
干旱胁迫条件下信阳五月鲜桃光合特性和生理生化指标变化规律研究
2019-01-21 05:47阎腾飞黄玉杰李文杨赵师成
节水灌溉 2019年1期
阎腾飞,黄玉杰,李文杨,杨 乐,赵师成
(信阳农林学院,河南 信阳 464000)
0 引 言
干旱胁迫是目前影响植物生长发育和分布的首要因素,由水分胁迫造成的损失在所有非生物胁迫中占首要地位[1],探索植物干旱胁迫机制与光合作用和生物量之间的关系,成为节水农林业研究的核心目标。长期以来,国内外关于干旱胁迫对农作物、果树、木本植物的光合特性和生理生态做了大量研究,研究水平已深入到分子水平,特别是荧光特性的研究以及抗旱基因的研究[2,3]。大量的试验证明干旱胁迫对果树光合作用、膜透性和渗透调节物质具有重要影响[4]。因此,研究植物对干旱胁迫的反应机理,具有重要的理论价值和实践意义。
信阳五月鲜桃是信阳市平桥区林业局从乡土桃树自然芽变中选育出的优良品种,该品种成熟期早,果质优良,口感鲜美,适宜在豫南地区栽培推广。2008年经河南省林木良种审定委员会组织中国农业科学院郑州果树研究所、河南省林科院的有关专家现场品尝鉴定,被命名为“信阳五月鲜桃”(豫林审证字第100 号),确定为河南省林木良种[5]。信阳地处淮河流域,属于亚热带暖温带气候,降雨时空分布不均,年度差异明显,受传统农业思想的制约,豫南地区农业灌溉多以大水漫灌为主,靠天吃饭思想占主要因素,水资源浪费严重[6],因此,研究信阳五月鲜桃水分生理对于选择优良抗旱种质资源,提高水资源利用率具有重要意义,同时也为这种优良品种的推广提供了理论依据。
信阳五月鲜桃的研究目前仍停留在物候期观测、果实品质[5,7]等阶段,深入探索信阳五月鲜桃干旱胁迫条件下的反应机制的研究鲜有报道。董晓颖等研究了“超红短枝”和“早红珠”水分胁迫条件下桃叶片的水分利用效率和羧化效率的变化规律[8]。陈晓强等对不同类型桃品种的不同叶位的叶片光合特性进行了研究[9]。陈雪梅等[10]研究了水分胁迫对金蒲桃幼苗生长和生物量分配的影响。徐迪等研究了滴灌条件下不同土壤湿度日光温室“艳红”桃的需水规律[11]。笔者以信阳五月鲜桃幼树为试材,研究连续干旱胁迫条件下信阳五月鲜桃光合特性和生理生化特性,以期为今后信阳五月鲜桃的栽培应用和抗旱优良品种的选育提供可借鉴的材料,同时也为探讨信阳五月鲜桃水分生理提供理论依据。
1 材料与方法
1.1 试验材料
试验于2017年7月18日开始在信阳农林学院林学院试验站进行(地理位置为114°06′E、31°12.5′N)。海拔75~300 m。该地属亚热带向暖温带过渡地区,冷暖适中,四季分明,年平均气温15.3 ℃,无霜期长,平均为220~230 d;降雨丰沛,年均降雨量约1100 mm,空气湿润,年均相对湿度为77%。
以3年生信阳五月鲜桃幼树为试材,供试土为沙壤土;盆栽规格为口径40 cm,高50 cm。选取3盆,在盆上标号,选择晴朗的天气在傍晚对盆栽进行全灌水处理,使土壤条件充分饱和,达到田间持水量(FC)。然后放置在自然条件下持续干旱形成水分梯度(用遮雨布防止外界水分进入)。使用6050X3K1B Mini Trase Kit土壤水分速测仪(California,USA)在观测五月鲜桃叶片光合速率当日早上8∶00测定土壤体积含水量SVWC,%,直至叶片萎蔫为止,共形成5个土壤水分梯度系列。测定时,每株在盆内选取3个样点进行测定,每个样点记录3个读数。利用计算公式得到质量含水量SGWC,%,以及相对含水量SRWC,%。
SGWC=SVWC/ρ
(1)
SRWC=SGWC/FC
(2)
经计算,5个土壤水分梯度分别为92.5%,74.9%,55.7%,24.1%,14.3%。
1.2 试验方法
1.2.1 光合特性测定
选取树体中部、长势中庸的三片叶片,采用Li-6400型便携式光合仪从8:00-18:00测定盆栽幼树叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等指标以及光合有效辐射(PAR)、大气CO2浓度(Ca)、气温(Tair)、叶温(Tleaf)、空气相对湿度(RH)等微气象参数。每隔2 h测定一次,试验开始的前两日连续测量,采用试验开始后第二天的光合特性变化规律作日变化规律分析,之后每隔2 d测量一次。直至树体干旱萎蔫为止。叶片水分利用效率EL,μmol/mmol,和气孔限制值Ls,%,分别用下式计算:
EL=Pn/Tr
(3)
Ls=1-Ci/Ca
(4)
1.2.2 生理生化特性测定
在光合测定日于早上8∶00时采集新鲜的信阳五月鲜桃功能叶片测定生理生化指标,主要指标包括脯氨酸、可溶性糖含量、可溶性蛋白、MDA、POD、SOD,实验方法主要参考李合生等的试验方法[12],直至树体干旱萎蔫为止。
1.3 数据处理
数据处理采用SPSS分析,Excel制图。
2 结果与分析
2.1 光合特性变化规律
2.1.1 干旱胁迫条件下信阳五月鲜桃光合特性日变化规律
由图1,图2可以看出,信阳五月鲜桃光合速率日变化呈现先下降后回升再下降的双峰曲线,在早上8∶00时光合速率达到最大峰值为5.48μmol/(m2·s),随后逐渐下降,并在14∶00时降到“谷底”为2.47μmol/(m2·s),在16:00时出现回升,达到第二个峰值3.63μmol/(m2·s),随后降低。信阳五月鲜桃蒸腾速率呈现先升高后降低再升高的双峰曲线,在10∶00时达到第一个峰值1.91mmol/(m2·s),随后逐渐下降,在14∶00时降到“谷底”,为1.32mmol/(m2·s),并在16∶00达到第二个峰值1.74mmol/(m2·s),随后降低。信阳五月鲜桃水分利用效率呈现先降低后升高再降低的双峰曲线,在早上8∶00达到最大峰值3.54,最后逐渐下降,在12∶00降到“谷底”,随后逐渐回升,在16∶00时达到第二个峰值2.08,之后缓慢降低。
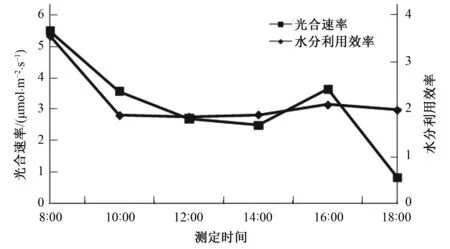
图1 信阳五月鲜桃光合速率、水分利用效率日变化规律
2.1.2 干旱胁迫条件下信阳五月鲜桃光合特性连日变化规律
由图3,图4可以看出,随着土壤相对含水量的不断降低,信阳五月鲜桃光合速率、蒸腾速率和水分利用效率都呈现先升高后降低的趋势,当土壤含水量降低到74.9%时,光合速率和蒸腾速率有一个明显的峰值,分别达到2.32和1.38mmol/(m2·s),之后逐渐下降,而水分利用效率随着土壤含水量的降低不断升高,在土壤相对含水量为24.1%时达到最大值,为3.01,之后开始下降。
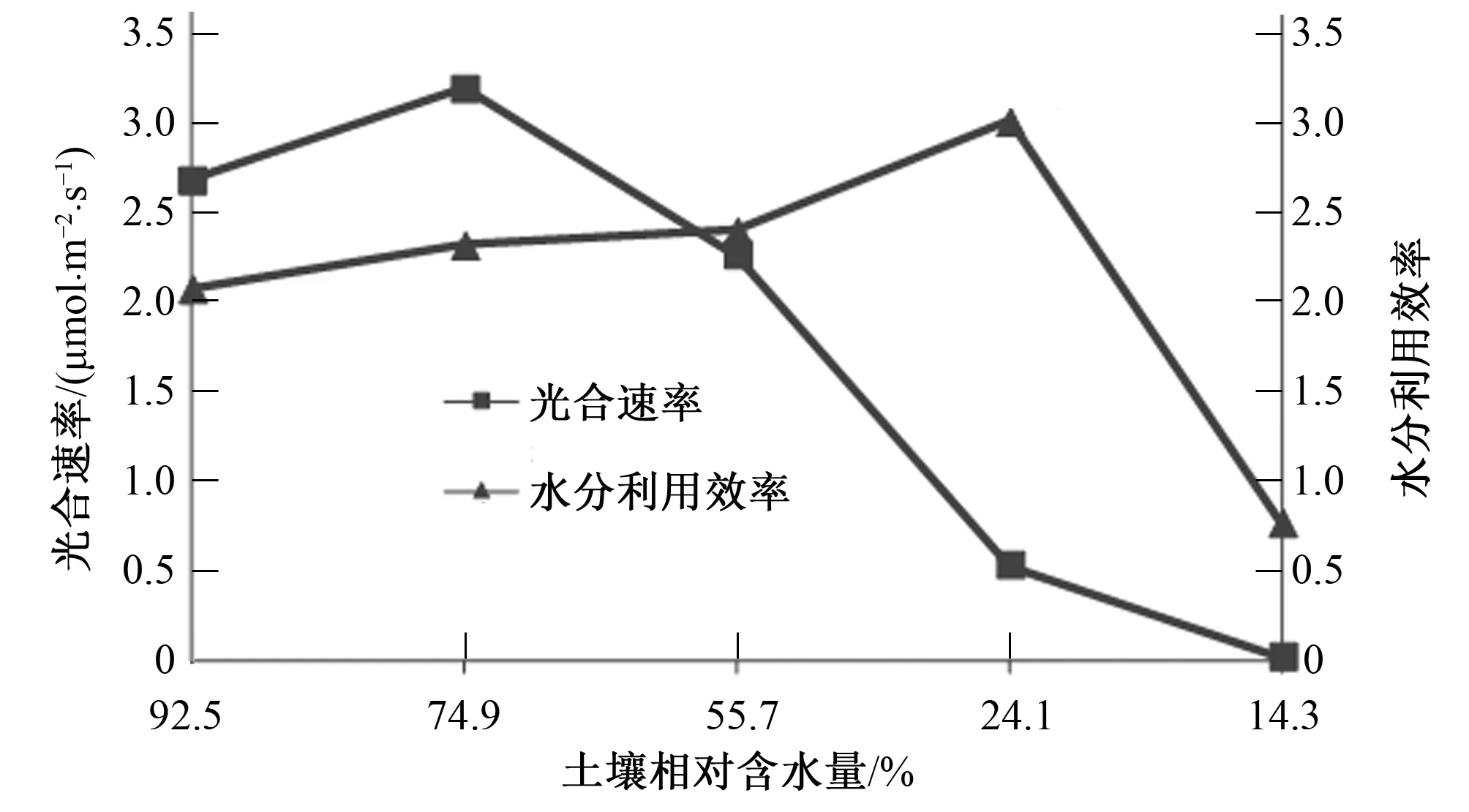
图3 信阳五月鲜桃光合速率、水分利用效率连日变化规律
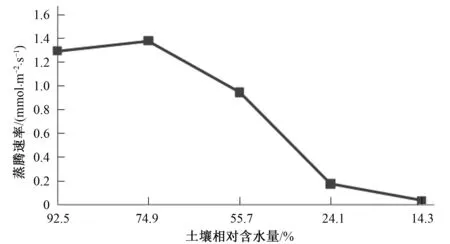
图4 信阳五月鲜桃蒸腾速率连日变化规律
由图5可以看出,随着土壤相对含水量的不断降低,信阳五月鲜桃胞间CO2浓度变化规律呈现先降低后升高的变化趋势,在土壤相对含水量为55.7%时,达到最低值为255.167mmol/mol。

图5 不同土壤相对含水量胞间CO2浓度变化
由图6可以看出,随着土壤相对含水量的不断降低,信阳五月鲜桃气孔限制值呈现先升高后降低的变化趋势,当土壤含水量为55.7%时达到最大值0.35%,然后逐渐下降。
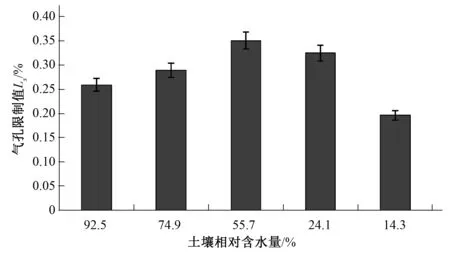
图6 不同土壤相对含水量气孔限制值变化
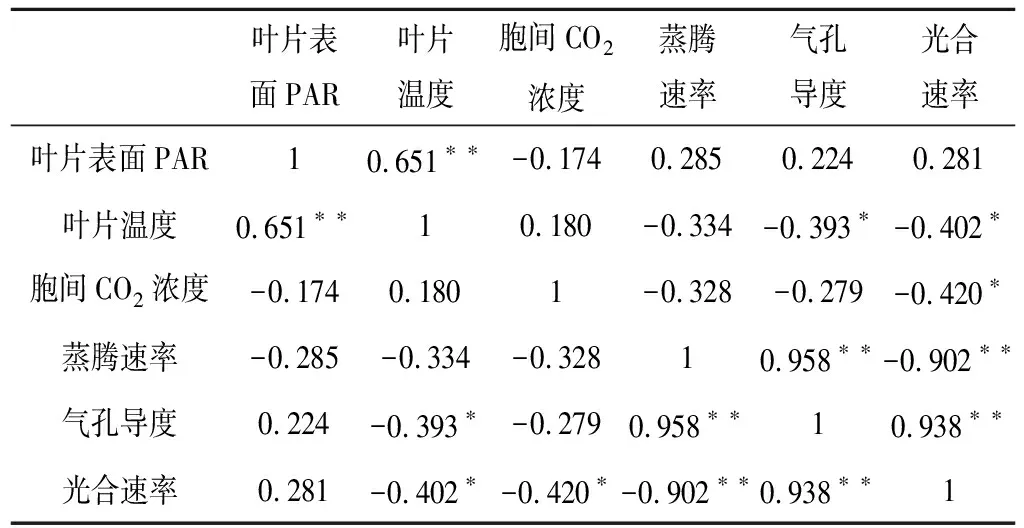
叶片表面PAR叶片温度胞间CO2浓度蒸腾速率气孔导度光合速率叶片表面PAR10.651∗∗-0.1740.2850.2240.281叶片温度0.651∗∗10.180-0.334-0.393∗-0.402∗胞间CO2浓度-0.1740.1801-0.328-0.279-0.420∗蒸腾速率-0.285-0.334-0.32810.958∗∗-0.902∗∗气孔导度0.224-0.393∗-0.2790.958∗∗10.938∗∗光合速率0.281-0.402∗-0.420∗-0.902∗∗0.938∗∗1
注:*表示在0.05水平下显著相关,**表示在0.01水平上显著相关。
将干旱胁迫条件下信阳五月鲜桃光合速率与叶片表面PAR、叶片温度、胞间CO2浓度、蒸腾速率、气孔导度进行相关性分析,由表一可以看出,光合速率与蒸腾速率呈现极显著的负相关(-0.902**),与气孔导度呈现极显著的正相关(0.938**),与叶片温度(-0.402*)和胞间CO2浓度(-0.420*)呈现显著的负相关。这说明信阳五月鲜桃光合速率与蒸腾速率和气孔导度具有极显著关系,与叶片温度和胞间CO2浓度具有显著关系。
2.2 生理生化特性变化规律
由图7、图8可以看出,随着土壤相对含水量的不断降低,信阳五月鲜桃在干旱胁迫条件下可溶性糖含量呈现先升高后降低的趋势,当土壤相对含水量为55.7%时,达到最大值43.30%,随后逐渐下降;可溶性蛋白含量在干旱胁迫条件下逐渐升高,增幅达到133%;脯氨酸含量在干旱胁迫条件下逐渐升高,增幅达587%;丙二醛含量在干旱胁迫条件下呈现先降低后升高的趋势,当土壤含水量为24.1%时,降到最低值22.16 μmol/g,FW;POD和SOD含量在水分胁迫条件下都变现为先升高后降低的趋势,在土壤含水量为55.7%时分别达到最大值48.76 u/(g·min)和148 u/(g,FW),随后逐渐下降。

图7 信阳五月鲜桃生理生化指标变化规律
3 讨论与结论
干旱胁迫是植物最普遍的逆境形式,对植物的光合作用有着重要影响。光合作用对坏境因子的影响高度敏感,植物受到干旱胁迫时光合作用受阻,净光合速率下降[13]。干旱胁迫可以通过各种途径降低果树植株的光合作用能力,抑制叶面积的伸展,降低叶绿体的光化学和生物化学活性[14]。试验结果表明:信阳五月鲜桃光合速率日变化在清晨8∶00时最大,随后逐渐下降,在中午14∶00降到最低,这可能是由于试验时间为夏季,叶表面光辐射强度自清晨开始迅速增强,叶温上升,气孔逐渐关闭,光合作用下降,在14∶00时降到最低,之后叶表面光合辐射强度下降,叶温降低,气孔张开,光合作用出现回升,有明显的“午休”现象,这与张征坤等[15]的研究结果是一致的。通过相关性分析表明,信阳五月鲜桃光合速率的变化与蒸腾速率、气孔导度、叶片温度和胞间CO2浓度具有密切的关系。这与黄拯[16]等研究结果是一致的。这表明在轻度或者快速干旱胁迫条件下,信阳五月鲜桃对于水分亏缺反应较为敏感,通过调节气孔的关闭降低叶片温度,进而降低光合速率,这是一种对于外界环境快速变化的自我保护机制。
干旱胁迫下植物光合特性一直都是研究的热点问题。影响光合作用的因素一般普遍认为主要是两个因素:气孔因素和非气孔因素[17]。气孔因素主要是由于水分胁迫导致气孔导度下降,进入叶片的CO2浓度降低从而使光合下降,非气孔因素主要是由于叶肉细胞的光合同化酶活性受损导致光合作用降低[18]。试验结果表明:随着土壤相对含水量的降低,当土壤相对含水量降低到74.9%时,光合速率、蒸腾速率、水分利用效率迅速上升,随后光合速率和蒸腾速率下降,水分利用效率继续上升。当土壤相对含水量低于24.1%时,水分利用效率下降。这说明当土壤相对含水量在74.9%时,干旱胁迫促进了信阳五月鲜桃的光合速率、蒸腾速率,进而提高了水分利用效率。当土壤含水量在74.9%到24.1%时,随着水分亏缺的程度加深,气孔导度降低,光合速率和蒸腾速率下降,由于蒸腾作用对于水分亏缺较光合作用更为敏感,水分利用效率上升。这表明适度的干旱胁迫能够有效地促进信阳五月鲜桃的光合特性。这与郭卫华、安玉艳等的研究结果是一致的[19,20]。Farqhar和Sharkey认为[21],当胞间CO2浓度(Ci)降低及气孔限制值(Ls)增大时,才可以做出光合速率Pn降低主要是由于气孔导读Gs降低所引起的结论。如果光合速率Pn降低,而胞间CO2浓度Ci升高,那么限制光合作用的主要因素肯定是非气孔因素,即植物叶肉细胞的光合活性下降。干旱胁迫条件下信阳五月鲜桃光合速率连日变化在土壤含水量大于55.7%时,胞间CO2浓度Ci降低,气孔限制值Ls上升。这表明当土壤相对含水量大于55.7%时,影响光合作用的主要因素是气孔因素,当土壤相对含水量低于55.7%时,干旱胁迫程度加深,光合作用相关酶活性降低,此时影响光合作用降低的主要因素是非气孔因素。
渗透调节是植物忍耐和抵御干旱的一种重要生理机制,是选择耐旱作物的一种重要的生理指标。渗透调节物质主要有无机离子(如K+,Cl-,Na+,Ca2+,Mg2+等)和有机溶质(如可溶性糖、脯氨酸、甜菜碱等)。Jones(1978年)指出,渗透调节作用一般发生在干旱胁迫程度较轻或者中度干旱胁迫情况下,逆境时植物体内累积各种有机物质和无机物质,增大细胞液浓度,降低渗透势,保持植物体内水分,适应逆境环境。当水分胁迫程度严重时,渗透调节能力减弱或者丧失[22]。试验结果表明:随着土壤相对含水量的不断降低,信阳五月鲜桃叶片内可溶性蛋白、脯氨酸不断积累;在土壤含水量降到55.7%之前,可溶性糖含量不断累积,随着土壤含水量的继续降低,可溶性糖含量又开始下降。这表明在当土壤相对含水量大于55.7%时,五月鲜桃渗透调节物质不断积累,维持细胞膨压,延缓植物组织由于失水所遭受的破坏。当土壤相对含水量低于55.7%时,水分亏缺加重,渗透调节能力减弱,可溶性糖向脯氨酸和可溶性蛋白补偿以提高植物组织抗逆性[23]。
水分胁迫会使植物积累大量的活性氧,从而造成对植物细胞的伤害,植物在长期的适应过程中,形成了抗氧化酶类保护系统,大量研究表明:SOD、POD、CAT和丙二醛(MDA)含量及膜透性是评价植物抗旱性的重要指标[24]。当土壤相对含水量降到55.7%之前,信阳五月鲜桃SOD、POD活性不断升高,随着土壤相对含水量的不断降低,信阳五月鲜桃SOD、POD活性逐渐降低;当土壤含水量降到24.1%之前,MDA含量不断上升,随着土壤相对含水量的降低开始下降。这表明当土壤相对含水量大于55.7%时,干旱胁迫能够促进以SOD为主导的细胞保护酶系统活性,提高信阳五月鲜桃抗旱性。当土壤相对含水量低于55.7%时,质膜系统遭到破坏,植物细胞受害程度加深,质膜过氧化。这与王娟等[25]的研究结果也是一致的。
植物的水分逆境生理是一个复杂的过程,是多种因素共同影响的结果。信阳五月鲜桃作为一种经济作物进行栽培与开发,可为豫南地区消费市场提供一种优良果品,进而产生巨大的经济、社会和生态效益,对信阳五月鲜桃水分胁迫的研究,可为进一步了解信阳五月鲜桃逆境生理、选择合适的栽培方式、筛选抗旱品种提供有力的理论依据,对于信阳五月鲜桃在稳定水分胁迫条件下的水分生理特性将是下一步的研究方向。
猜你喜欢
今日农业(2021年19期)2021-11-27
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
人大建设(2020年5期)2020-09-25
中国农业科技导报(2020年12期)2020-03-15
天津诗人(2019年4期)2019-11-27
农产品市场周刊(2017年27期)2017-08-09
美食堂(2017年5期)2017-05-19
电影故事(2016年16期)2016-11-21
电影故事(2016年5期)2016-06-15
