
土壤氮磷状况对小麦叶片养分生态化学计量特征的影响*
2019-01-03 03:13王凡坤付为国
中国生态农业学报(中英文) 2019年1期
王凡坤, 薛 珂, 付为国
土壤氮磷状况对小麦叶片养分生态化学计量特征的影响*
王凡坤, 薛 珂, 付为国**
(江苏大学现代农业装备与技术教育部重点实验室/江苏省农业装备与智能化高技术研究重点实验室 镇江 212013)
生态化学计量学是研究生态系统能量平衡和多重化学元素平衡的科学, 可利用植物体内C、N和P含量及其相互比值(N∶P、C∶N和C∶P, 即生态化学计量比)来判断植物生长过程中营养的供给状况。本文通过研究小麦叶片主要元素的生态化学计量比对不同土壤N、P状况的响应, 为小麦生产中的精确施肥提供技术指导。选取研究区当家品种弱筋小麦‘扬麦15’和强筋小麦‘镇麦168’为试验对象, 采用盆栽试验, 设置16个土壤N、P梯度处理, 分析不同处理下小麦拔节期、孕穗期和灌浆期叶片N∶P、C∶N和C∶P等生态化学计量比及其相互关系。结果表明: 1)在拔节期和孕穗期, 土壤N∶P范围在7.04~8.73时, 两小麦品种叶片的N∶P均达到较高水平; 而在灌浆期, 土壤N∶P范围在8.73~10.42时, 两小麦叶片的N∶P均达较高水平。且小麦叶片N∶P与土壤N∶P具有极显著的正相关关系, 但其相关性随着生育期的推进逐渐降低。2)当土壤处于低N水平时(108.4 mg∙kg-1), 两小麦品种叶片C∶N均达较高水平, 且其与土壤N∶P呈显著负相关, 即随着土壤N∶P比率增大, 小麦叶片中C∶N呈减小的趋势。3)当土壤处于低P水平时(29.6 mg∙kg-1), 两小麦品种叶片C∶P达较高水平, 且其与土壤P含量呈极显著负相关, 即随土壤中P的增加, 小麦叶片中C∶P均呈减小趋势。以上结果表明, 土壤中N和P的不同供应水平显著改变不同品种小麦叶片的生态化学计量比, 因此, 具有稳态特性的生态化学计量比可作为小麦生产中养分调控的重要参照指标并加以应用。
生态化学计量; 小麦叶片; 生育期; 土壤氮磷
生态化学计量学是研究生态系统能量平衡和多重化学元素平衡的科学, 是揭示分析多重化学元素平衡对生态交互作用影响的一种理论[1-3]。生态化学计量学目前主要强调活的有机体的主要组成元素之间的关系, 特别是C、N、P等重要生命元素间的计量关系[4]。该理论已发展了两个非常重要的具体理论, 即动态平衡理论(homeostasis hypothesis)和生长速率理论(growth rate hypothesis)。其中, 动态平衡理论指有机体中的元素组成与其周围环境(包括它们利用的资源)养分元素供应保持相对稳定的一种状态[5], 它是生态化学计量学存在的理论基础; 生长速率理论认为有机体必须改变其C∶N∶P比值, 从而使生长速率得以改变, 因此, 可利用植物体内C、N和P含量及其相互比值(N∶P、C∶N和C∶P), 作为判断植物生长对营养供给适应的指标[6-7]。其中, 单个有机体P的生物地球化学过程与生长和繁殖紧密相关(它是该种群动态的一个主要决定者)[1]。因为有机体C∶N∶P的变化主要由有机体P含量的变化决定[8]。
国外学者在生态化学计量学方面已经取得了大量研究成果。Koerselman等[9]在陆地生态系统中的研究结果表明, N和P是限制植物生长和发育的主要营养元素, N∶P是决定群落结构和功能的关键性指标。Güsewell等[10]发现当植物叶片N∶P<10时, 植物生长受N元素的限制, 当N∶P>20时, 植物生长受P元素限制, 而介于两者之间时, 植物生长受N、P两元素的共同限制。Niklas等[11]通过研究131种乔草分析得出, 叶片N含量与叶片C含量呈显著的等速生长, 叶片N含量与叶片P含量呈幂指数为3/4的异速生长。我国在生态化学计量学方面的研究起步较晚, 最近几年国内学者在生态化学计量学方面的发展取得了丰富的研究成果。但目前对该类研究主要集中在未经人工施肥的自然生态系统, 如对西双版纳热带雨林、塔里木河上游荒漠区、太白山、滨海盐地和滇池流域富磷区[12-16], 而在需要进行人工施肥的农业生产生态系统上的研究较少, 目前仅在园艺蔬菜上进行了相关研究, 如番茄(Mill.)和小青菜(L.)的生态化学计量特征研究[17-18]。而关于大田作物的研究则很少, 如不同施肥对塔里木河上游绿洲棉花(spp.)和黄泥田水稻(L.)生态化学计量比的影响[19-20]。本课题组也曾研究了不同施肥状况对不同品种小麦(L.)化学计量特征及其光合特性的影响[21]。前人的研究表明, 植物叶片元素含量会受海拔和干湿季节影响; 相同环境中不同植物的叶片化学计量特征并非由土壤养分含量特征直接决定的, 更多受植物自身遗传特性的影响。通过对不同施肥模式下园艺蔬菜的元素含量以及化学计量特征, 可优化施肥模式。施肥可补充土壤N、P养分, 但农田系统对外源养分需求具有差异性, 土壤有机碳、N和P含量与产量均呈极显著正相关, 土壤N、P养分与对应的植物N、P养分也呈显著正相关, 说明土壤C、N和P库总量供应水平与农田系统产量及植株养分累积具有协同性; 大田作物在生长期内会受到N、P元素的共同限制。本研究利用生态化学计量学方法, 研究不同土壤养分状态下不同品种小麦生态化学计量特征, 基于内稳态理论, 确定不同生长阶段小麦养分的最低适宜供给量; 基于生长速率理论, 确定不同生长阶段的小麦养分元素供给比例结构, 从而为小麦科学施肥提供技术指导。
1 材料和方法
1.1 供试材料
以弱筋小麦品种‘扬麦15’(低蛋白)和强筋小麦品种‘镇麦168’(高蛋白)为供试材料, 采用上底面直径24 cm、下底面直径18 cm、高20 cm的花盆进行盆栽试验, 栽培土取自江苏大学农业工程研究院作物栽培槽表土层(0~25 cm), 花盆土壤深度为16 cm。供试土壤基本理化性质为: 有机质11.2 g∙kg-1、速效氮108.4 mg∙kg-1、速效磷29.6 mg∙kg-1, pH为8.14。供试肥料为硝酸铵(NH4NO3)、磷酸二氢钠[NaH2(PO4)·2H2O]和硫酸钾(K2SO4)。土壤风干后进行分样和细磨, 过5 mm筛, 装瓶, 储存。
1.2 试验设计
以栽培槽表土为对照(表示为N0P0), 通过向盆土中添加硝酸铵和磷酸二氢钠的方式, 使土壤速效氮和速效磷浓度达到预设浓度。共设置16个氮磷组合处理, 分别表示为N0P0、N0P1、N0P2、N0P3、N1P0、N1P1、N1P2、N1P3、N2P0、N2P1、N2P2、N2P3、N3P0、N3P1、N3P2和N3P3。各处理土壤N、P化学计量特征见表1。每个处理3次重复, 盆栽后每盆代表1个处理的1次重复。每盆均施硫酸钾为底肥, 施用量以K土壤含量74 mg∙kg-1为标准, 将肥料按处理设计量与风干土壤充分混匀, 稳定一段时间后装盆待小麦播种。于2014年10月14日进行小麦播种, 每盆播种14粒, 3叶期后定苗, 每盆留苗7株, 露天栽培, 定期浇水使土壤保持湿润状态。
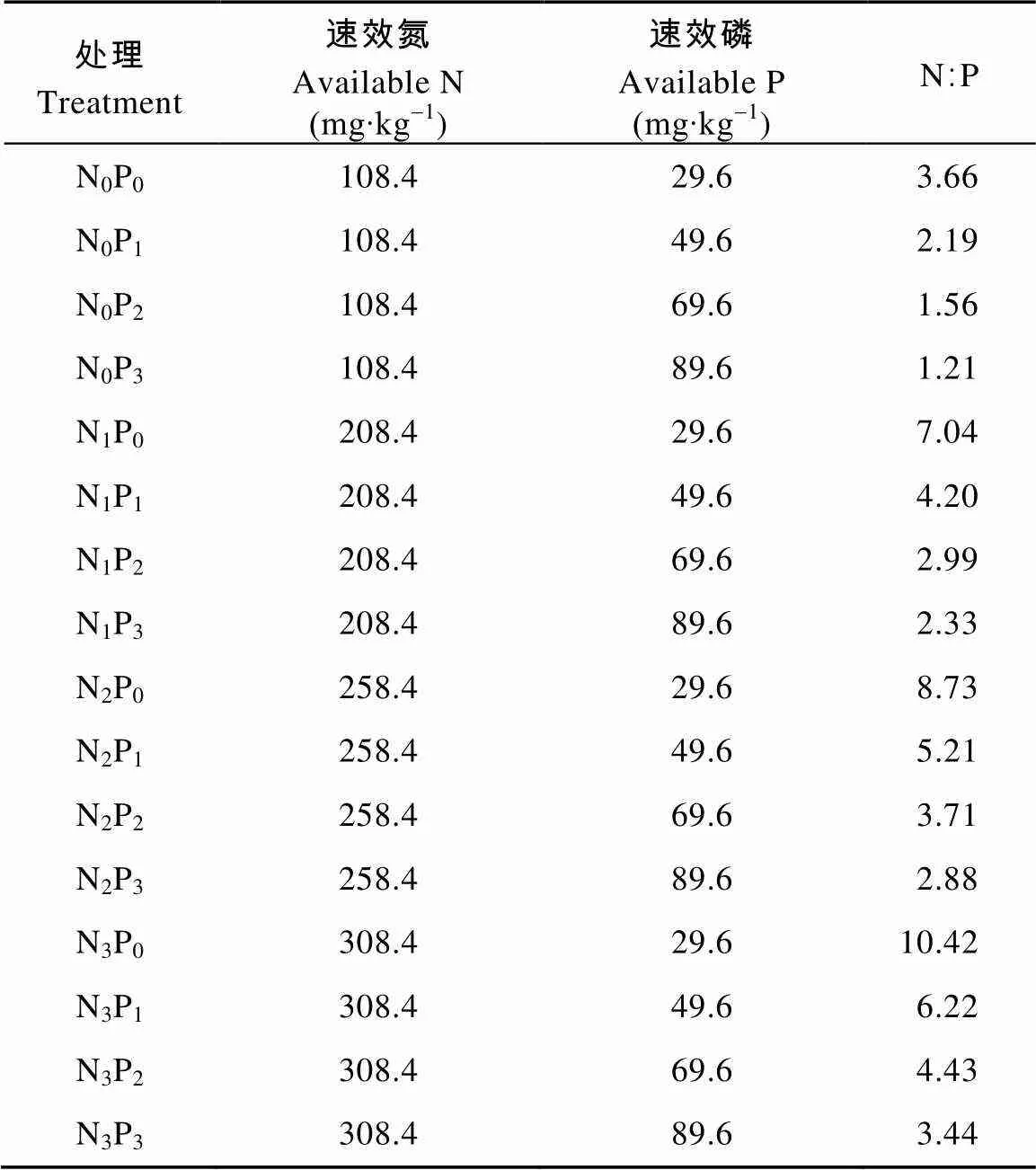
表1 试验各处理的土壤N、P含量及N、P化学计量特征
1.3 测定指标及方法
于2015年3月17日(拔节期)采摘顶端叶片, 4月2日(孕穗期)和4月23日(灌浆期)采摘旗叶叶片, 然后用去离子水冲洗以去除灰尘及泥沙等杂物。样品于105 ℃下杀青15 min, 70 ℃下烘干至恒重。同一样品的叶片混合粉碎后过100目筛制成供试样品。叶片全氮采用浓H2SO4消煮-半自动凯氏定氮仪法测定, 全磷采用NaOH熔融-钼锑抗比色法测定, 全碳含量采用K2Cr2O7氧化-分光光度法测定[22]。
1.4 数据处理和统计分析
对各处理水平的3次重复取平均值后进行统计分析, 在=0.05水平下进行Tukey多重比较(ANOVA)。
2 结果与分析
2.1 土壤N∶P对小麦叶片N∶P的影响
试验设计的土壤N∶P变化范围是1.2~10.4。由拔节期小麦叶片N∶P和土壤N∶P的关系(图1)可知, 拔节期‘扬麦15’顶端叶片N∶P的变化范围为4~18(变异系数为0.379 9), ‘镇麦168’顶端叶片N∶P的变化范围为7~20(变异系数为0.307 1)。拔节期‘扬麦15’和‘镇麦168’顶端叶片N∶P均极显著地高于相应的土壤N∶P(扬麦15=0.000 7,镇麦168<0.000 1)。通过计算可知, 拔节期两品种小麦顶端叶片N∶P与土壤N∶P均呈极显著正相关关系(扬麦15=0.758,<0.01;镇麦168=0.853,<0.01), 随土壤N∶P的增加叶片中N∶P也呈现出增加的态势。且两品种小麦顶端叶片N∶P与土壤N∶P的变化均不是等比例的, 两品种小麦顶端叶片N∶P关于土壤N∶P线性回归方程的斜率均大于1(‘扬麦15’为1.184 0, ‘镇麦168’为1.239 4), 即拔节期两品种小麦旗叶叶片N∶P的增加速率均大于土壤N∶P的增加速率。
由孕穗期小麦叶片N∶P和土壤N∶P的关系(图2)可知, 孕穗期‘扬麦15’旗叶叶片N∶P的变化范围为5~15(变异系数为0.269 1), ‘镇麦168’旗叶叶片N∶P的变化范围为8~15(变异系数为0.253 1), 孕穗期‘扬麦15’和‘镇麦168’旗叶叶片N:P均极显著地高于相应的土壤N∶P(扬麦15=0.001 2,镇麦168< 0.000 1)。通过计算可知, 孕穗期两品种小麦叶片N∶P与土壤N∶P均呈极显著正相关关系(扬麦15= 0.735**,镇麦168=0.877**), 随土壤N∶P的增加叶片N∶P也呈现出增加的态势。但孕穗期‘扬麦15’旗叶叶片N∶P的变化与土壤N∶P的变化不是等比例的, ‘扬麦15’旗叶叶片N∶P关于土壤N∶P线性回归方程的斜率为0.878 1, 即孕穗期‘扬麦15’旗叶叶片N∶P的增加速率小于土壤N:P的增加速率。而‘镇麦168’旗叶叶片N∶P的变化与土壤N∶P的变化是等比例的, ‘镇麦168’旗叶叶片N∶P关于土壤N∶P线性回归方程的斜率为1.037 4, 即孕穗期‘镇麦168’旗叶叶片N∶P的增加速率等于土壤N:P的增加速率。

图1 拔节期‘扬麦15’(a)和‘镇麦168’(b)叶片N∶P和土壤N∶P的关系
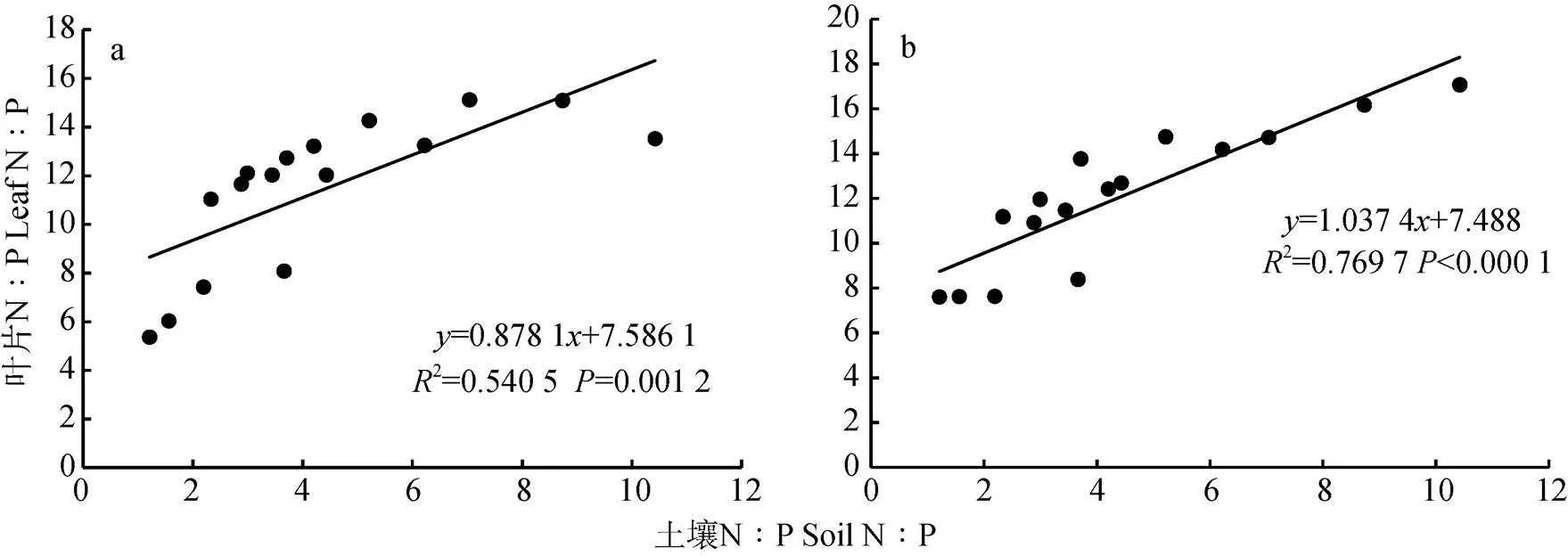
图2 孕穗期‘扬麦15’(a)和‘镇麦168’(b)叶片N∶P和土壤N∶P的关系
由灌浆期小麦叶片N∶P和土壤N∶P的关系(图3)可知, 灌浆期‘扬麦15’旗叶叶片N∶P的变化范围为3~10(变异系数为0.294 4), ‘镇麦168’旗叶叶片N∶P的变化范围为4~10(变异系数为0.270 7), 两品种小麦旗叶叶片N∶P均极显著地高于相应的土壤N∶P (扬麦15=0.000 5,镇麦168<0.000 1)(N3P0处理除外)。通过计算可知, 两品种小麦旗叶叶片N∶P与土壤N∶P均呈极显著正相关关系(扬麦15=0.770**,镇麦168=0.825**), 随土壤N∶P的增加叶片中N∶P也呈增加的态势。两品种小麦旗叶叶片N∶P的变化与土壤N∶P的变化均不是等比例的, 两品种小麦旗叶叶片N∶P关于土壤N∶P线性回归方程的斜率均小于1(‘扬麦15’为0.616 5, ‘镇麦168’为0.597 2), 即灌浆期两品种小麦旗叶叶片N∶P的增加速率均小于土壤N∶P的增加速率。

图3 灌浆期‘扬麦15’(a)和‘镇麦168’(b)叶片N∶P和土壤N∶P的关系
两品种小麦在拔节期—孕穗期—灌浆期这一生长发育进程中, 小麦顶端叶片N∶P的变异系数均呈先减少后增加的趋势, 且均为‘扬麦15’大于‘镇麦168’。说明在这一生长发育进程中, 两品种小麦旗叶叶片N∶P的变化幅度为孕穗期<灌浆期<拔节期。随土壤N∶P的增加叶片中N∶P也呈现出增加的态势, 并不是土壤中的速效氮和速效磷越多叶片中N∶P越大。并且两品种小麦顶端叶片N∶P关于土壤N∶P的线性回归方程的斜率均呈现持续减小的趋势, 说明两品种小麦顶端叶片N∶P的变化随土壤N∶P变化的影响减小。
2.2 土壤N∶P对小麦叶片C∶N的影响
由拔节期小麦叶片C∶N和土壤N∶P的关系(图4)可知, 拔节期‘扬麦15’顶端叶片C∶N的变化范围为8~21(变异系数为0.510 9), ‘镇麦168’叶片C∶N的变化范围为6~13(变异系数为0.317 3), 各处理间‘扬麦15’顶端叶片C∶N均高于‘镇麦168’。通过计算可知, 拔节期‘扬麦15’和‘镇麦168’顶端叶片C∶N与土壤N∶P均呈显著负相关关系(扬麦15=-0.510*,镇麦168=-0.536*)。拔节期两品种小麦顶端叶片C∶N的变化随土壤N∶P的变化均不是等比例的。N0水平下‘扬麦15’顶端叶片C∶N关于土壤N∶P线性回归方程的斜率为0.128 6, 即‘扬麦15’顶端叶片C∶N的增加速率小于土壤N∶P的增加速率, 随土壤N∶P的增加, 叶片C∶N呈小幅增加趋势; N1、N2和N3水平下‘扬麦15’顶端叶片C∶N关于土壤N∶P的线性回归方程的斜率为0.012 3, 表明在N1、N2和N3水平下‘扬麦15’顶端叶片C∶N随土壤N∶P的增加在小范围内保持相对稳定, 在相同N处理水平下叶片C∶N基本保持相对稳定。N0水平下‘镇麦168’顶端叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.067 2, 随土壤N∶P的增加, 叶片C∶N呈小幅减小的趋势; N1、N2和N3水平下‘镇麦168’顶端叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.035 7, 表明‘镇麦168’顶端叶片C∶N随土壤N∶P的增加呈小幅减小的趋势, 在相同N处理水平下保持相对稳定。

图4 不同土壤速效氮含量下拔节期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶N和土壤N∶P的关系
N0处理下土壤速效氮含量为108.4 mg∙kg-1; N1、N2、N3处理下土壤速效氮含量分别为208.4 mg∙kg-1、258.4 mg∙kg-1和308.4 mg∙kg-1。Under N0, N1, N2and N3treatments, soil available nitrogen content is 108.4 mg∙kg-1, 208.4 mg∙kg-1, 258.4 mg∙kg-1and 308.4 mg∙kg-1, respectively.
由孕穗期小麦叶片C∶N和土壤N∶P的关系(图5)可知, 孕穗期‘扬麦15’旗叶叶片C∶N的变化范围为8~21(变异系数为0.420 3), ‘镇麦168’旗叶叶片C∶N的变化范围为9~19(变异系数为0.350 6)。N0水平下‘扬麦15’旗叶叶片C∶N高于‘镇麦168’, 而N1、N2和N3水平下‘扬麦15’则低于‘镇麦168’。通过计算可知, 孕穗期两品种小麦旗叶叶片C∶N与土壤N∶P均呈显著负相关关系(扬麦15=-0.533*,镇麦168=-0.557*)。N0水平下孕穗期‘扬麦15’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为0.118 1, 即‘扬麦15’旗叶叶片C∶N的增加速率小于土壤N∶P的增加速率; 随土壤N∶P的增加, 叶片C∶N呈现出小幅度增加的趋势。N1、N2和N3水平下‘扬麦15’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.062 5, 表明N1、N2和N3水平下‘扬麦15’旗叶叶片C∶N随土壤N∶P的增加呈小幅减小的趋势, 但在小范围内保持相对稳定。N0水平下孕穗期‘镇麦168’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.185 5, 即‘镇麦168’旗叶叶片C∶N的减少速率小于土壤N∶P的增加速率, 随土壤N∶P的增加, 叶片C∶N呈小幅减小的趋势; N1、N2和N3水平下‘镇麦168’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.103 8, 表明‘镇麦168’旗叶叶片C∶N随土壤N∶P的增加呈小幅减小的趋势, 在相同N处理水平下保持相对稳定。
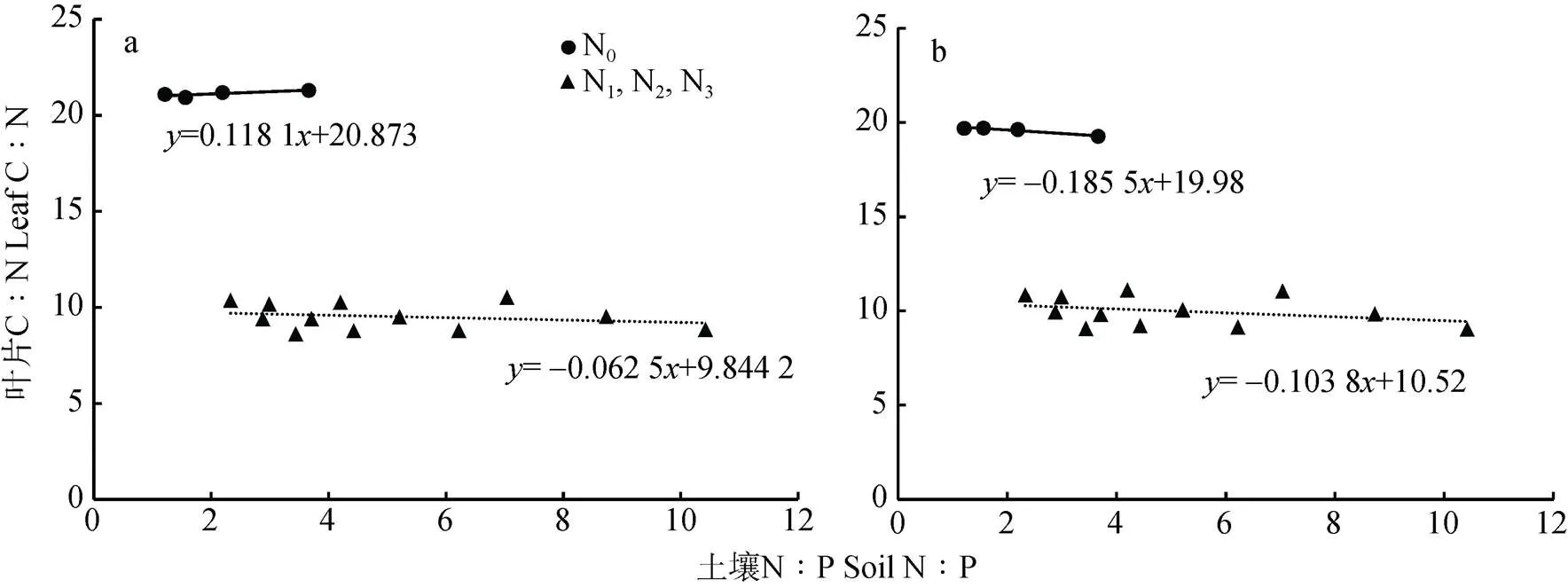
图5 不同土壤速效氮含量下孕穗期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶N和土壤N∶P的关系
N0处理下土壤速效氮含量为108.4 mg∙kg-1; N1、N2、N3处理下土壤速效氮含量分别为208.4 mg∙kg-1、258.4 mg∙kg-1和308.4 mg∙kg-1。Under N0, N1, N2and N3treatments, soil available nitrogen content is 108.4 mg∙kg-1, 208.4 mg∙kg-1, 258.4 mg∙kg-1and 308.4 mg∙kg-1, respectively.
由灌浆期小麦叶片C∶N和土壤N∶P的关系(图6)可知, 灌浆期‘扬麦15’旗叶叶片C∶N的变化范围为9~25(变异系数为0.461 5), ‘镇麦168’旗叶叶片C∶N的变化范围为9~23(变异系数为0.386 1)。N0水平下‘扬麦15’旗叶叶片C∶N高于‘镇麦168’, 而N1、N2和N3水平下‘扬麦15’则低于‘镇麦168’。通过计算可知, 灌浆期两品种小麦旗叶叶片C∶N与土壤N∶P均呈显著负相关关系(扬麦15=-0.539*,镇麦168=-0.567*)。灌浆期两品种小麦旗叶叶片C∶N的变化与土壤N∶P的变化均不是等比例的, N0水平‘扬麦15’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为0.211 3, 即‘扬麦15’旗叶叶片C∶N的增加速率均小于土壤N∶P的增加速率, 随土壤N∶P的增加, 叶片C∶N呈小幅增加的趋势; N1、N2和N3水平下‘扬麦15’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.107 9, 表明N1、N2和N3水平下‘扬麦15’旗叶叶片C∶N的减少速率小于土壤N∶P的增加速率, 随土壤N∶P的增加, 叶片C∶N呈小幅减小的趋势。N0水平‘镇麦168’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.045 7, 表明‘镇麦168’旗叶叶片C∶N在N0水平下保持相对稳定; N1、N2和N3水平下‘镇麦168’旗叶叶片C∶N关于土壤N∶P线性回归方程的斜率为-0.185 1, 表明N1、N2和N3水平下‘镇麦168’旗叶叶片C∶N的减少速率小于土壤N∶P的增加速率, 随土壤N∶P的增加, 叶片C∶N呈小幅减小的趋势。
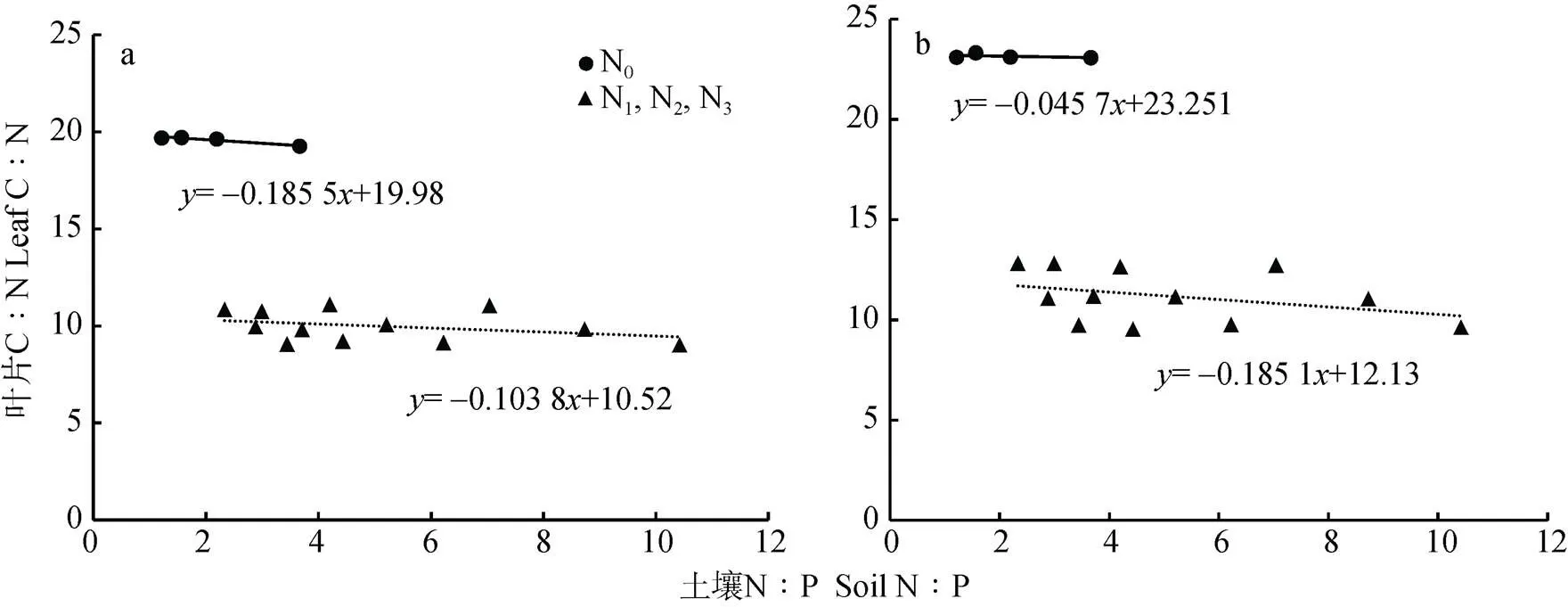
图6 不同土壤速效氮含量下灌浆期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶N和土壤N∶P的关系
N0处理下土壤速效氮含量为108.4 mg∙kg-1; N1、N2、N3处理下土壤速效氮含量分别为208.4 mg∙kg-1、258.4 mg∙kg-1和308.4 mg∙kg-1. Under N0, N1, N2and N3treatments, soil available nitrogen content is 108.4 mg∙kg-1, 208.4 mg∙kg-1, 258.4 mg∙kg-1and 308.4 mg∙kg-1, respectively.
在两品种小麦由拔节期—孕穗期—灌浆期这一生长发育进程中, ‘扬麦15’顶端叶片C∶N的变异系数呈先减少后增加的变化趋势, ‘镇麦168’则呈持续增加的趋势, 且均为‘扬麦15’大于‘镇麦168’。说明在这一生长发育进程中, 小麦叶片C∶N的变化幅度为‘扬麦15’>‘镇麦168’; ‘扬麦15’叶片C∶N的变化幅度为孕穗期<灌浆期<拔节期; ‘镇麦168’则为拔节期<孕穗期<灌浆期。两品种在各个生长发育期, 叶片的C∶N受土壤N∶P变化影响较小, 在相同N处理水平下, 叶片的C∶N在小范围内维持稳定; 随土壤N含量的改变, 不同N梯度间叶片C∶N有较为明显的改变。
2.3 土壤P含量对小麦叶片C∶P的影响
试验设计的土壤P变化范围为30~90 mg∙kg-1。由拔节期小麦叶片C∶P和土壤P含量的关系(图7)可知, 拔节期‘扬麦15’顶端叶片C∶P的变化范围为74~175(变异系数为0.272 2), ‘镇麦168’顶端叶片C∶P的变化范围为74~152(变异系数为0.251 7)。通过计算可知, 拔节期两品种小麦顶端叶片C∶P与土壤P含量呈极显著负相关关系(扬麦15=-0.758**,镇麦168=-0.831**)。拔节期两品种小麦顶端叶片C∶P的变化与土壤P含量的变化均不是等比例的, 两品种小麦顶端叶片C∶P关于土壤P线性回归方程的斜率均大于-1(‘扬麦15’为-0.919 0, ‘镇麦168’为-0.911 4), 即拔节期两品种小麦随土壤P含量的增加叶片C∶P呈减小的趋势。
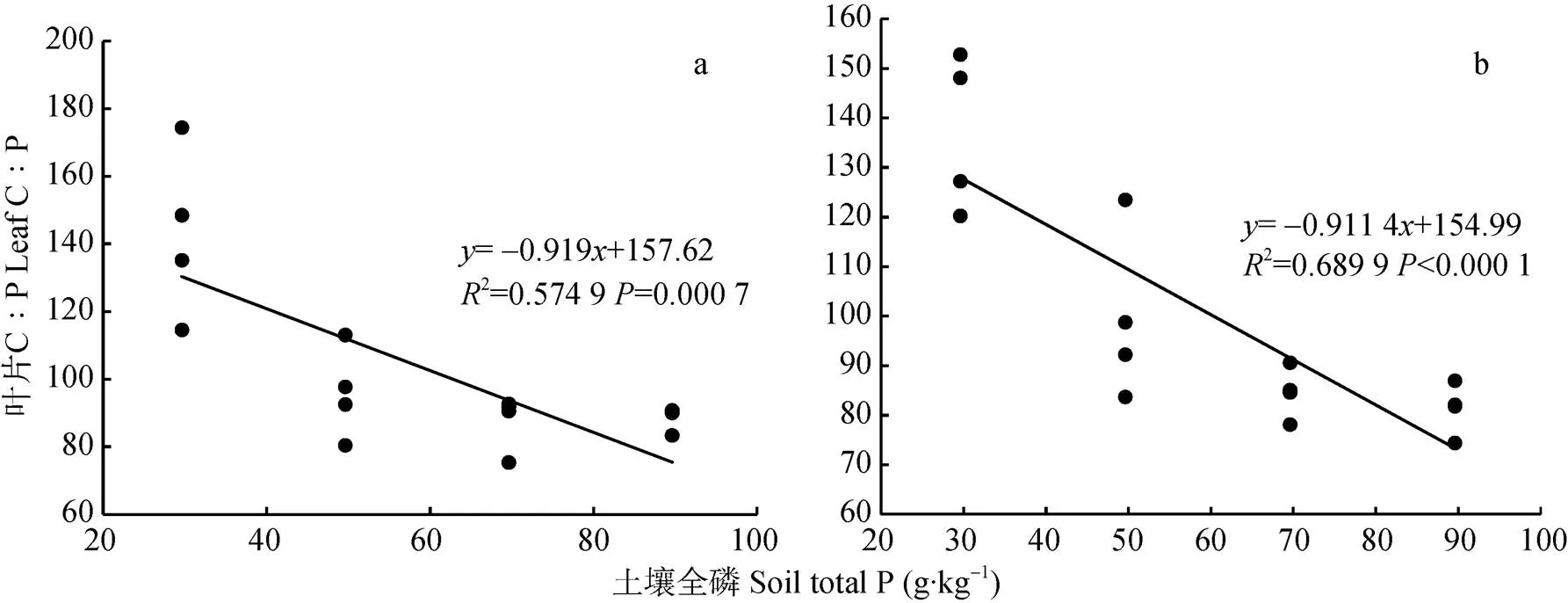
图7 拔节期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶P和土壤P含量的关系
由孕穗期小麦叶片C∶P和土壤P含量的关系(图8)可知, 孕穗期‘扬麦15’旗叶叶片C∶P的变化范围为103~172(变异系数为0.158 9), ‘镇麦168’旗叶叶片C∶P的变化范围为104~163(变异系数为0.135 5)。通过计算可知, 孕穗期两品种小麦旗叶叶片C∶P与土壤P含量呈极显著负相关关系(扬麦15=-0.751**,镇麦168=-0.760**)。孕穗期两品种小麦旗叶叶片C∶P的变化与土壤P含量的变化均不是等比例的, 两品种小麦旗叶叶片C∶P关于土壤P线性回归方程的斜率均大于-1(‘扬麦15’为-0.664 7, ‘镇麦168’为-0.618 4), 即孕穗期两品种小麦随土壤P含量的增加叶片C∶P呈减小的趋势。
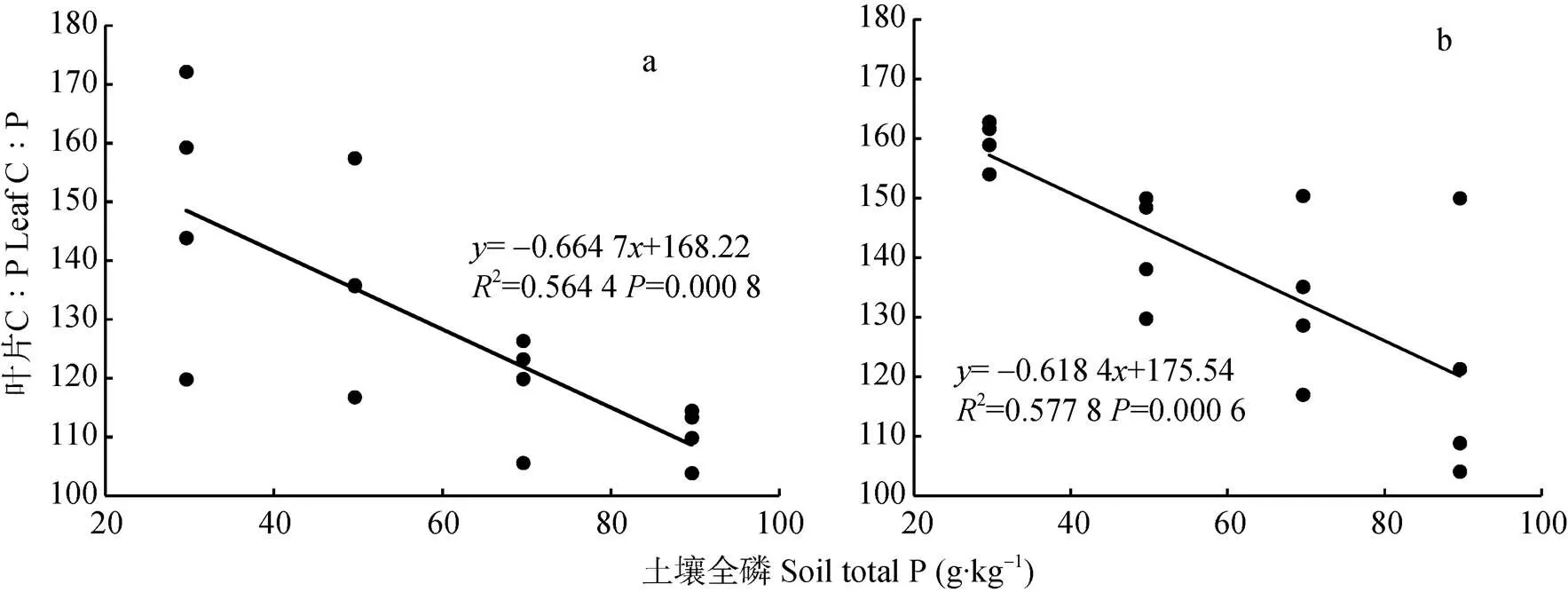
图8 孕穗期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶P和土壤P含量的关系
由灌浆期小麦叶片C∶P和土壤P含量的关系(图9)可知, 灌浆期‘扬麦15’旗叶叶片C∶P的变化范围为69~105(变异系数为0.125 3), ‘镇麦168’旗叶叶片C∶P的变化范围为75~108(变异系数为0.120 1)。通过计算可知, 灌浆期两品种小麦旗叶叶片C∶P与土壤P含量呈极显著负相关关系(扬麦15=-0.797**,镇麦168=-0.806**)。灌浆期两品种小麦旗叶叶片C∶P的变化与土壤P含量的变化均不是等比例的, 两品种小麦旗叶叶片C∶P关于土壤P线性回归方程的斜率均大于-1(‘扬麦15’为-0.365 4, ‘镇麦168’为-0.372 7), 即灌浆期两品种小麦随土壤P含量的增加叶片C∶P呈减小的趋势,。
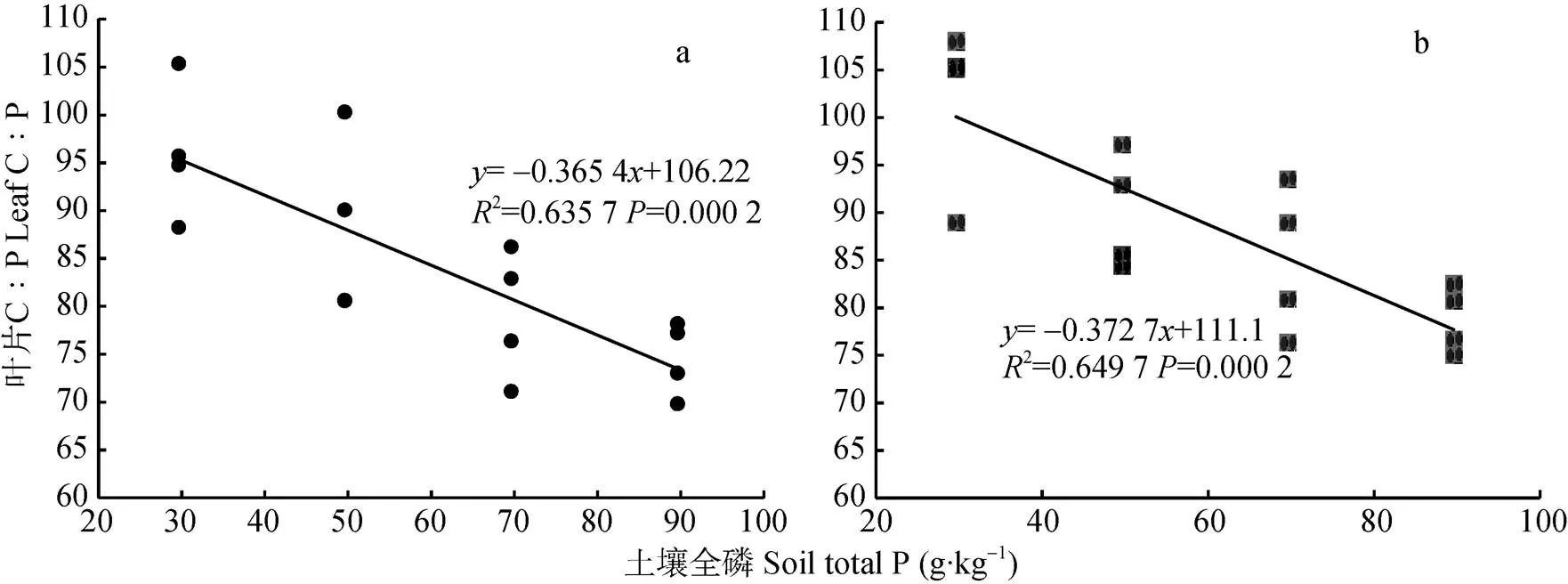
图9 灌浆期‘扬麦15’(a)和‘镇麦168’(b)叶片C∶P和土壤P含量的关系
两品种小麦在拔节期—孕穗期—灌浆期这一生长发育进程中, 小麦顶端叶片C∶P的变异系数均呈持续减少的趋势, 且均为‘扬麦15’大于‘镇麦168’。说明在这一生长发育进程中, 两品种小麦叶片生态化学计量C∶P比越来越稳定。在拔节期到孕穗期这一生长发育进程中, 两品种小麦旗叶叶片C∶P关于土壤P含量的线性回归方程的斜率均呈持续增加的趋势。说明两品种小麦顶端叶片C∶P值的变化随土壤P含量的变化的影响减小。
3 讨论
作物叶片各元素含量及其相互比例虽然与自身生理结构和物质合成有关, 但同时它对土壤养分环境具有积极响应特性[23]。有学者表示, 通过环境条件改变, 作物通常具有调节自身元素化学计量特征的能力[24]。从作物施肥总量角度来看, 根据内稳态理论, 可通过控制施肥总量, 找到作物施肥的最低下限; 根据异速生长理论可知, 环境的改变会导致作物生态化学计量比的改变, 作物在不同生长阶段需要不同的养分供应结构, 将之应用在农业水肥调控上, 通过调整养分结构, 来满足作物养分的需求, 对农业水肥调控有指导性意义。因此, 下一步可开展实际的应用研究, 通过大田试验, 研究实际生产过程中小麦对氮磷营养元素的需求。
3.1 土壤不同N∶P处理对小麦叶片N∶P的影响
土壤中可以被植物利用的养分含量对植物叶片的元素组成及含量具有直接的影响[25]。植物体内的N、P在吸收和利用上相互依赖[26]。一般情况土壤中添加N使土壤磷酸酶活性增加, 从而加速植物对土壤中P的吸收[27-29]。
通过施加不同比例的化肥, 土壤中N、P含量相应增加, 小麦对N、P吸收量随之增强, 小麦叶片中N、P浓度增大, 导致C∶N和C∶P降低; 但对N∶P而言, 虽然施肥会补充土壤N、P养分, 但小麦对外界补充N、P需求产生差异。叶片的N∶P能较好地反映生境的养分状况对植物生长的限制性, 学者们也普遍认为, 当叶片N∶P低时, 意味着植物生长受N限制, 当N∶P高, 则表示受P限制。在土壤N、P供应充足的情况下, 植物叶片N∶P会在一定的范围内稳定[30]。严正兵等[31]研究了不同氮磷施肥方案对拟南芥()叶片C、N和P化学计量特征的影响, 认为拟南芥叶片N∶P和C∶P的增加, 叶片N含量与基质N含量之间无显著相关, 叶片N和P含量具有显著的异速生长关系。顾大形等[32]通过土壤添加不同浓度N、P研究其对四季竹()叶片N、P化学计量特征的影响, 结果为叶片N∶P随土壤N∶P的增大而增大, 但其增加速率小于土壤N∶P的增加速率, 土壤全氮较土壤全磷对四季竹叶片N、P化学计量特征具有更大的影响。本试验中不同生育期两品种小麦叶片N∶P与土壤N∶P具有极显著的正相关关系, 随土壤N∶P的增加, 小麦叶片N∶P也呈增加的趋势。但随着生育期的推进, 其相关性逐渐降低。说明小麦叶片N∶P在一定范围内具有可塑性, 对土壤N、P的营养平衡具有较强的适应能力。值得注意的是, 小麦叶片N∶P随着土壤N含量的提高较N0处理有显著提高(<0.05)。这可能是因为当土壤N供应充足时, 小麦叶片对N元素存在奢侈吸收[33]。即小麦叶片能储存超过其需要的养分量, 以供应灌浆期籽粒对N元素的大量而迅速的需求, 进而灵活地调整其N库储量, 以适应生活史(特别是灌浆期)和周围环境的动态变化[34]。
在本试验中, 小麦在拔节期和孕穗期, 土壤N∶P范围在7.04~8.73时, 两小麦叶片的N∶P均达到较高水平; 而在灌浆期, 土壤N∶P范围在8.73~10.42时, 两小麦叶片的N∶P均达到较高水平。土壤较优N∶P范围为7.04~10.42, 符合该比例的土壤N含量分别为208.9 mg∙kg-1、258.4 mg∙kg-1和308.4 mg∙kg-1, P含量均为29.6 mg∙kg-1。由此可得, 在小麦不同生长期中, 保持较低P含量, 适量添加N有助于作物生长。
3.2 土壤不同N∶P处理对小麦叶片C∶N的影响
C、N作为有机物的基本组成元素, 在生态系统中构成庞大且复杂的C、N循环。小麦叶片C∶N也能反映出小麦对土壤中N、P的利用效率[35]。C、N代谢作为小麦生长发育最基本的生理代谢, 是反映小麦生理状况、生长活力及抗病能力的重要指标[36]。C是组成植物体的结构性物质, N、P则为功能性物质, 功能性物质的含量受环境影响变化较大, 结构性物质受环境的影响较小[37]。本试验结果变相印证了这一点, 不同生育期两品种小麦叶片C∶N与土壤N∶P具有显著的负相关关系, 这主要是因为小麦叶片N含量随土壤N∶P的增加而减少, 低的土壤N∶P引起小麦叶片N含量增加, 而不是小麦叶片C含量减少。由此可见, 小麦叶片N素并不是随土壤N素增加而等量增加, 而是随土壤中的养分组成变化而在小范围内保持相对稳定, 这一现象与生态化学计量学的内稳态理论相符合。在小麦的生长发育进程中, 不仅要保持充足的C、N营养, 而且要求C∶N适当。朱新开等[38]的研究结果表明, 在小麦孕穗开花期, 小麦植株体内C∶N处于高峰, 小麦生育中后期是籽粒形成的关键时期, 而较高的C∶N特别是茎鞘高的C∶N, 能够促进小麦穗、粒、重协调发展, 提高小麦产量。
在本试验中, 当土壤处于低N水平时(108.4 mg∙kg-1), 两小麦叶片在不同生长时期C∶N均达到较高水平, 符合该条件的土壤N含量为108.4 mg∙kg-1, 其他水平下叶片C∶N随土壤N∶P变化的差异较小。
3.3 土壤P对小麦叶片C∶P的影响
P在土壤中的有效性较低, 因此会成为大多生态系统的主要限制因子[39]。P不但是植物体中重要化合物的成分, 而且以多种形式参与植物的新陈代谢过程。植物快速生长时, 植物细胞会通过rRNA快速合成蛋白质, 从而使分配到rRNA的P增加[40]。当土壤P素处于一定浓度范围内, 随其浓度的增加, 植物会加快对P的吸收以保证蛋白质的合成。本试验得出在不同土壤N、P状况下, 不同生长时期的两品种小麦叶片C∶P均与土壤P含量具有极显著负相关关系, 随土壤中P的增加, 小麦叶片中C∶P均呈减小的趋势。小麦叶片C∶P随土壤P含量的增加而减少主要是由于小麦叶片P含量随土壤P含量的增加而增加的缘故, 而不是小麦叶片C含量减少的结果。由此可知, 小麦C∶P降低是由于受到外界P素的限制。小麦叶片P素并不是随土壤P素增加而等量增加, 而是随土壤中的养分组成变化而在小范围内保持相对稳定, 这一现象与生态化学计量学的内稳态理论相符合。
在本试验中, 当土壤处于低P水平时(29.6 mg∙kg-1), 两小麦叶片C∶P达到较高水平。在小麦不同生长期中, 保持较低P含量、适量添加N有助于作物生长。
4 结论
通过盆栽试验研究了不同土壤N、P化学计量特征对强筋小麦‘镇麦168’和弱筋小麦‘扬麦15’叶片C、N、P化学计量特征的影响。通过对两品种小麦不同生长阶段施肥, 明显提高了土壤N素、P素含量及小麦叶片N、P养分含量, 说明土壤中N、P总供应量水平是影响小麦叶片化学元素的重要指标。
1)不同生育期两品种小麦叶片N∶P与土壤N∶P具有极显著的正相关关系。在拔节期和孕穗期, 土壤N∶P范围在7.04~8.73时, 两小麦叶片的N∶P均达到较高水平; 而在灌浆期, 土壤N∶P范围在8.73~10.42时, 两小麦叶片的N∶P均达到较高水平。随土壤N∶P的增加, 小麦叶片N∶P也呈增加的趋势。但随着生育期的推进, 其相关性逐渐降低。
2)不同生育期两品种小麦叶片C∶N与土壤N∶P具有显著的负相关关系。当土壤处于低N水平时(108.4 mg∙kg-1), 两小麦叶片C∶N均达到较高水平, 当土壤处于较高N水平时(208.4 mg∙kg-1、258.4 mg∙kg-1、308.4 mg∙kg-1), 两品种小麦叶片C∶N均为较低水平。‘扬麦15’在整个生育期, 高N水平下叶片C∶N随土壤N∶P的增加呈现出增加的趋势, 低N水平下叶片C∶N随土壤N∶P的增加呈现出减少的趋势。在‘镇麦168’在整个生育期叶片C∶N均随土壤N∶P的增加呈减少的趋势。
3)不同生育期两品种小麦叶片C∶P与土壤P含量具有极显著负相关关系。当土壤处于低P水平时(29.6 mg∙kg-1), 两小麦叶片C∶P达到较高水平。随土壤中P的增加, 小麦叶片C∶P均呈减小的趋势。从而显示出各不同施肥模式下小麦不同程度受到了P素的限制, 而N素供应相对丰富。
4)不同品种小麦生长过程中, 各自叶片N∶P、C∶N和C∶P随土壤N、P变化均表现出较为一致的变化, 表明即使如‘镇麦168’和‘扬麦15’两种遗传特性差异较大的品种, 其生态化学计量亦对土壤N、P状况具有较为一致的响应特征, 这将便于人们利用生态化学计量特征对某一作物的不同品种施肥进行统一调控。
本试验只研究了土壤N、P添加对小麦旗叶叶片C、N、P化学计量的影响, 而影响小麦品质及产量的营养元素不仅仅是N和P, 众多研究结果表明, 土壤K、S及其他微量元素也对小麦产量形成和品质性状具有重要的影响, 这些研究内容将有待今后进一步开展。
[1] ELSER J J, DOBBERFUHL D R, MACKAY N A, et al. Organism size, life history, and N∶P stoichiometry: Toward a unified view of cellular and ecosystem processes[J]. Bioscience, 1996, 46(9): 674–684
[2] HESSEN D O. Stoichiometry in food webs: Lotka revisited[J]. Oikos, 1997, 79(1): 195–200
[3] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索[J]. 植物生态学报, 2005, 29(6): 1007–1019ZENG D H, CHEN G S. Ecological stoichiometry: A science to explore the complexity of living systems[J]. Acta Phytoecologica Sinica, 2005, 29(6): 1007–1019
[4] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论[J]. 植物生态学报, 2010, 34(1): 2–6 HE J S, HAN X G. Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems[J]. Chinese Journal of Plant Ecology, 2010, 34(1): 2–6
[5] ÅGREN G I. The C∶N∶P stoichiometry of autotrophs — theory and observations[J]. Ecology, 2004, 7(3): 185–191
[6] WASSEN M J, OLDE VENTERINK H G M, DE SWART E O A M. Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems[J]. Journal of Vegetation Science, 1995, 6(1): 5–16
[7] GÜSEWELL S. N∶P ratios in terrestrial plants: Variation and functional significance[J]. New Phytologist, 2004, 164(2): 243–266
[8] VANNI M J, FLECKER A S, HOOD J M, et al. Stoichiometry of nutrient recycling by vertebrates in a tropical stream: Linking species identity and ecosystem processes[J]. Ecology Letters, 2002, 5(2): 285–293
[9] KOERSELMAN W, MEULEMAN A F M. The vegetation N∶P ratio: A new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33(6): 1441–1450
[10] GÜSEWELL S, KOERSELMAN W, VERHOEVEN J T A. Biomass N∶P ratios as indicators of nutrient limitation for plant populations in wetlands[J]. Ecological Applications, 2003, 13(2): 372–384
[11] NIKLAS K J, COBB E D. N, P, and C stoichiometry of(Ranunculaceae) and the allometry of plant growth[J]. American Journal of Botany, 2005, 92(8): 1256–1263
[12] 卢同平, 王艳飞, 王黎明, 等. 西双版纳热带雨林土壤与叶片生态化学计量特征的干湿度效应[J]. 生态学报, 2018, 38(7): 2333–2343 LU T P, WANG Y F, WANG L M, et al. Effect of the humidity/aridity gradient on the ecological stoichiometry of soil and leaves in Xishuangbanna tropical rainforest[J]. Acta Ecologica Sinica, 2018, 38(7): 2333–2343
[13] 罗艳, 贡璐, 朱美玲, 等. 塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征[J]. 生态学报, 2017, 37(24): 8326–8335 LUO Y, GONG L, ZHU M L, et al. Stoichiometry characteristics of leaves and soil of four shrubs in the upper reaches of the Tarim River Desert[J]. Acta Ecologica Sinica, 2017, 37(24): 8326–8335
[14] 李丹维, 王紫泉, 田海霞, 等. 太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征[J]. 土壤学报, 2017, 54(1): 160–170 LI D W, WANG Z Q, TIAN H X, et al. Carbon, nitrogen and phosphorus contents in soils on Taibai Mountain and their ecological stoichiometry relative to elevation[J]. Acta Pedologica Sinica, 2017, 54(1): 160–170
[15] 李征, 韩琳, 刘玉虹, 等. 滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征[J]. 植物生态学报, 2012, 36(10): 1054–1061 LI Z, HAN L, LIU Y H, et al. C, N and P stoichiometric characteristics in leaves ofduring different growth phase in coastal wetlands of China[J]. Chinese Journal of Plant Ecology, 2012, 36(10): 1054–1061
[16] 阎凯, 付登高, 何峰, 等. 滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征[J]. 植物生态学报, 2011, 35(4): 353–361 YAN K, FU D G, HE F, et al. Leaf nutrient stoichiometry of plants in the phosphorus-enriched soils of the Lake Dianchi watershed, southwestern China[J]. Chinese Journal of Plant Ecology, 2011, 35(4): 353–361
[17] 袁伟, 董元华, 王辉. 氨基酸态氮和硝态氮混合营养下番茄生长及其生态化学计量学特征[J]. 土壤, 2010, 42(4): 664–668 YUAN W, DONG Y H, WANG H. Growth of tomato and its ecological stoichiometry characteristics under hydroponic condition with mixture nitrogen nutrition of amino acid and nitrate[J]. Soils, 2010, 42(4): 664–668
[18] 袁伟, 董元华, 王辉. 菜园土壤不同施肥模式下小青菜生长和品质及其生态化学计量学特征[J]. 土壤, 2010, 42(6): 987–992 YUAN W, DONG Y H, WANG H. Growth and qualities of greengrocery and its ecological stoichiometry characteristics under different fertilization[J]. Soils, 2010, 42(6): 987–992
[19] 贡璐, 李红林, 刘雨桐, 等. N、P施肥对塔里木河上游绿洲棉花C、N、P生态化学计量特征的影响[J]. 生态学报, 2017, 37(22): 7689–7697 GONG L, LI H Y, LIU Y T, et al. Effects of nitrogen and phosphorus fertilizers on carbon, nitrogen, and phosphorus stoichiometry of oasis cotton in the upper reaches of Tarim River, Xinjiang, China[J]. Acta Ecologica Sinica, 2017, 37(22): 7689–7697
[20] 王飞, 林诚, 李清华, 等. 长期不同施肥下黄泥田土壤-水稻碳氮磷生态化学计量学特征[J]. 土壤通报, 2017, 48(1): 169–176 WANG F, LIN C, LI Q H, et al. Stoichiometry of carbon, nitrogen and phosphorus in soil and plant under long-term different fertilizations in a yellow paddy field[J]. Chinese Journal of Soil Science, 2017, 48(1): 169–176
[21] 付为国, 王凡坤, 赵云, 等. 土壤氮磷化学计量特征对小麦光合气体交换参数和叶绿素荧光参数的影响[J]. 西北植物学报, 2016, 36(7): 1435–1442 FU W G, WANG F K, ZHAO Y, et al. Effects of soil nitrogen and phosphorus stoichiometric characteristics on photosynthetic gas exchange and chlorophyll fluorescence of wheat[J]. Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(7): 1435–1442
[22] 鲍士旦. 土壤农化分析[M]. 第3版. 北京: 中国农业出版社, 2000 BAO S D. Soil and Agricultural Chemistry Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000
[23] 邬畏, 何兴东, 周启星. 生态系统氮磷比化学计量特征研究进展[J]. 中国沙漠, 2010, 30(2): 296–302 WU W, HE X D, ZHOU Q X. Review on N∶P stoichiometry in eco-system[J]. Journal of Desert Research, 2010, 30(2): 296–302
[24] 朴河春, 刘丛强, 朱书法, 等. 贵州石灰岩和砂岩地区C4和C3植物营养元素的化学计量对N/P比值波动的影响[J]. 第四纪研究, 2005, 25(5): 552–560 PIAO H C, LIU C Q, ZHU S F, et al. Variations of C4and C3plant N∶P ratios influenced by nutrient stoichiometry in limestone and sandstone areas of Guizhou[J]. Quaternary Sciences, 2005, 25(5): 552–560
[25] 杨晓晖, 王葆芳, 江泽平. 乌兰布和沙漠东北缘三种豆科绿肥植物生物量和养分含量及其对土壤肥力的影响[J]. 生态学杂志, 2005, 24(10): 1134–1138 YANG X H, WANG B F, JIANG Z P. Biomass and nutrient concentrations of three green manure leguminous plants and their effects on soil nutrients at northeastern edge of Ulanbuhe Desert[J]. Chinese Journal of Ecology, 2005, 24(10): 1134–1138
[26] MARSCHNER H. Mineral Nutrition of Higher Plants[M]. 2nd ed. San Diego, CA: Academic Press, 2002
[27] ÅGREN G I, WETTERSTEDT J Å M, BILLBERGER M F K. Nutrient limitation on terrestrial plant growth — modeling the interaction between nitrogen and phosphorus[J]. New Phytologist, 2012, 194(4): 953–960
[28] FUJITA Y, ROBROEK B J M, DE RUITER P C, et al. Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity[J]. Oikos, 2010, 119(10): 1665–1673
[29] MARKLEIN A R, HOULTON B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems[J]. New Phytologist, 2012, 193(3): 696–704
[30] MCGRODDY M E, DAUFRESNE T, HEDIN L O. Scaling of C∶N∶P stoichiometry in forests world wild: Implications of terrestrial Redfield-type ratios[J]. Ecology, 2004, 85(9): 2390–2401
[31] 严正兵, 金南瑛, 韩延伸, 等. 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响[J]. 植物生态学报, 2013, 37(6): 551–557 YAN Z B, JIN N Y, HAN T S, et al. Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of[J]. Chinese Journal of Plant Ecology, 2013, 37(6): 551–557
[32] 顾大形, 陈双林, 黄玉清. 土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响[J]. 植物生态学报, 2011, 35(12):1219-1225. GU D X, CHEN S L, HUANG Y Q. Effects of soil nitrogen and phosphonium on leaf nitrogen and phosphonium stoichiometric characteristics and chlorophyll content of Oligostachyum lubricum[J]. Chinese Journal of Plant Ecology, 2007, 15(5): 377–382
[33] 周桦, 宇万太, 马强, 等. 追施氮肥对尾巨桉叶片主要营养元素浓度及贮量变化的影响[J]. 生态学杂志, 2010, 29(8): 1488–1492 ZHOU H, YU W T, MA Q, et al. Effects of topdressing nitrogen fertilization on the concentration and storage of major nutrients in eucalyptus leaves[J]. Chinese Journal of Ecology, 2010, 29(8): 1488–1492
[34] 王丽芳, 王德轩, 上官周平. 大穗型小麦叶片性状、养分含量及氮素分配特征[J]. 生态学报, 2013, 33(17): 5219–5227 WANG L F, WANG D X, SHANGGUAN Z P. Structural characters and nutrient contents of leaves as well as nitrogen distribution among different organs of big-head wheat[J]. Acta Ecologica Sinica, 2013, 33(17): 5219–5227
[35] 丁凡, 廉培勇, 曾德慧. 松嫩平原草甸三种植物叶片N、P化学计量特征及其与土壤N、P浓度的关系[J]. 生态学杂志, 2011, 30(1): 77–81 DING F, LIAN P Y, ZENG D H. Characteristics of plant leaf nitrogen and phosphorus stoichiometry in relation to soil nitrogen and phosphorus concentrations in Songnen Plain meadow[J]. Chinese Journal of Ecology, 2011, 30(1): 77–81
[36] 冯伟, 姚霞, 田永超, 等. 基于高光谱遥感的小麦叶片糖氮比监测[J]. 中国农业科学, 2008, 41(6): 1630–1639 FENG W, YAO X, TIAN Y C, et al. Monitoring the sugar to nitrogen ratio in wheat leaves with hyperspectral remote sensing[J]. Scientia Agricultura Sinica, 2008, 41(6): 1630–1639
[37] STERNER R W. Ecological Stoichiometry: Biology of Elements from Molecules to the Biosphere[M]. Princeton: Princeton University Press, 2002
[38] 朱新开, 严六零, 郭文善, 等. 淮北稻茬超高产小麦碳氮代谢特征研究[J]. 麦类作物学报, 2002, 22(1): 51–55 ZHU X K, YAN L L, GUO W S, et al. Carbon-nitrogen characteristics of super-high yielding wheat after rice in Huaibei Region[J]. Journal of Triticeae Crops, 2002, 22(1): 51–55
[39] SHEN J B, YUAN L X, ZHANG J L, et al. Phosphorus dynamics: From soil to plant[J]. Plant Physiology, 2011, 156(3): 997–1005
[40] MAKINO W, COTNER J B, STERNER R W, et al. Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C∶N∶P stoichiometry[J]. Functional Ecology, 2003, 17(1): 121–130
Effects of soil nitrogen and phosphorus contents on ecological stoichiometry of wheat leaf*
WANG Fankun, XUE Ke, FU Weiguo**
(Key Laboratory of Modern Agricultural Equipment and Technology of Jiangsu University and Ministry of Education / Key Laboratory of Agricultural Equipment and High-Tech Intelligentization of Jiangsu Province, Zhenjiang 212013, China)
Ecological stoichiometry is a science that studies the balance of energy and multiple chemical elements in the ecosystem. The contents of C, N and P and the related ecological stoichiometric ratios (N∶P, C∶N and C∶P) in plants can be used to judge the level of nutrient supply during plant growth period. In this paper, a technical guidance for precise fertilization in wheat production was provided by determining the response of ecological stoichiometry of major elements in wheat leaf to different soil nitrogen and phosphorus conditions. The weak gluten wheat variety ‘Yangmai 15’ and strong gluten wheat variety ‘Zhenmai 168’ that were widely cultivated in the study area were selected as test materials. Using pot experiment, 16 treatments of different nitrogen and phosphorus gradients were set up and the ecological stoichiometric ratios of N∶P, C∶N and C∶P at jointing, booting and grain-filling stages analyzed for different treatments. The results showed that: 1) at jointing and booting stages, when soil N∶P was 7.04–8.73, N∶P of the two wheat varieties leaves were higher. At grain-filling stage when soil N∶P was 8.73–10.42, N∶P of the two wheatvarieties was higher. There was significant positive correlation between N∶P in wheat leaf and N∶P in soil, but the correlation became weaker with wheat growth. 2) At a low soil N level (108.4 mg∙kg-1), leaf C∶N of the two wheat varieties was at a higher level. There was a significant negative correlation between C∶N in wheat leaf and N∶P in soil indicating that C∶N in wheat leaf decreased with increasing of N∶P in soil. 3) At low soil P level (29.6 mg∙kg-1), C∶P of the two wheat varieties leaves were higher as well. There was extremely significant negative correlation between C∶P in wheat leaf and P content of soil, which suggested that C∶P in wheat leaf decreased with increasing soil P content. The above results indicated that different supply levels of N and P in soil significantly changed leaf ecological stoichiometric ratios of different wheat varieties. Therefore, the steady-state characteristics of ecological stoichiometric ratio can be used as an important reference indicator for nutrient regulation in wheat production.
Ecological stoichiometry; Wheat leaf; Growth stage; Soil nitrogen and phosphorus
, E-mail: fuweiguo@ujs.edu.cn
Feb. 1, 2018;
Jun. 24, 2018
Q945.79
A
2096-6237(2019)01-0060-12
10.13930/j.cnki.cjea.180133
王凡坤, 薛珂, 付为国. 土壤氮磷状况对小麦叶片养分生态化学计量特征的影响[J]. 中国生态农业学报(中英文), 2019, 27(1): 60-71
WANG F K, XUE K, FU W G. Effects of soil nitrogen and phosphorus contents on ecological stoichiometry of wheat leaf[J]. Chinese Journal of Eco-Agriculture, 2019, 27(1): 60-71
* 江苏省农业科技自主创新资金[CX(15)1004]资助
付为国, 主要从事作物栽培研究。E-mail: fuweiguo@ujs.edu.cn
王凡坤, 主要研究方向为农业生态。E-mail: 13213185673@163.com
2018-02-01
2018-06-24
* This study was supported by the Agricultural Science and Technology Independent Innovation Funds of Jiangsu Province [CX(15)1004].
猜你喜欢
农业科技通讯(2022年11期)2022-11-30
现代农业科技(2022年20期)2022-11-09
核农学报(2022年11期)2022-10-29
作物学报(2022年9期)2022-07-18
成都信息工程大学学报(2022年2期)2022-06-14
华北农学报(2022年2期)2022-05-12
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年6期)2021-11-27
今日农业(2021年17期)2021-11-26
麦类作物学报(2021年1期)2021-04-12
