
Fossil Taiwannia from the Lower Cretaceous Yixian Formation of western Liaoning, Northeast China and its phytogeography significance
2019-01-05 02:01:00MingZhenZhangBaoXiaDuPeiHongJinBaiNianSun
MingZhen Zhang , BaoXia Du , PeiHong Jin 1,, BaiNian Sun
1. Key Laboratory of Petroleum Resources, Gansu Province/Key Laboratory of Petroleum Resources Research, Institute of Geology and Geophysics, Chinese Academy of Sciences, Lanzhou, Gansu 730000, China
2. Key Laboratory of Mineral Resources in Western China (Gansu Province) and School of Earth Science, Lanzhou University, Lanzhou, Gansu 730000, China
ABSTRACT Fossil Taiwania was discovered from the Lower Cretaceous Yixian Formation of Lingyuan City, western Liaoning Province, Northeast China. It is identified as a new species, Taiwania lingyuanensis sp. nov.. The present specimen is preserved as impressions with well defined leaf shoots system and reproductive structures. Leaves are dimorphic, spirally and imbricately arranged. They are scale-like on the main and cone-bearing branchlets, and subulate to falcate-subulate on the juvenile or sterile shoots. The seed cones are singly elliptic, ovate or elongate-ovate and terminally borne on ultimate shoots, bearing 22-24 scale-bracts complexes imbricately and helically arranged around the cone axis, the bracts are broadovate, rhomboidal or hexagonal with entire margins. Both the leafy shoots morphology and reproductive structures are similar to extant Taiwania. Furthermore, geological distribution and molecular biological evidences support that Taiwania is probably originated from the eastern Asia at least in the Early Cretaceous and widely distributed in the North Hemisphere thereafter.
Keywords: Taiwannia; Lower Cretaceous; Yixian Formation; leaves; seed cones
1 Introduction
The extant cupressaceaeous genus Taiwania Hayata is endemic to East Asia (Farjon and Garcia,2003; Zhou and Momohara, 2005; Li et al., 2008),with only one species, Taiwania cryptomerioides Hayata distributed in a localized and remarkably disjunct area in South China, northern Myanmar and northern Vietnam (Farjon and Garcia, 2003; Farjon,2005, Ma et al., 2009; Chou et al., 2011). However,the fossil records of vegetative and reproductive remains of Taiwania indicate a much wider distribution that extended into the polar regions of the Northern Hemisphere during the Mesozoic and Cenozoic(LePage, 2009; Chou et al., 2011). Extant Taiwania is characterized by spirally arranged, shortly decurrent and imbricate leaves, leaves dimorphic, juvenile leaves falcate-subulate and mature leaves scale-like(Farjon and Garcia, 2003; Ma, 2003; LePage, 2009),the ovuliferous cones are ovate or short-cylindrical with appressed, serrulate and slightly keeled bractscales (Farjon and Garcia, 2003; LePage, 2009), and the ovuliferous scales produce two ovules on the base of the ventral side of each bract (Farjon and Garcia,2003; Schulz and Stützel, 2007).
Members of fossil taxodiaceous Cupressaceae are close to extant taxon based on morphological features.Yezosequoia resembles Sequoia or Sequoiadendron morphologically, while resembles Sequoia and Cunninghamia micro-morphologically (Nishida et al.,1991; Ohsawa, 1994). Parataxodium is related to Taxodium and Metasequoia (Arnold and Lowther, 1955),while Austrohamia is close to Athrotaxis and Cunninghamia (Escapa et al., 2008; Zhang et al., 2012).The current fossil plant bears subulate or scale-like leaves, which are similar to those of living Athrotaxis,Taiwania and Glyptostrobus, while the ovuliferous cones shows similarities to those of living Cunninghamia and Taiwania. Presently, the fossil specimens preserved as leafy twigs with associated seed cones is assigned to Taiwania morphologically. Moreover, the distribution and evolution of the genus Taiwania are discussed based on the fossil records and modern phylogenetic analysis.
2 Materials and methods
The present fossil plant was collected by the western Liaoning Expedition team of the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences from the Lower Cretaceous Yixian Formation at Dawangzhangzi Village, Lingyuan City, western Liaoning Province, Northeast China(Figure 1). The fossils are preserved as impressions,and collected from the grey to yellowish shale of the Dawangzhangzi bed of Yixian Formation. Grey to yellowish mudstone with horizontal bedding and rich fossil products (Zhou et al., 2003) indicated a lacustrine sedimentary environments for the Yixian Formation. U-Pb and40Ar/39Ar dating suggests a late Early Cretaceous age of the Yixian Formation (Smith et al.,1995; Swisher et al., 1999; Wang et al., 2001; Swisher et al., 2002; Zhu et al., 2002; Zhou et al., 2003; He et al., 2006; Zhou, 2006; Sha, 2007; Yang et al.,2007; Zhu et al., 2007; Chang et al., 2009), and most data support a Barremian to Aptian age (Chang et al.,2009), and the Dawangzhangzi bed is dated as early Aptian (Zhou et al., 2003; Meng et al., 2008; Sun et al., 2011).
3 Systematic Description
Order: Coniferales
Family: Cupressaceae
Genus: Taiwania Hayata 1906
Species: Taiwania lingyuanensis B.X. Du and B.N. Sun sp. nov. 2014 (Plate I and Plate II).
Holotype: B0031, the specimen are housed in the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing,100044, China.
And the Princess, who had learnt to find diversion in very small things in her quiet life, was in a continual state of delight over the treasures which the Prince constantly laid where she must find them
Type locality: Dawangzhangzi Village, Lingyuan City, Liaoning Province, Northeast China.
Stratigraphy: Dawangzhangzi bed, Yixian Formation
Age: Early Cretaceous.
Etymology: the specific epithet is after the name of the fossil locality, Lingyuan City.
Diagnosis: Branchlets with terminally borne on ultimate shoots seed cones. Branches irregularly alternate. Leaves dimorphic, spiral and imbricate on the leafy shoots. Leaves scale-like on the main branch and fertile branches, ovoid or rhomboidal, leaf base decurrent and appressed to the leafy shoots, leaf apex acute or sub-acute; Leaves subulate or falcate-subulate on the young or sterile branches, leaf base decurrent, and leaf apex acute or sub-acute, slightly incurved. Seed cones sessile and elliptic, ovate or elongate, narrowly towards the apex, 10.7-13.0 mm long and 6.5-7.5 mm wide, with 22-24 scale-bracts complexes helically arranged around the cone axis. Cone scales imbricate and tightly, broad-ovate, rhomboidal or hexagon in shape, margins entire, base broadrhomboidal, triangular or round and apex gradually tapering.
The present specimen was preserved as leafy branches bearing single terminal and sub-terminal ovulate cones (Plate I, 1-2). Shoot shows up to three orders of branching (Plate I, 1-2). The main branch is slightly curved, about 12 mm long and 3 mm wide(Plate I, 1-2, 4-6, 8). The lateral branches are slender,irregular and alternate, commonly cured and about 1.5 mm in diameter (Plate I, 3, 5-7, 9-10).
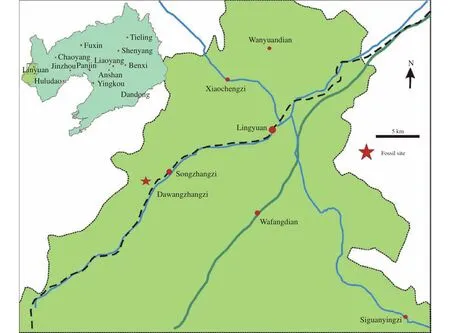
Figure 1 Location of the fossil site in western Liaoning, Northeast China
Leaves are dimorphic, spirally arranged and adpressed to the twigs (Plate I, 2-10). Leaves on the main branch are scale-like, more or less rhomboidal and ovoid, 3.0-4.0 mm long by 2.0-2.5 mm wide, the greater part of the leaves is very appressed and is more or less fused to the twig. Leaf bases are decurrent, leaf tips are acute or sub-acute (Plate I, 4-8);Leaves on the lateral fertile branches are elongated scale-like and more or less ovate, about 2.0-2.5 mm long and 0.8-1.2 mm wide, spirally and tightly arranged, with a decurrent oval base and a distinctive acuminate apex (Plate I, 3, 5-7, 9-10). Leaves on the juvenile or lateral sterile branches are subulate or falcate-subulate, about 1.5-3.0 mm long and 0.7-1.2 mm wide, slightly incurved and some time spread up towards the apex (Plate I, 3, 6-7, 9-10).
Five seed cones are borne singly on the terminal to sub-terminal twigs (Plate II, 1). The seed cones are sessile and elliptic, ovate or elongate-ovate in shape(Plate II, 2-3). The broadest par is near the middle of the cone (Plate II, 3). Seed cones are about 10.7-13.0 mm long and 6.5-7.5 mm wide (Plate II, 1-3). Each cone has about 22-24 scale-bracts complexes arranged helically around the cone axis (Plate II, 2-3).
The scale-bracts are broad-ovate, rhomboidal or hexagon with entire margins, imbricate, spirally and tightly arranged around the cone axis, about 2.2-3.4 mm wide and 2.5-4.0 mm long, and the base of the scales is broad-rhomboidal, triangular or round in shape, and reduced apically. The apex is acute or sub-acute, more or less rounded (Plate II, 3-5, 7-9). The middle portion is expanded, shows a distinctive keel (convex) on the abaxial surface (Plate II, 3-4, 8). The ovuliferous scale develops as a small lob with irregularly sinuate margins, 2-3 ovules can be barely observed (Plate II,5-6, 9).
4 Discussions
4.1 Compared to extent conifers
The Coniferopsida is divided into four orders mainly according to the morphological features of the reproductive structures. The Pinales, including Araucariaceae, Cupressaceae sensu lato, Pinaceae and Sciadopityaceae are characterized by having typical female cones with compact ovuliferous complexes helically arranged (Table 1), while other families, including Podocarpaceae Cephalotaxaceae and Taxaceae are characterized by having atypical female cones, which can be called seed-bearing structures,with bracts at the bases of the seeds (Saiki and Kim-ura, 1993; Fu et al., 1999e, 1999f, 1999g). The present fossil plant are preserved as impressions consists of both vegetative and compact seed cones, leave twigs are characterized by scale-like or subulate leaves helically and more or less imbricately arranged; the seed cones are terminal with numerous scale-bracts helically and imbricate arranged (Figure 2a), which can be compared to Araucariaceae, Cupressaceae sensu lato, Pinaceae and Sciadopityaceae. However,among conifers, scale-like or subulate and spirally arranged leaves are common in Araucariaceae, Podocarpaceae and Cupressaceae sensu lato (Fu et al.,1999a, 1999d, 1999e; Du et al., 2013). Therefore, the present specimen can be compared to Araucariaceae and Cupressaceae sensu lato both on the leafy shoots morphology and productive structures (Table 1).

Plate II Seed cone morphology of Taiwania lingyuanensis (1-3. singly and terminally inserted seed cones,scale bar=5 mm; 4. scale-like leaves and rhomboidal scale-bracts of Taiwania lingyuanensis, shows the distinctive keel on the abaxial surface, scale bar=5 mm; 5. spirally and imbricate scale-bracts of Taiwania lingyuanensis,shows the trace of seeds, scale bar=5 mm; 6. scale-bracts morphology of Taiwania lingyuanensis, shows the trace of the small lob with irregularly sinuate margins, scale bar=5 mm; 7. scale-bracts morphology at the base of the seed cone, scale bar=2 mm; 8-9. scale-bracts morphology at the middle and upper part of the seed cone, scale bar=2 mm)
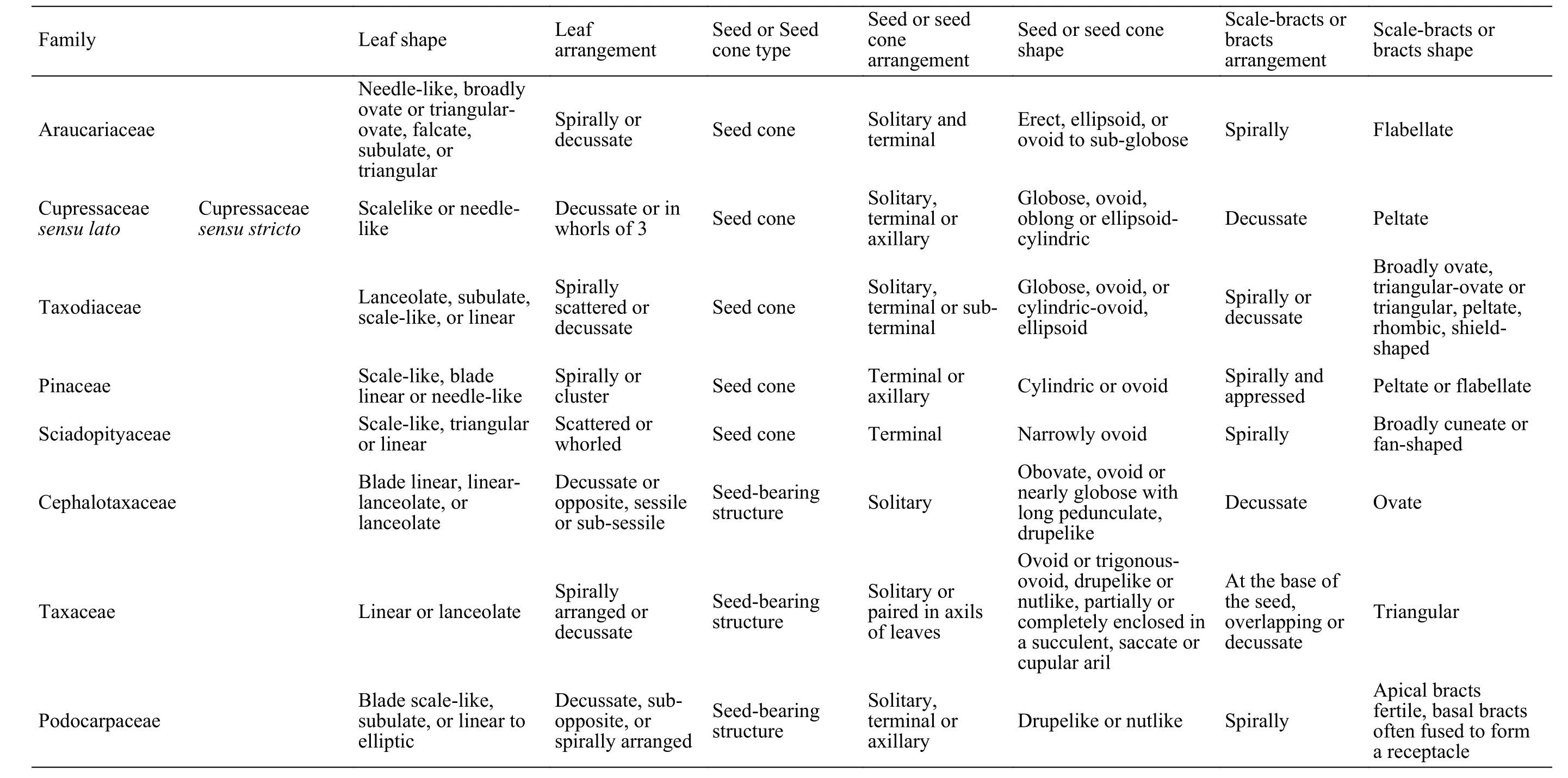
Table 1 Morphological comparisons amongthe extant families of Coniferopsida
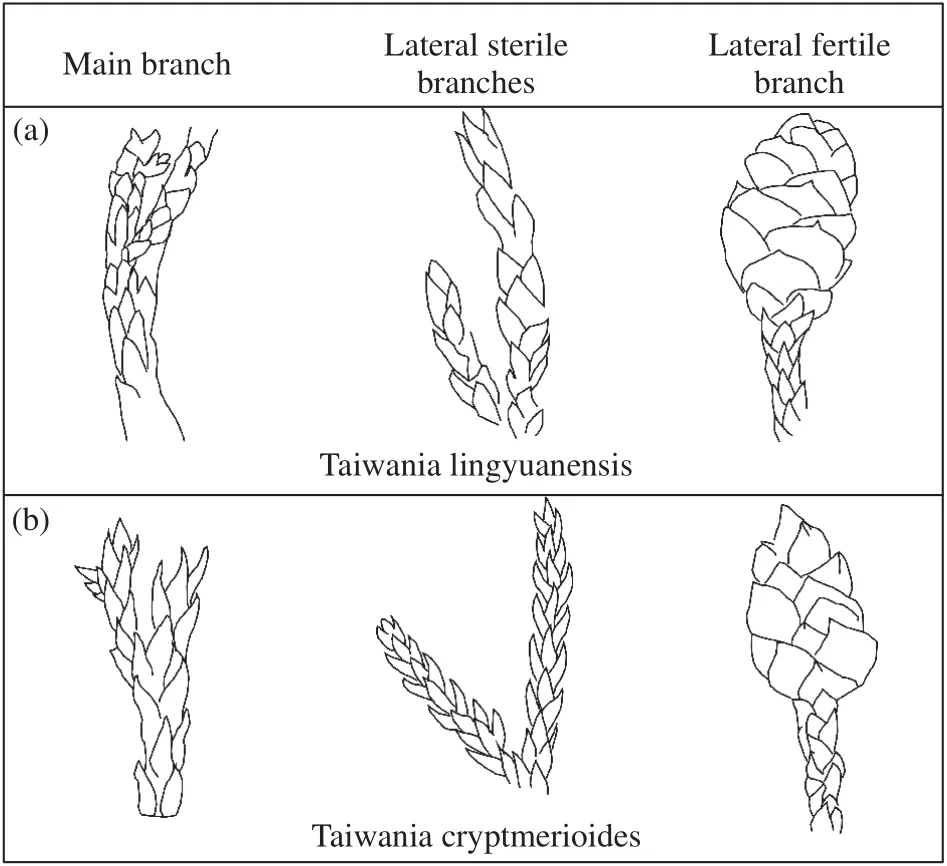
Figure 2 Comparison of morphological structures between the holotype and the extant Taiwania (a: foliage of the holotype; b: foliage of Taiwania cryptmerioides),derive from Delectis Florae Reipublicae Popularis Sinicae Agendae Academiae Sinicae, 1978
Leaves of the Araucariaceae are extremely variable in morphology (Stockey, 1982, 1994). Agathis is characterized by broad, flattened, lanceolate to ovate leaves with inconspicuous veins in parallel arrangement, leaves are shortly petiolate, spirally arranged on the main branches and opposite or alternate arranged on the lateral branches (Stockey, 1994; Fu et al.,1999a; Kunzmann, 2007), which are quite different from the present specimen. Leaves of Araucaria are sessile and decurrent, which is similar to the present specimen. While Araucaria is distinguished by broadtriangular, curved, needle-like, lanceolate or subulate shaped leaves and reflexed or upcurved seed cone bracts apices (Stockey, 1994; Fu et al., 1999a; Kunzmann, 2007).
Scale-like leaves are common in Cupressaceae sensu lato. Thereinto, Cupressaceae sensu stricto have verticillate or decussate arranged leaves (Bartel et al.,1993; Fu et al., 1999d; Farjon, 2005; Schulz et al.,2005; Taylor et al., 2009), while helically arranged leaves occur in taxodiaceous genera, such as Cryptomeria, Sequoia, Glyptostrobus, Taxodium, Taiwania,Sequoiadendron and Athrotaxis (Fu et al., 1999c; Ma and Li, 2002; Ma et al., 2004; Schulz et al., 2005; Ma et al., 2009; Du et al., 2012). Among them Athrotaxis,Glyptostrobus, Sequoiadendron and Taiwania have subulate or scale-like leaves (Fu et al., 1999c; Schulz et al., 2005; Ma et al., 2009; Du et al., 2012), which can be compared with the present specimen. Leaves of Athrotaxis are scale-like or needle-like, apically spreading or closely appressed to the twigs (Schulz et al., 2005; Dong et al., 2014), while scale-like leaves are serrate on the leaf margins (Schulz et al., 2005),which is distinct from the present specimen. Leaves of Glyptostrobus are trimorphic: leaves on the main, perennial and fertile branches are scale-like, while leaves on the annual branches of mature trees are subulate and quadrangular in cross section, and leaves of annual branches of young trees and seedlings are linear in shape (Fu et al., 1999b; Ma et al., 2004). Moreover,the seed cones of Glyptostrobus are shortly pedunculate, with 6-10 triangular, acute teeth at distal margin of the cone scales (Fu et al., 1999c), which cannot be observed on the current specimen. Leaves of Sequoiadendron are subulate and scale-like, ovate to lanceolate, loosely imbricated and appressed to branches or radically spreading on the axis, which are quite similar to the present specimen, while the seed cones are pedunculate (Fu et al., 1999b). Leaves of extant Taiwania are falcate-subulate in the juvenile stage and scale-like in the mature stage (Fu et al., 1999c; Farjon and Garcia 2003; Ma et al., 2009), which are similar to the present specimen (Figure 2b and Table 1).
As the current fossil is poorly preserved, lack of cuticular features for micro-morphological comparison, while the macro-morphological features, such as the vegetative and reproductive morphology are well identified. However, the seed cone morphology resembles extant Cunninghamia and Taiwania morphologically, such as the shape of the seed cones and bract-scale complexes. While comparing to Cunninghamia, the size of seed cone is much smaller, and the number of the inserted bract-scales are less of the present specimen (Table 2). The bract-scale number of the present specimen is 22-24, showing more similarities to Taiwania cryptmerioides (Figure 2). Therefore, based on the external morphology of leafy shoots, seed cones and bract-scale complexes, the present specimen is related to Taiwania.
4.2 Comparison with fossil Taiwania and Taiwania affinity species
Leafy shoots and seed cones of Taiwania have been reported from the Lower Cretaceous to the Pliocene of the Northern Hemisphere (LePage, 2009). Vegetative shoots bearing seed cones from the Paleocene of Spitzbergens were described and illustrated as Taiwania schaeferi by Schloemer-Jäger (1958), and they are consistent with modern Taiwania both in leaf morphology and reproductive structures (Ferguson,1967). It has also been reported from the Lower Miocene of eastern Germany (Jähnichen, 1998). Taiwania cf. schaeferi was described from the Late Oligocene of Kleinsaubernitz (Walther, 1999). Kilpper (1968)described T. paracryptomeroides based on seed cone and leaf with cuticle from the late Miocene of Rheinland, foliar branches of T. cf. paracryptomeroides with well preserved cuticle have been reported from the Miocene of Weisselster Basin (Mai and Walther,1991). In addition, fossil Taiwania, such as T. mesocryptomeroides, T. cretacea, and T. eocenica are reported from the Miocene to Pliocene of Japan (Matsuo, 1967, 1970; Ishida, 1970; Nishida et al., 1992).Recently, T. cf. T. cryptomeroides was described from the Lower Cretaceous of Alaska, North America, and it quite resembles modern T. cryptomeroides in leafy shoots and seed cone morphology (LePage, 2009).
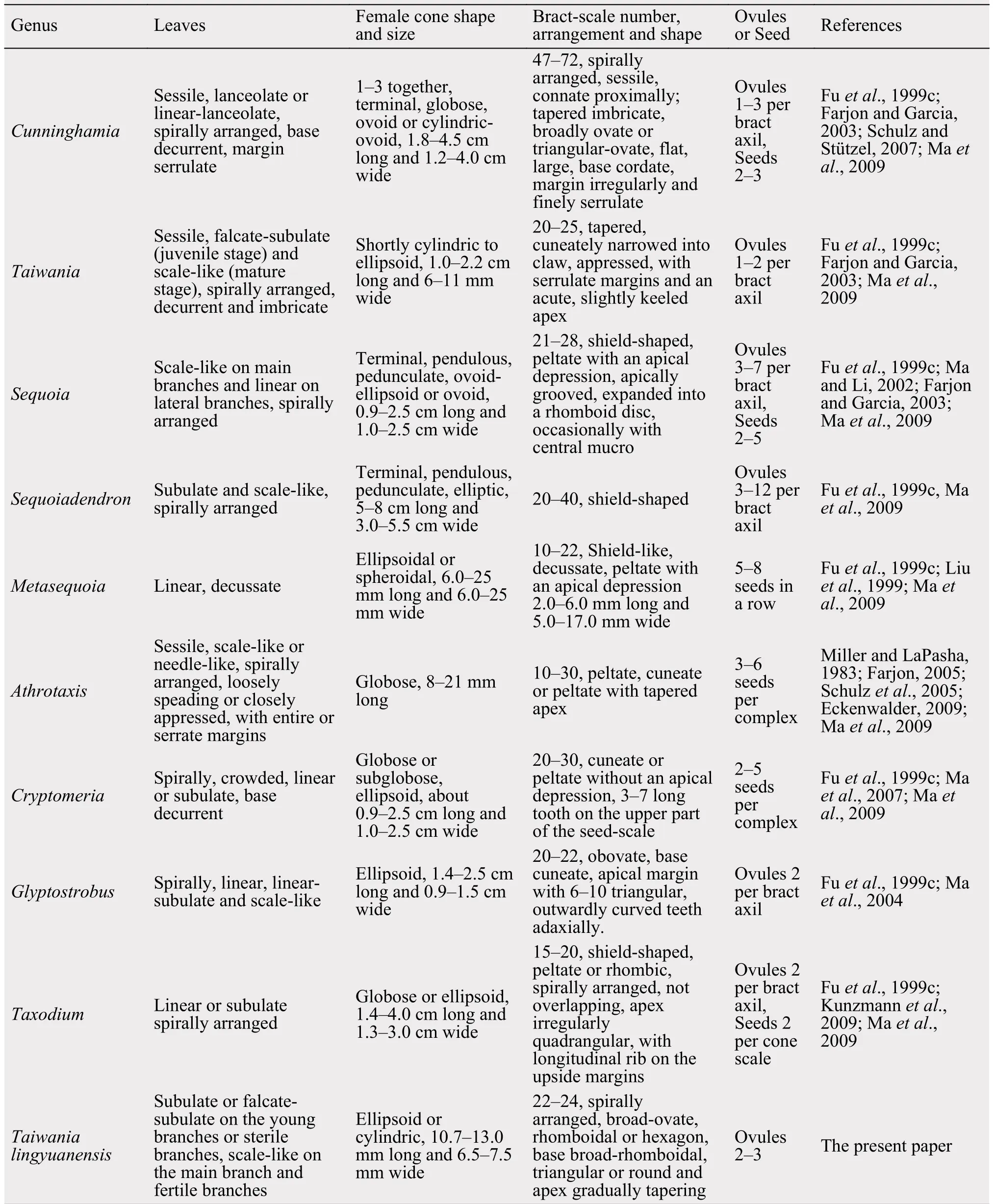
Table 2 Morphological comparisons between taxodiaceous genera and the present specimen
Among the fossil specimens of Taiwania (Table 3),T. mesocryptomeroides, T. japonica, T. microphylla,T. schaeferi and T. cf. T. cryptomeroide have dimorphic (subulate and scale-like) leaves (Schloemer-Jäger, 1958; Kilpper, 1968; Sveshnikova and Budantsev, 1969; Ishida, 1970; Matsuo, 1970; Jähnichen,1998; LePage, 2009), which are consistent with the present specimen, seed cones of T. mesocryptomeroides are only 6-8 mm long and 4-5 mm wide(Matsuo, 1970), which is quite smaller than the present specimen. The subulate leaves of T. japonica are up to 14 mm long (Ishida, 1970), while leaves of T. microphylla and T. schaeferi are up to 7 mm long(Sveshnikova and Budantsev, 1969; Jähnichen, 1998),which are much longer than the present specimen.The present specimen resembles the Alaskan specimens T. cf. T. cryptomeroide in leaf shape, size, seed cone shape and size, while subulate leaves are much falcate, and the scale leaves are more triangular than T. cf. T. cryptomeroide, and the scale leaves apex of T. cf. T. cryptomeroide is sharply recurved towards the branches (LePage, 2009), which cannot be observed in the present specimen.
Another two fossil genes Mikasastrobus and Parataiwania show similarities to extant Taiwania in seed cone morphology (Nishida et al., 1991; Nishida et al., 1992; Saiki and Kimura, 1993). Mikasastrobus is described on the basis of six permineralized seed cones and a leafy shoot collected from the Upper Cretaceous of Hokkaido, Japan. The seed cone arrangement, shape and cone scale complexes arrangment are similar to the present specimens, while Mikasastrobus is differs in having needle-like leaves and 4-5 inverted ovules (Saiki and Kimura, 1993); Parataiwania is represented by a seed cone collected from the Upper Cretaceous of North Japan (Nishida et al.,1991; Nishida et al., 1992). The seed cone is ovoid to ellipsoid, approximately 22 mm long and longitudinal sections show about 30 helically arranged bractscale complexes (Nishida et al., 1991), which are quite different from the current specimen.
According to the detailed comparisons of both leafy shoots and female cone morphology of fossil species of Taiwania or related to Taiwania, the present plant fossil is defined as a new species,Taiwania lingyuanensis sp. nov. which is close to the extant Taiwania morphologically.
5 Distribution and evolution
Modern Taiwania is endemic to low-latitude areas of eastern Asia (Figure 3), where the climate is warm and humid (Chou et al., 2011). And it has remained almost unchanged morphologically for over 100 Ma(LePage, 2009; Chou et al., 2011). While fossil record indicates a wide distribution in the middle-high latitude areas of Asia and Europe and high-latitude areas of North America (Figure 3), with a diverse range of ecological conditions (LePage, 2009; Chou et al., 2011).
Modern phylogenetic analysis shows that the stem lineage of Taiwania appeared around the Late Triassic, probably 170-238 Ma (Mao et al., 2012). The Ancestral area reconstruction (AAR) suggests that living Taiwania originated in Asia and later its early members expanded to North America and Europe(Mao et al., 2012). The earliest occurrence of Taiwania, based on the fertile and vegetative remains,is recovered from the early Aptian deposits of middlelatitudes in Northeast China, indicates that the genus has already originated at least in the late Early Cretaceous from the middle-latitude areas of Asia, and its ancestral members extended to the high-latitude region of North America through Beringian Corridor during the Albian time (LePage, 2009). Due to the overall warm climate in the middle-high latitude areas during the middle-late Cretaceous (Huber et al., 2002;Herman and Spicer, 2010; Linnert et al., 2014; Wang et al., 2014), the distribution area expanded north to the high Arctic areas of Russia (Figure 3).
The palaeoclimate was characterized by a relatively warm climate during the Paleocene to Eocene and a cooler condition from the Oligocene to the Pleistocene (McIver and Basinger, 1999; Zachos et al., 2001; Williams et al., 2008; Leng et al., 2010).During the Paleogene and Eocene, the distribution of Taiwania has expanded to the north-polar region(McIver and Basinger, 1999). As climate deteriorated globally during the middle Miocene-Pliocene (Chou et al., 2011), the geographical extent of their former widespread distribution contracted to the middle-latitude areas. By the Pliocene and Pleistocene, the fossil record is mainly occurred in Japan (Momohara, 1999;LePage, 2009).
Furthermore, Cunninghamia is reported to be the genus most closely related to Taiwania based on morphological and molecular characters (Gadek et al.,2000; Farjon and Garcia, 2003; Stockey et al., 2005),and its ancestral stocks might be traced back to the Early Cretaceous of Northeast China (Meng et al.,1988; Yu, 1995). Moreover, the common ancestor of both Taiwania and Cunninghamia appeared around the Early Cretaceous (Meng et al., 1988; Yu, 1995;Chou et al., 2011). Consequently, Taiwania is probably originated from Asia during the Early Cretaceous and widespread to North America and Europe afterwards, which is supported by both fossil records and modern phylogenetic analysis.
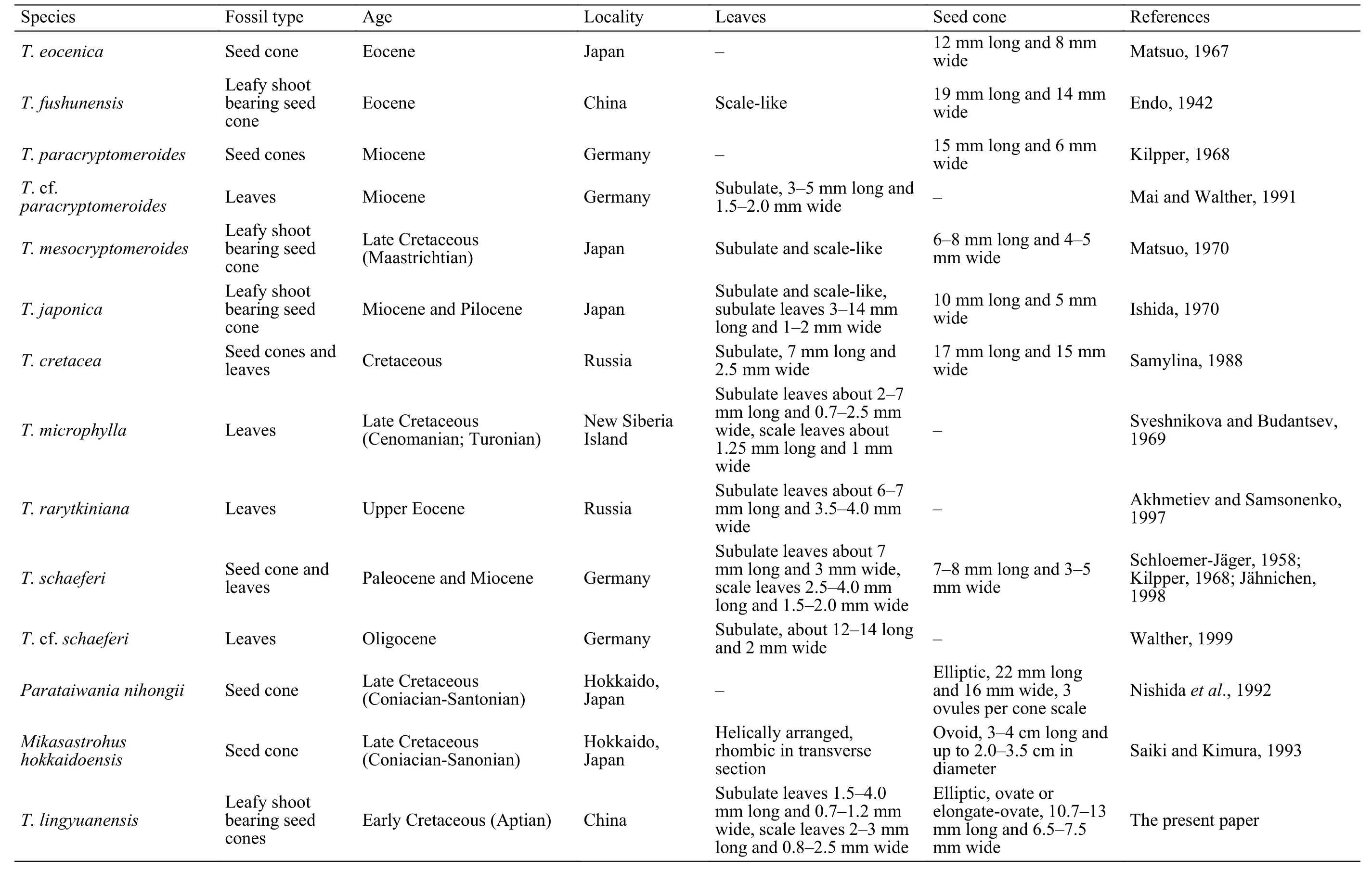
Table 3 Morphological comparisons among the reliable fossil records of Taiwania and Taiwania-like species
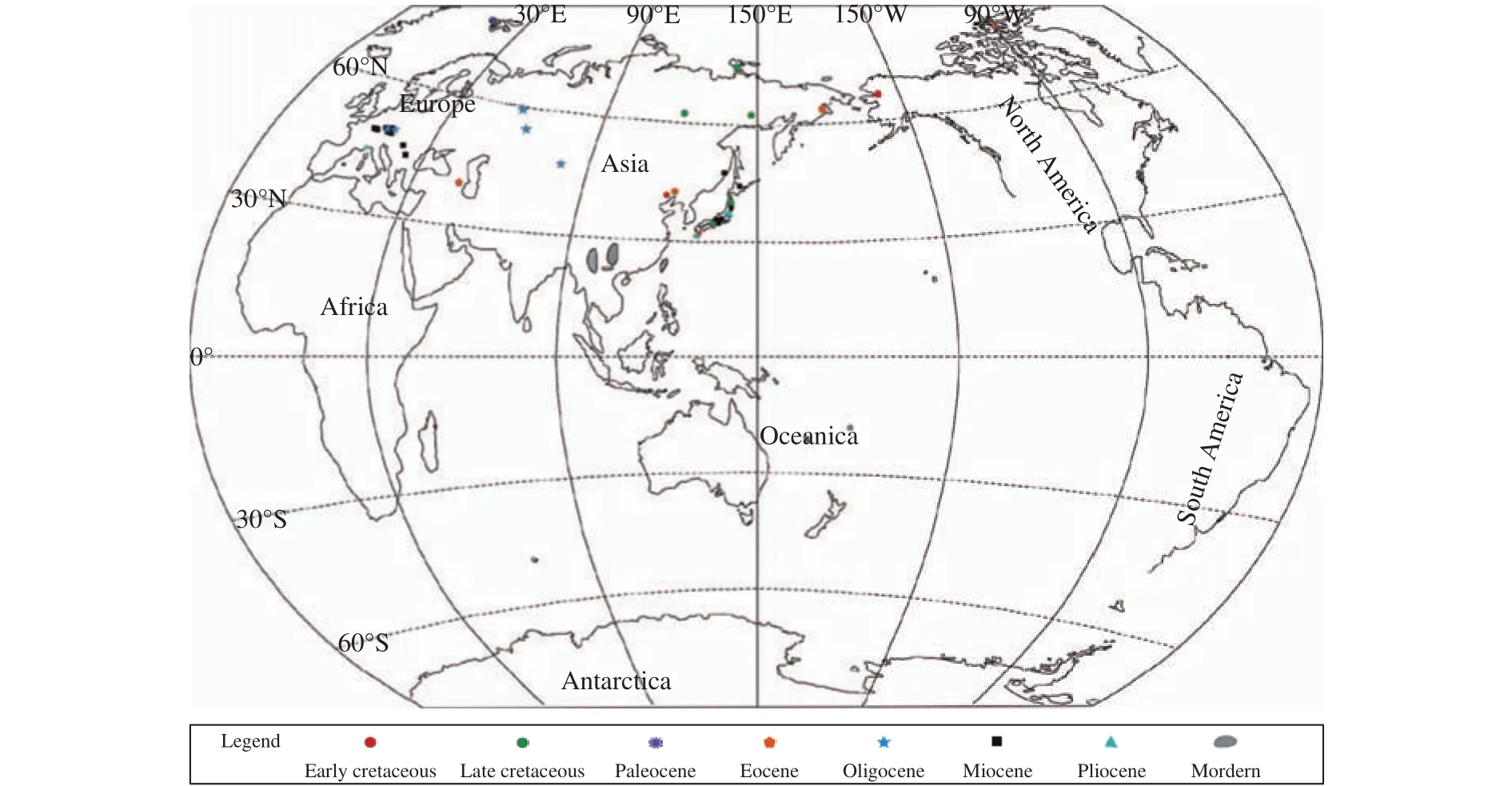
Figure 3 Fossil and extant distribution of Taiwania (Data from: Endo, 1942; Schloemer-Jäger, 1958; Matsuo, 1967; Kilpper, 1968;Sveshnikova and Budantsev, 1969; Ishida, 1970; Matsuo, 1970; Samylina, 1988; Mai and walther, 1991; Nishida et al., 1992;Akhmetiev and Samsonenko, 1997; Jähnichen, 1998; McIver and Basinger, 1999; Walther, 1999; LePage, 2009)
Acknowledgments:
We are grateful to Prof. XiaoLin Wang from the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing,China for providing the present fossil specimen. This work was conducted under the 973 Program of China(No. 2012CB822003), the National Natural Science Foundation of China (Nos. 41402007, 41602023) and the Fundamental Research Funds for the Central Universities (Lzujbky-2014-132).
 Sciences in Cold and Arid Regions2018年6期
Sciences in Cold and Arid Regions2018年6期
- Sciences in Cold and Arid Regions的其它文章
- The 2018 Academic annual meeting of China Society of Cryospheric Science was held successfully in Foshan on November 17-18, 2018
- Academic Workshop of China Society of Desert in the Geographical Society of China successfully held in Changsha, Hunan Province
- Local meteorology in a northern Himalayan valley near Mount Everest and its response to seasonal transitions
- Contrasting vegetation changes in dry and humid regions of the Tibetan Plateau over recent decades
- Simulation and prediction of monthly accumulated runoff,based on several neural network models under poor data availability
- Stable isotopes reveal varying water sources of Caragana microphylla in a desert-oasis ecotone near the Badain Jaran Desert