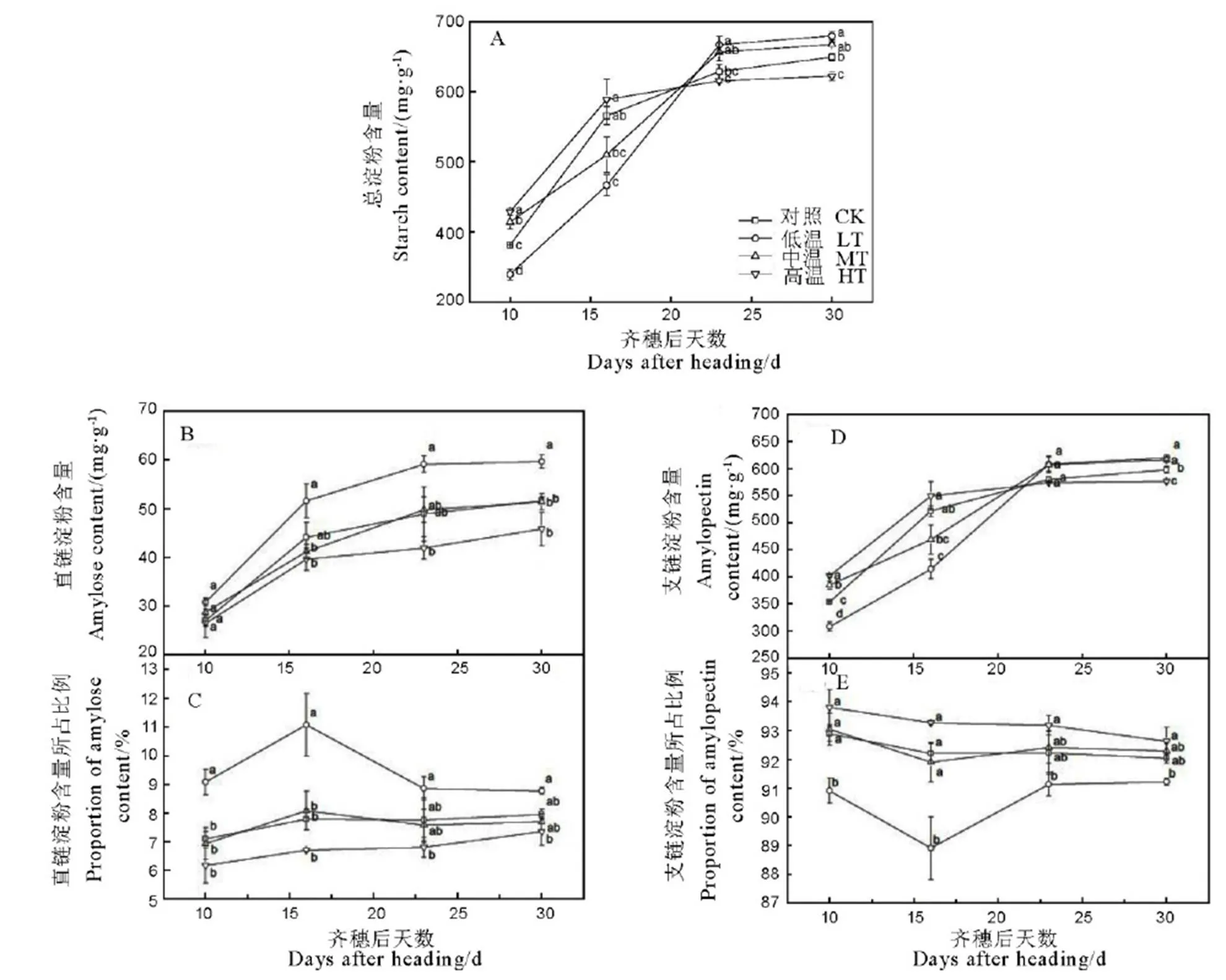
图2 水稻籽粒总淀粉及其组分含量的变化
Fig. 2. Changes in grain starch contents and its components.
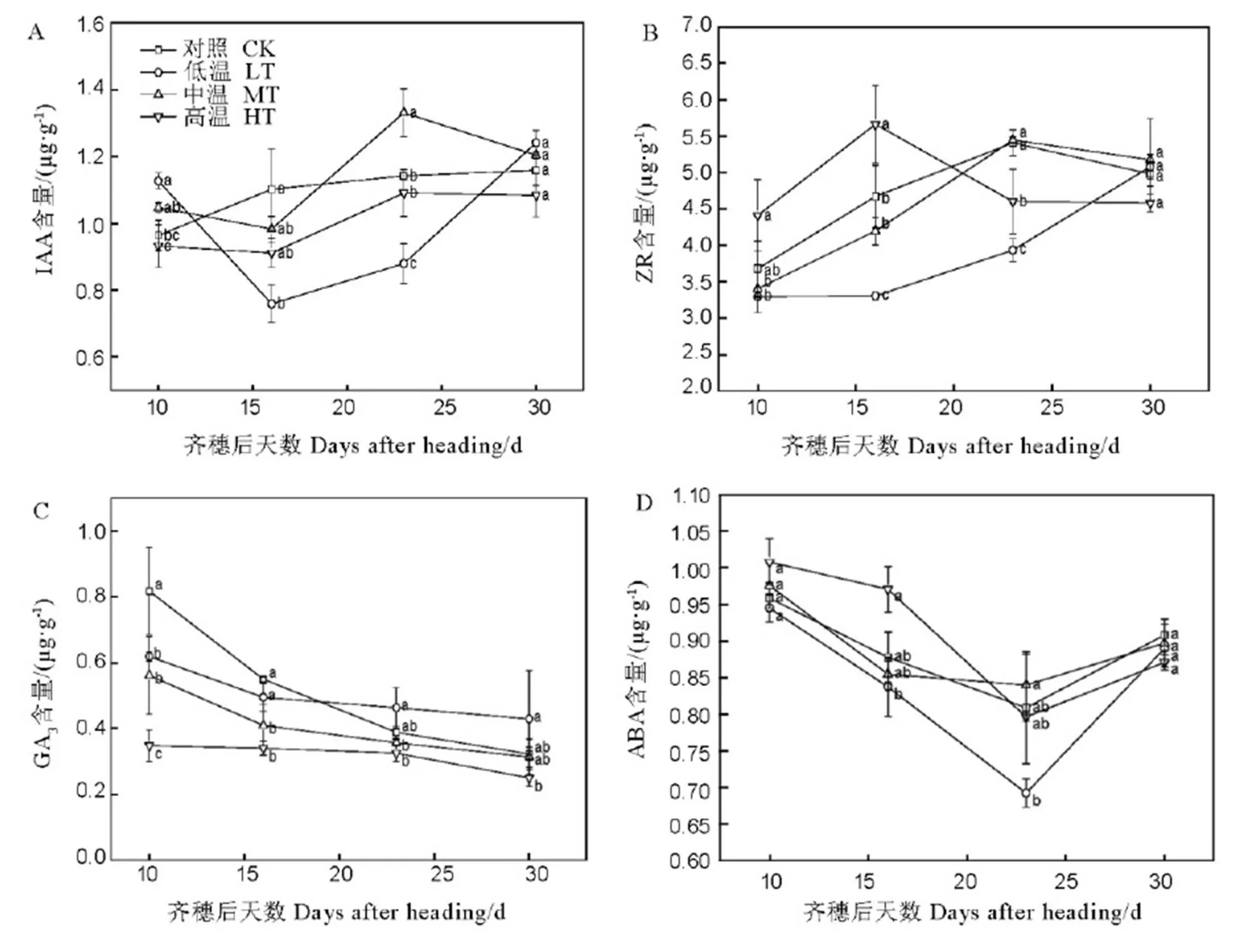
图3 不同温度处理下晚粳稻籽粒灌浆过程中内源激素含量的变化
Fig. 3. Changes in grain endogenous hormone contents in developing grains of laterice under different temperature.
2.3.4 脱落酸(ABA)含量
各处理籽粒ABA含量在齐穗后10 d开始降低,至齐穗后23 d达到最低值,随后略有增加(图3-D)。在齐穗后10 d和16 d籽粒ABA含量表现为HT>MT(CK)>LT;在齐穗后23 d和30 d, HT、MT和CK处理三者ABA含量在齐穗后23 d均高于LT处理,至齐穗后30 d各处理ABA含量均无显著差异。表明高温胁迫能促进灌浆前中期籽粒ABA含量,对灌浆后期影响不大。
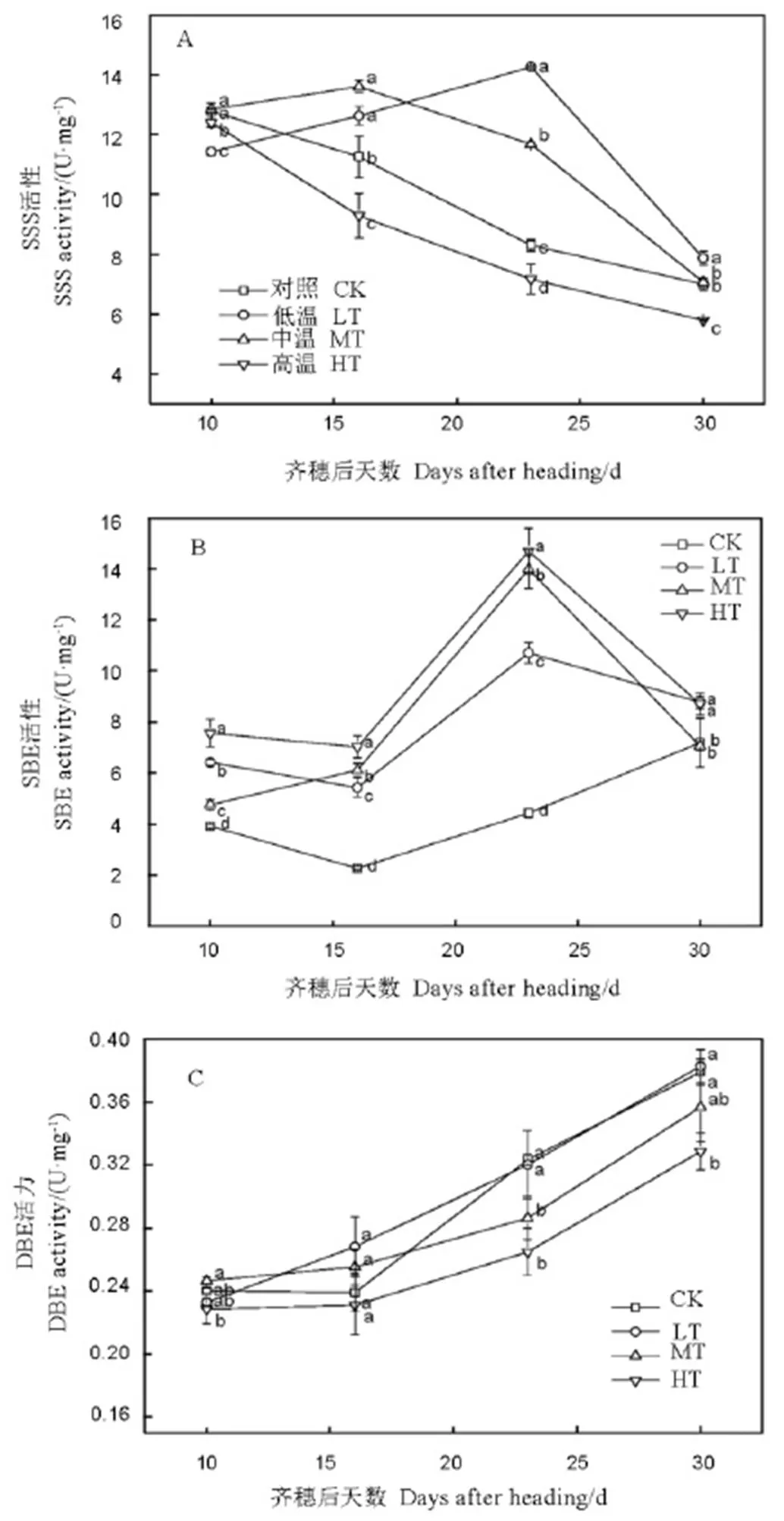
图4 水稻籽粒可溶性淀粉合成酶、可溶性淀粉分支酶及淀粉脱分支酶活性的变化
Fig. 4. Changes in activities of grain soluble starch synthase (SSS), starch branching enzyme (SBE) and starch debranching enzyme (DBE).
2.4 淀粉合成关键酶活性
2.4.1 可溶性淀粉合成酶(SSS)活性
由图4-A可知,HT处理籽粒SSS活性在齐穗10 d之后处于一直降低的趋势,而MT处理和LT处理均呈现先增后降单峰型变化规律,其中二者分别在齐穗后16 d和23 d达到峰值。HT和MT处理SSS活性在齐穗后10 d显著高于LT处理,在齐穗后23-30d各处理SSS活性均表现为LT>MT>HT。而CK处理与MT处理SSS活性的变化规律相似,但在整个灌浆期均低于MT处理。表明随着灌浆结实前中期温度升高,籽粒SSS活性持续增加时间缩短,降低时间提前,在整个灌浆期基本处于较低水平,呈现“先增先降”的变化规律。
2.4.2 淀粉分支酶(SBE,又称Q酶)活性
各处理籽粒在灌浆前期(齐穗后10-16 d)SBE活性均较低且变幅不大,随后快速增加至齐穗后23 d又大幅下降,而CK处理SBE活性在齐穗后16-30 d逐渐增加(图4-B)。在灌浆中期SBE活性表现为HT> MT>LT,而CK处理的SBE活性均处较低水平。说明,随着灌浆结实前中期温度升高,能增加灌浆中期籽粒SBE活性,对灌浆后期影响不大。
2.4.3 淀粉脱分支酶(DBE,又称R酶)活性的变化
各处理籽粒DBE活性在齐穗后10-30 d均呈逐渐增加的趋势,其中,各处理籽粒DBE活性在齐穗后10 d较低且差异不大,而在齐穗后16-30 d增幅较大,处理间差异显著(图4-C)。在灌浆中后期各处理籽粒DBE活性表现为LT >MT >HT,CK处理籽粒DBE活性变化规律与MT处理总体一致。表明随灌浆结实前中期温度升高,籽粒DBE活性表现降低的趋势。
3 讨论
3.1 不同温度对晚粳稻籽粒直链淀粉和支链淀粉形成的影响
水稻籽粒中直链淀粉含量主要受品种遗传控制,多数学者认为灌浆结实期温度是影响籽粒淀粉组分的重要因素,但前人关于二者之间关系的研究结果存在差异。Jiang等[16]研究认为与低温相比水稻生长在高温条件下其直链淀粉含量较低。Ahmed等[17]报道,高温降低淀粉总含量和改变淀粉组成,寒冷的天气会造成同一品种直链淀粉含量的增加。本研究也得到了类似的结果,随着灌浆前中期胁迫温度的增加,晚粳稻籽粒直链淀粉含量及其所占总淀粉含量比例在整个灌浆期均降低,而支链淀粉含量所占总淀粉含量比例却逐渐增加。究其原因,从SBE活性变化来看,在灌浆中期各处理表现为HT> MT>LT,而在灌浆前、后期差异不大(图4-B);SBE切开来自支链淀粉的直链区或者直链淀粉中α-1,4-葡聚糖直链供体的α-1,4-糖苷键,并催化其所切下的短链与受体链二者间形成α-1,6-糖苷键,因而产生分支,是参与合成支链淀粉的关键酶,也是催化直链淀粉转化为支链淀粉的催化剂[18-19],说明高温下SBE较高的酶活性是增加支链淀粉含量及其比例的重要原因之一。从DBE活性变化来看,在灌浆中后期各处理籽粒DBE活性表现为LT>MT> HT,而在灌浆前期差异不大(图4-C);DBE是对裂解支链淀粉的α-1,6-糖苷键、形成线性葡萄糖链具有专一性的酶,即DBE具有催化支链淀粉产生的还原糖的特性,从而对调整支链淀粉分子的链长方面具有重要的“修饰”作用[20, 21]。说明低温下DBE较高的酶活性是提高直链淀粉含量及其比例重要因素之一。综上所述,随灌浆结实前中期温度增加(21℃~31℃),SBE活性增加及DBE活性降低,二者的共同作用是晚粳稻籽粒直链淀粉含量降低、支链淀粉含量增加的重要原因之一。
吕艳梅等[22]却研究认为齐穗后高温胁迫降低了籽粒淀粉和支链淀粉含量,而使直链淀粉含量增加。而周德翼等[23]指出,稻米直链淀粉含量主要受灌浆结实前期温度控制,高直链淀粉含量品种在高温胁迫下其直链淀粉含量增加,而直链淀粉含量中、低的品种却呈现相反的规律,此外直链淀粉对温度的敏感性与胁迫温度的高低有关联,且品种类型间直链淀粉含量存在显著差异。蔡水文[24]也研究表明,在高温条件下直链淀粉含量较高的品种湘早籼33号和湘早籼24号在整个灌浆结实期阶段其直链淀粉含量均高于适温条件,而高温条件下直链淀粉含量较低品种湘早籼31号于灌浆中后期籽粒直链淀粉含量比适温条件下较低。因此,为了更加全面分析晚粳稻淀粉及其组分含量对温度的响应,有必要对不同直链淀粉含量的粳稻品种进一步深入研究。
3.2 晚粳稻籽粒淀粉合成及其灌浆对不同温度的响应
水稻灌浆结实期的温度是影响籽粒灌浆、稻米总淀粉含量及其组成的重要因素[25, 26]。刘萍等[4]认为籽粒总淀粉含量变化规律与胁迫温度有关,灌浆结实期温度从25℃增加至30℃时有利于籽粒总淀粉积累,而当温度超过30℃时,籽粒总淀粉积累能力降低,这与Macleod 等[27]研究结果较为一致。而杨欢等[28]研究表明,结实期高温胁迫使玉米籽粒淀粉含量在灌浆前期增加,而于灌浆后期较低于常温;高温胁迫使灌浆前中期胚乳细胞中淀粉体充实提前,胚乳细胞中淀粉体积累速率增加,但淀粉持续积累时间减少,中后期淀粉积累能力明显不足。蔡水文[24]也研究表明,与适温条件相比,高温条件下不同水稻类型品种籽粒淀粉含量在齐穗后5-14 d均显著较高,但在齐穗14 d后淀粉累积即趋于停止,而适温条件下灌浆前中期淀粉积累速度较慢,但其齐穗后20 d淀粉含量仍有增加,至成熟期高温处理下淀粉含量显著低于适温处理。本研究得到类似结果,即随着灌浆期胁迫温度增加,晚粳稻籽粒总淀粉累积含量在灌浆前中期增加而在灌浆中后期降低;其中HT和LT处理淀粉积累分别在齐穗后16 d和23 d停止,而MT(CK)处理介于二者之间。说明本研究结果与前人基本一致,但在籽粒淀粉积累持续时间上存在一定的差异,这可能与灌浆期胁迫温度以及品种的差异有关。
水稻籽粒中淀粉占最终糙米干质量的80%~ 90%,籽粒灌浆及粒重增加的本质是淀粉合成与积累的过程[29]。因此,灌浆结实期晚粳稻籽粒灌浆、粒重变化规律应与籽粒总淀粉含量对不同温度的响应较为一致,即随温度上升,籽粒前期灌浆速率增加,但灌浆持续时间缩短、灌浆进程提前,最终籽粒粒重降低、灌浆不充实。本研究结果也表明,与LT处理相比,MT和HT处理结实率分别显著降低3.8%和27.2%,千粒重分别显著降低4.2%和14.9%。说明高温处理是导致“源”不足的重要原因,从而显著降低晚粳稻结实率和千粒重,这与前人研究结果较为一致[30]。
3.3 不同温度对晚粳稻籽粒淀粉合成及其灌浆的作用机理
从籽粒SSS活性变化特征来看,本研究表明,随着灌浆结实前中期温度增加,籽粒SSS活性增强持续时间缩短,降低时间提前,在整个灌浆期基本处于较低水平,呈现“先增先降”的变化规律。SSS是催化淀粉合成的一个关键酶,主要以游离态的形式存在于胚乳淀粉粒中,其催化前体物质ADPG 与淀粉引物(葡聚糖)发生反应,达到将葡萄糖分子有效转移到淀粉引物上的目的,从而使催化淀粉链延长;即籽粒中SSS 活性越强, 其利用ADPG 合成淀粉的能力就越强[31, 32]。因此,HT处理利用ADPG 合成淀粉能力在灌浆期呈逐渐降低且较低的水平,而LT处理利用ADPG 合成淀粉的能力总体表现较强,说明高温不利于籽粒总淀粉的积累。综上,灌浆前中期不同温度下通过改变籽粒SSS活性是调控淀粉合成及其灌浆过程的重要途径之一,但通过其他淀粉合成关键酶(如蔗糖合成酶、ADPG焦磷酸化酶等)的调控作用及机理还有待于研究。
从籽粒内源激素来看,前人研究表明[33, 34],植物内源激素对籽粒灌浆结实过程也具有重要的调控作用,主要包括以下两种途径:一方面,植物各种内源激素含量及其之间的协同或拮抗作用,通过调控籽粒中碳水化合物、蔗糖、蛋白质及其各种酶等物质的合成及运输来调控水稻籽粒灌浆进程;另一方面,籽粒内源激素含量及其相互间的关系受生态环境因子(温、光等)的调节影响,最终影响籽粒发育。
水稻籽粒中ZR及IAA含量主要通过调节细胞发育相关酶的活性来实现对籽粒胚乳细胞数目、体积及内容充实物的调控,相关研究表明二者与最大胚乳细胞数目以及胚乳细胞的平均增殖速率均呈显著正相关[9]。本研究表明,MT处理有利于提高灌浆结实期晚粳稻籽粒IAA含量,高温或低温均有效降低晚粳稻籽粒IAA含量,其中LT处理下灌浆结实期籽粒IAA含量先降后升,而HT处理籽粒IAA含量一直处于较低水平。同时高温促进晚粳稻籽粒ZR含量快速增加并提前达到峰值,温度越高,达到峰值时间越短,下降时间也提前,呈“先增先降”趋势。由此说明,灌浆前期高温胁迫下通过快速提高ZR含量来加强胚乳细胞分裂及分化,从而促使晚粳稻籽粒灌浆加快,并通过持续降低IAA含量来减少胚乳细胞的数量,减小体积,抑制籽粒同化物的积累,从而造成高温下籽粒灌浆持续时间短但灌浆速率增加的现象。而低温下ZR含量缓慢增加,胚乳细胞分裂及分化速率逐渐提升,籽粒灌浆速率较为平缓,IAA含量先降后增,胚乳细胞数量和体积增加速率先降低后上升,从而同化物积累速率也呈先降后增的趋势,因此低温处理灌浆时间持续时间长,于灌浆后期依然具有较强的同化物积累能力。这与前人有关不同温度下IAA及ZR的研究结果较为一致[9, 35]。
GA3可能具有激活或诱导产生蛋白水解酶及淀粉酶等活性的特性,而不利于淀粉及蛋白质等物质的合成积累,其与胚乳细胞活跃增殖和籽粒活跃灌浆期呈显著负相关,但对花粉的发育具有促进作用[36]。本研究结果表明,高温处理降低整个籽粒灌浆过程中GA3含量,而低温处理有利于提升GA3含量。说明高温(低温)处理通过降低(增加)晚粳稻籽粒中GA3含量来进一步调节籽粒胚乳细胞的增殖与灌浆进程,增加(降低)晚粳稻籽粒灌浆速率,从而缩短(延长)籽粒灌浆持续时间。曾研华等[36]也研究认为,灌浆前、中期低温处理通过增加甬优17和中浙优1号籽粒的 GA3含量来降低籽粒灌浆速率和延长籽粒灌浆持续期,有利于千粒重的提高。
ABA主要通过调控ATP酶和蔗糖合酶的活性及酸性转化酶,促进光合同化物“源”向籽粒“库”的运输并加强蔗糖在籽粒库的卸载能力;同时还可降低一些水解酶活性,提高淀粉合成酶、ADGP焦磷酸酶和淀粉分支酶活性,加快籽粒淀粉合成速率及灌浆进程[35]。本研究结果表明,温度上升,促进灌浆前中期晚粳稻籽粒ABA含量提升,但对灌浆后期影响不明显。说明高温促进ABA含量增加从而进一步提高晚粳稻灌浆前中期籽粒同化产物的积累及充实速率。这与杨欢等[28]研究结果较为一致,其认为高温胁迫下灌浆前期较高的ABA促进光合同化物向籽粒运转及加快籽粒灌浆速率,但持续高温会加速叶片衰老和降低同化物供应量,使灌浆后期籽粒增重严重受抑。
综上,温度胁迫下各内源激素含量及淀粉合成关键酶活性变化的作用结果与籽粒灌浆结实、总淀粉及其组分含量变化规律吻合,从而再次论证籽粒内源激素含量及淀粉合成关键酶活性的变化是调控籽粒灌浆的重要途径。由本研究结果可以推测,随着纬度增加,晚粳稻灌浆结实期日均温度增加,是导致籽粒千粒重、结实率和总淀粉含量降低以及抽穗至成熟期生育期缩短重要原因之一。水稻籽粒灌浆是一个复杂的调控过程,各内源激素的作用机理及其互作效应对籽粒灌浆形成的影响非常复杂,其协同调控机制及分子机理还不清楚,以及与淀粉合成关键酶之间的作用关系仍需深入探讨;此外,本研究不同温度处理持续时间较长,而双季晚粳稻灌浆结实期遭遇寒露风以及异常高温等天气的次数及程度均呈增加的趋势[37, 38],若晚粳稻遭遇短期高温或低温其内源激素含量及淀粉合成关键酶活性的变化特征及作用结果还有待于进一步研究。
[1] 凌启鸿, 张洪程, 丁艳锋,张益彬. 水稻高产技术的新发展——精确定量栽培. 中国稻米, 2005, 11(1): 3-7.
Ling Q H, Zhang H C, Ding Y F, Zhang Y L. The new development of rice high yield technology: Accurate and quantitative cultivation., 2005, 11(1): 3-7.(in Chinese)
[2] Huang L, Jun Y U, Yang J, Zhang R, Bai Y, Sun C M, Zhuang H Y. Relationships between yield, quality and nitrogen uptake and utilization of organically grown rice varieties., 2016, 26(1): 85-97.
[3] 程方民, 刘正辉, 张嵩午. 稻米品质形成的气候生态条件评价及我国地域分布规律. 生态学报, 2002, 22(5): 636-642.
Cheng F M, Liu Z H, Zhang S W. The evaluation of climatic-ecology condition for the rice quality formation and its distribution laws in China., 2002, 22(5): 636-642. (in Chinese with English abstract)
[4] 刘萍, 郭文善, 浦汉春, 封超年, 朱新开, 彭永欣. 灌浆期短暂高温对小麦淀粉形成的影响. 作物学报, 2006, 32(2): 182-188.
Liu P, Guo W S, Pu H C, Feng C N, Zhu X K, Peng Y X. Effects of transient high temperature during grain filling period on starch formation in wheat (L .)., 2006, 32(2): 182-188. (in Chinese with English abstract)
[5] 梁成刚. 水稻灌浆期高温对籽粒氮代谢酶活性及蛋白质含量的影响. 温江: 四川农业大学, 2010.
Liang C G.Effects of high temperature on the nitrogen metabolism enzymes activities and protein contents in rice grain-filling stage. Wenjiang:, 2010. (in Chinese with English abstract)
[6] 程方民, 张嵩午. 水稻籽粒灌浆过程中稻米品质动态变化及温度影响效应. 浙江大学学报: 农业与生命科学版, 1999, 25(4): 347-350.
Cheng F M, Zhang S W. The dynamic change of rice quality during the grain filling stage and effects of temperature upon it., 1999, 25(4): 347-350. (in Chinese with English abstract)
[7] 朱雪梅, 邵继荣, 杨文钰, 任正隆. 温度对杂交中籼稻灌浆期淀粉体发育及胚乳透明度的影响. 作物学报, 2005, 31(6): 790-795.
Zhu X M, Shao J R, Yang W Y, Ren Z L. Effect of temperature on the starch grain development and endosperm transparency of mediumhybrid rice during the period of grain filling., 2005, 31(6): 790-795. (in Chinese with English abstract)
[8] 李木英, 石庆华, 胡志红,潘晓华, 谭雪明. 高温胁迫对不同早稻品种胚乳淀粉合成酶类活性的影响. 中国农业科学, 2007, 40(8): 1622-1629.
Li M Y, Shi Q H, Hu Z H, Pan X H, Tan X M. Effects of high temperature stress on activity of amylosynthease in endosperm of earlyrice varieties., 2007, 40(8): 1622-1629. (in Chinese with English abstract)
[9] 王丰, 程方民, 刘奕, 钟连进张国平. 不同温度下灌浆期水稻籽粒内源激素含量的动态变化. 作物学报, 2006, 32(1): 25-29.
Wang F, Cheng F M, Liu Y, Zhong L J, Zhang G P. Dynamic changes of plant hormones in developing grains at rice filling stage under different temperatures., 2006, 32(1): 25-29. (in Chinese with English abstract)
[10] 成臣, 汪建军, 程慧煌, 罗亢, 曾勇军, 石庆华, 商庆银. 秸秆还田与耕作方式对双季稻产量及土壤肥力质量的影响. 土壤学报, 2018, 55(1): 247-257.
Cheng C, Wang J J, Cheng H H, Luo K, Zeng Y J, Shi Q H. Effects of straw returning and tillage system on crop yield and soil fertility quality in paddy field under double-cropping-rice system., 2018, 55(1): 247-257. (in Chinese with English abstract)
[11] Xie Z, Tu S, Shah F, Xu C X, Chen J R, Han D, Liu G R, Li H L, Imtiaz M, Cao W D. Substitution of fertilizer-N by green manure improves the sustainability of yield in double-rice cropping system in south China., 2016, 188: 142-149.
[12] 张军, 张洪程, 霍中洋, 李国业, 董啸波, 花劲, 郭保卫, 周培建, 程飞虎, 黄大山, 陈忠平, 陈国梁, 戴其根, 许轲, 魏海燕, 高辉. 不同栽培方式对双季晚粳稻产量及温光利用的影响. 中国农业科学, 2013, 46(10): 2130-2141.
Zhang J, Zhang H C, Huo Z Y, Li G Y, Dong X B, Hua J, Guo B W, Zhou P J, Cheng F H, Huang D S, Chen Z P, Chen G L, Dai Q G, Xu k, Wei H Y, Gao H. Effects of cultivation methods on yield and utilization of temperature and light of laterice in southern double cropping rice areas.2013, 46(10): 2130-2141. (in Chinese with English abstract)
[13] 郭保卫, 李超, 韦还和,王子杰, 许轲, 张洪程, 戴其根, 霍中洋, 魏海燕. 不同株高类型甬优系列籼粳杂交稻产量差异及其形成机理.核农学报, 2017, 31(1): 135-144.
Guo B W, Li C, Wei H H, Wang Z J, XU K, Zhang H C, Dai Q G, Huo Z Y, Wei H Y. Yield difference and its formation mechanism ofhybrid rice of Yongyou series in different types of plant height., 2017, 31(1): 135-144. (in Chinese with English abstract)
[14] Ahmed N, Tetlow I J, Nawaz S, Ahsan, Muhammad M, Muhammad S N R, Aisha B, David A L, Masahiko M. Effect of high temperature on grain filling period, yield, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice., 2015, 95(11): 2237-2243.
[15] Zhang H, Tan G, Yang L, Yang J C, Zhang J H, Zhao B H. Hormones in the grains and roots in relation to post-anthesis development of inferior and superior spikelets inhybrid rice., 2009, 47(3): 195-204.
[16] Jiang H W, Dian W M, Wu P. Effect of high temperature on fine structure of amylopectin in rice endosperm by reducing the activity of the starch branching enzyme., 2003, 63(1): 53-59.
[17] Ahmed N, Tetlow I J, Nawaz S, Iqbal A, Mubin M, Nawaz R M, Butt A, Lightfoot D, Maekawa M. Effect of high temperature on grain filling period, yield, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice.2015, 95(11): 2237.
[18] Natsuko A, Hiroki A, Hikari Y, Naoko F O, Rumiko I N C, Yasunori N, Naoko F. Relationships between starch synthase I and branching enzyme isozymes determined using double mutant rice lines., 2014, 14(1): 80.
[19] Brummell D A , Watson L M, Zhou J, McKenzie M J, Hallett I C, Simmons L, Carpenter M, Timmerman V G M. Overexpression of starch branching enzyme II increases short-chain branching of amylopectin and alters the physicochemical properties of starch from potato tuber., 2015, 15(1): 1-14.
[20] 王云飞, 张伟丽, 黄丹, 窦烨. 淀粉脱支酶的研究进展及应用. 中国酿造, 2008, 27(24): 25-26.
Wang Y F, Zhang W L, Huang D, Dou Y. The advance of starch-debranching enzyme and the application., 2008, 27(24): 25-26. (in Chinese with English abstract)
[21] Nakamura Y, Utsumi Y, Sawada T, Aihara S, Utsumi C, Yoshida M, Kitamura S. Characterization of the reactions of starch branching enzymes from rice endosperm., 2010, 51(5): 776-794.
[22] 吕艳梅, 谭伟平, 肖层林, 范美蓉, 廖育林. 高温对优质水稻籽粒淀粉形成及淀粉合成相关酶活性的影响. 华北农学报, 2014, 29(1): 135-139.
Lv Y M, Tan W P, Xiao C L, Fan M R, Liao Y L. Effect of high temperature on starch formation of grain and activities of enzymes related to starch synthesis of quality rice varieties., 2014, 29(1): 135-139. (in Chinese with English abstract)
[23] 周德翼, 张嵩午, 高如嵩, 朱碧岩. 稻米直链淀粉含量与结实期温度间的关系研究. 西北农业大学学报, 1994, 22(2): 1-5.
Zhou D Y, Zhang S W, Gao R S, Zhu B Y. The relationship between amylose content in rice grain and temperature in grain-filling stage.:, 1994, 22(2): 1-5. (in Chinese with English abstract)
[24] 蔡水文. 灌浆期间温度对不同用途早籼稻碳、氮代谢及其品质的影响. 长沙: 湖南师范大学, 2004.
Cai S W. Effect of temperature on metabolization of the element of carbon and nitrogen and characters of early-rice with different usage during grain filling stage.Changsha: Hunan Normal University, 2004. (in Chinese with English abstract)
[25] 程方民, 钟连进. 不同气候生态条件下稻米品质性状的变异及主要影响因子分析. 中国水稻科学, 2001, 15(3): 28-32.
Cheng F M, Zhong L J. Variation of rice quality traits under different climate conditions and its main affected factors., 2001, 15(3): 28-32. (in Chinese with English abstract)
[26] Gunaratne A, Sirisena N, Ratnayaka U K, Ratnayaka J, Kong X L, Arachchi L P V, Corke H. Effect of fertilizer on functional properties of flour from four rice varieties grown in Sri Lanka., 2011, 91(7): 1721-1726.
[27] MacLeod L C, Duffus C M. Reduced starch content and sucrose synthase activity in developing endosperm of barley plants grown at elevated temperatures., 1988, 15(3): 367-375.
[28] 杨欢, 沈鑫, 丁梦秋, 陆大雷, 陆卫平. 结实期高温胁迫对糯玉米子粒发育和内源激素含量的影响. 玉米科学, 2017, 25(2): 55-60.
Yang H, Shen X, Ding M Q, Lu D L, Lu W P. Effects of high temperature after pollination on grain development and endogenous hormone contents of waxy maize., 2017, 25(2): 55-60. (in Chinese with English abstract)
[29] Cao Z Z, Pan G, Wang F B, Wei K S, Li Z W, Shi C H, Geng W, Cheng F M. Effect of high temperature on the expressions of genes encoding starch synthesis enzymes in developing rice endosperms., 2015, 14(4): 642-659.
[30] 张桂莲, 廖斌, 武小金, 肖应辉, 肖浪涛, 陈立云. 高温对水稻胚乳淀粉合成关键酶活性及内源激素含量的影响. 植物生理学报, 2014, 50(12): 1840-1844.
Zhang G L, Liao B, Wu X J, Xiao Y H, Xiao L T, Chen L Y. Effect of high temperature on activities of enzymes associated with starch synthesis and hormones contents in endosperm of rice., 2014, 50(12): 1840-1844. (in Chinese with English abstract)
[31] Zhang G, Cheng Z, Zhang X, Guo X, Su N, Jiang L, Mao L, Wan J. Double repression of soluble starch synthase genes SSIIa and SSIIIa in rice (L.) uncovers interactive effects on the physicochemical properties of starch., 2011, 54(6): 448-459.
[32] 程方民, 钟连进, 孙宗修. 灌浆结实期温度对早籼水稻籽粒淀粉合成代谢的影响. 中国农业科学, 2003, 36(5): 492-501.
Cheng F M, Zhong L J, Sun Z X. Effect of temperature at grain-filling stage on starch biosynthetic metabolism in developing rice grains of early-.2003, 36(5): 492-501. (in Chinese with English abstract)
[33] 杨建昌, 彭少兵, 顾世梁, Visperas R M, 朱庆森. 水稻灌浆期籽粒中3个与淀粉合成有关的酶活性变化. 作物学报, 2001, 27(2): 157-164.
Yang J C, Peng S B, Gu S L, Visperas R M, Zhu Q S. Changes in activities of three enzymes associated with starch synthesis in rice grains during grain filling., 2001,27(2):157-164. (in Chinese with English abstract)
[34] 曹转勤, 杨建昌. 水稻强、弱势粒灌浆差异的激素调控机理研究进展. 中国稻米, 2014, 20(1): 12-16.
Cao Z Q, Yang J C. Research progress in hormonal regulation mechanism in the grain filling of superior and inferior spikelets of rice., 2014, 20(1): 12-16. (in Chinese with English abstract)
[35] 滕中华, 智丽, 吕俊, 宗学凤, 王三根, 何光华. 灌浆期高温对水稻光合特性、内源激素和稻米品质的影响. 生态学报, 2010, 30(23): 6504-6511.
Teng Z H, Zhi L, Lv J, Zong X F, Wang S G, He G H. Effects of high temperature on photosynthesis characteristics, phytohormones and grain quality during filling-periods in rice., 2010, 30(23): 6504-6511. (in Chinese with English abstract)
[36] 曾研华, 张玉屏, 潘晓华, 朱德峰, 向镜, 陈惠哲, 张义凯. 花后低温对水稻籽粒灌浆与内源激素含量的影响. 作物学报, 2016, 42(10): 1551-1559.
Zeng Y H, Zhang Y P, Pan X H, Zhu D F, Xiang J, Chen H Z, Zhang Y K. Effect of low temperature after flowering on grain filling and plant hormones contents in rice., 2016, 42(10): 1551-1559. (in Chinese with English abstract)
[37] 李军, 邓先瑞, 陈龙祥. 江西省寒露风的时空分布与预防措施. 华中师范大学学报: 自然科学版, 1994, 28(1): 123-128.
Li J, Deng X R, Chen L X. The characters of space-time distribution of cool-wind in “cool dew” period and its protections in Jiangxi Province.:, 1994, 28(1): 123-128. (in Chinese with English abstract)
[38] 李勇, 杨晓光, 叶清, 陈阜. 全球气候变暖对中国种植制度可能影响: Ⅸ. 长江中下游地区单双季稻高低温灾害风险及其产量影响. 中国农业科学, 2013, 46(19): 3997-4006.
Li Y, Yang X G, Ye Q, Chen F. The possible effects of global warming on cropping systems in China: Ⅸ. The risk of high and low temperature disasters for single and double rice and its impacts on rice yield in the middle-lower Yangtze plain., 2013, 46(19): 3997-4006. (in Chinese with English abstract)
Effects of Different Temperature from Full Heading to Milking on Grain Filling Stage on Grain Hormones concentrations, Activities of Enzymes Involved in Starch Synthesis and Accumulation in Rice Nanjing 9108
CHENG Chen, ZENG Yongjun*, CHENG Huihuang, TAN Xueming, SHANG Qingyin, ZENG Yanhua, SHI Qinghua
(,,,,,;,:)
【Objective】Our aim is to reveal the mechanism of the influence of different temperatures on grain-filling, starch and its components contents of laterice during early-middle grain filling stage. 【Methods】With a representativerice cultivar with high quality of Nanjing 9108 as material, high temperature (HT, average daily temperature of 31℃), medium temperature (MT, average daily temperature of 26℃) and low temperature (LT, average daily temperature of 21℃) were set during grain filling in plant incubators at natural temperature as control check (CK). The effects of high and low temperature on grain-filling, hormone contents, and activities of enzymes involved in starch synthesis and accumulation of laterice were studied during early-middle grain filling stage. 【Result】With rising temperature (21℃~31℃), the seed setting rate and 1000-grain weight of laterice were on the downward slide. The accumulation of total starch in grain was accelerated with shortened duration and decreased total contents. The amylose content and its proportion followed a decreasing trend during the whole grain filling period. The abscisic acid (ABA) content in grain was significantly increased during the early and middle grain-filling stage. The gibberellin (GA3) content decreased, and the zeatin riboside (ZR) content firstly increased and then dropped. The content of grain indole-3-acetic acid (IAA) was increased in MT plots during the grain filling stage. The IAA content in grains was reduced in HT and LT plots. The IAA content in the LT plots was decreased firstly and then increased, while in the HT plots, the content was consistently at lower level. The activity of soluble starch synthase (SSS) in grains was equal to that of ZR, which also showed ‘a increasing-decreasing trajectory’, but the SSS and the ZR activities were not synchronized in filling stage. The activity of grain starch branching enzyme (SBE) increased and the activity of grain starch decarboxylase (DBE) decreased.【Conclusion】The changes of grain endogenous hormones and activities of enzymes involved in grain starch synthesis of laterice were significantly influenced by different temperature, as well as the accumulation of starch and its components, grain filling in laterice.
laterice; temperature; grain; endogenous hormones; starch synthesis
10.16819/j.1001-7216.2019.8077
Q948.112+.2; S511.01
A
1001-7216(2019)01-0057-11
2018-06-15;
2018-08-28。
国家重点研发计划(2016YFD0300501, 2017YFD0301605);江西省水稻产业技术体系专项(JXARS-02-03);江西省重点研发计划(20161ACF60013, 20171BBF60030);江西现代农业科研协同创新专项(JXXTCX2015001-011)资助。
猜你喜欢
包装工程(2022年1期)2022-01-26
中国稻米(2021年6期)2021-12-02
今日农业(2021年15期)2021-10-14
中国农业科学(2021年7期)2021-04-21
农民致富之友(2020年32期)2020-12-03
农产品加工(2020年3期)2020-03-11
阅读(快乐英语高年级)(2019年8期)2019-09-10
小火炬·智漫悦读(2018年12期)2018-03-13
莫愁(2017年36期)2017-12-25
农村百事通(2017年8期)2017-04-26
