
夹尾刺激对大鼠背外侧前额叶和杏仁核神经网络振荡的影响
2018-12-07 03:48张忠楠王美美孙睿琦王邦安汪萌芽
皖南医学院学报 2018年6期
张忠楠,王美美,江 楠,孙睿琦,王邦安,汪萌芽
(皖南医学院 1.细胞电生理研究室;2.启明星小组,安徽 芜湖 241002)
神经振荡是大脑内有节律的放电现象,各脑区间的神经网络振荡在大脑执行高级功能时起重要作用[1],且神经振荡可以调节脑区间的信息交流强度[2]。背外侧前额叶(dorsolateral prefrontal cortex,DLPFC)和杏仁核(amygdala)[3]是情绪调节的重要核团,两核团的神经振荡在情绪调控中的作用还未见相关报道。因此,我们参照脑内双核团和外周生理指标同步记录技术[4],记录夹尾刺激前后DLPFC亚区岛叶无颗粒皮质背部(agranular insular area,AID)[5]和杏仁核亚核基底外侧杏仁核(basolateral amygdala,BLA)的细胞外电活动,以及心电、呼吸肌肌电,观察和比较夹尾刺激前后核团神经振荡的特征和外周生理指标的变化,为进一步研究伤害性刺激引起的负性情绪变化奠定基础。
1 材料和方法
1.1 实验动物 成年雄性SD大鼠12只,体质量260~350 g,由南京青龙山动物繁殖场提供。
1.2 仪器和药品 脑立体定位仪(瑞沃德68000系列)、保温毯系统(上海奥尔科特生物科技有限公司)、电生理屏蔽罩、防震台、PowerLab 八通道生理信号采集系统及其 Chart Pro 分析软件 (AD Instruments,Australia)、生物电放大器(美国 A-M Systems,MODEL 1800)、玻璃微电极和金属参考电极、20%乌拉坦、3%双氧水等。
1.3 实验方法
1.3.1 动物麻醉 用20%乌拉坦麻醉大鼠(1.5 g/kg,ip),直至夹捏刺激无明显反应。麻醉稳定后,将大鼠俯卧位固定于电生理屏蔽罩内的防震台上的脑立体定位仪上,下面垫保温毯,温度设置为37 ℃。
1.3.2 外周生理指标记录[4]大鼠右前肢腕关节、左后肢踝关节上部的皮下分别插入针式电极,记录标准肢体Ⅱ导联心电图,右后肢针式电极接地。将两根针式电极插入大鼠右侧腋中线第9肋下缘肋间外肌,记录呼吸肌肌电图。
1.3.3 脑内双核团细胞外记录[4]使大鼠咬合门齿杆并插入耳杆呈左右对称,固定大鼠头部使之不能移动,用手术刀切开头皮,3%双氧水反复擦拭直至暴露颅骨前囟、人字缝,调节前囟和人字缝至同一水平面。定位:BLA为前囟后1.56 ~3.36 mm,右旁开3.90 ~ 4.60 mm,硬脑膜表面以下7.80 ~ 8.82 mm;AID为前囟前2.52 ~ 4.20 mm,右旁开2.40 ~ 4.20 mm,硬脑膜表面以下4.40 ~ 5.40 mm[6]。用牙科钻钻颅开窗,以冷光源照明,用弯针头挑破硬脑膜、蛛网膜和软脑膜,玻璃电极(内充电极液)在操作臂的牵引下紧贴脑组织,安置好参考电极及接地电极,记录电极接入记录放大系统并下至目标脑区后寻找合适、明确的细胞外放电信号,待AID、BLA及各项外周生理指标信号稳定后,记录基础数据20 min并存盘。
1.3.4 伤害性刺激[7]使用充分绝缘的血管钳夹持大鼠尾巴中末1/3处,以后肢抽动为宜,5 s后撤离刺激,持续观察并记录刺激前20 min和刺激后40 min的各项信号。
1.3.5 信号采集参数设置[7-8]各项信号由Chart 5软件进行采集、存盘,采样频率4 kHz,心电为低通200 Hz、高通10 Hz,呼吸肌肌电为低通200 Hz、高通0.1 Hz,AID和BLA 为高通1 Hz、低通5000 Hz。
1.3.6 记录位点鉴定 实验结束,大鼠断头取脑于4%甲醛溶液内固定后,进行冠状面切片,观察滂胺天蓝标记位置,以鉴定记录位点。仅将记录位点正确的样本纳入结果的统计和分析。
1.4 记录信号分析
1.4.1 放电频率和神经振荡分析 对核团电信号高通80 Hz数字滤波[8]以分析放电频率。对核团放电原始数据带宽滤波得各神经振荡信号:delta(1~4 Hz),theta(4~10 Hz),beta(10~30 Hz),gamma(30~80 Hz),fast(80~200 Hz)[1],用FFT功能分析各神经振荡的能量。将各神经振荡信号另存为TXT格式文件,将两核团相同神经振荡的数据(TXT文件)导入MATLAB R2010b软件里,用Hilbert功能提取数据的相位信息,按照计算相位锁值(PLV)的公式进行编程[9]和分析。
1.4.2 外周生理指标分析[7]使用直接计数法对呼吸频率进行统计。使用记录一段时间内心率的平均值代表心率值。
1.5 统计分析 结果以均数±标准误表示,用SPSS 18.0软件进行数据的统计分析,刺激前后数据比较用配对t检验分析,P<0.05为差异有统计学意义。
2 结果
2.1 脑内双核团细胞外电活动和外周生理指标同步记录 对AID和BLA细胞外电活动、呼吸肌肌电(EMG)、心电(ECG)的同步记录(图1),经高通80 Hz滤波可得到单位放电记录(图2),而分波段进行带宽滤波则得到各神经振荡记录(图3)。

从上往下依次为右侧岛叶无颗粒皮质背部 (R-AID,μV)、基底外侧杏仁核 (R-BLA,μV)电信号,心电(ECG,μV),呼吸肌肌电(EMG,μV)。
图1 大鼠双核团细胞外电活动和外周生理指标同步记录

从上往下依次为右侧AID、BLA的单位放电(μV)。
图2 大鼠双核团单位放电记录

从上到下依次为delta、theta、beta、gamma、fast神经振荡(μV)。
图3 大鼠双核团神经振荡记录
2.2 夹尾刺激对大鼠AID、BLA放电活动的影响
2.2.1 放电频率变化 夹尾刺激对大鼠双核团放电和外周生理指标、双核团单位放电及双核图神经振荡的典型影响分例示于图4、5、6。分析12只大鼠AID、BLA刺激前2 min、刺激时5 s、刺激后2 min放电频率显示,夹尾刺激显著升高了AID和BLA的放电频率 [AID:差值(16.62±3.31)Hz,t=5.007,P<0.001;BLA:差值(14.80±3.27)Hz,t=4.528,P<0.01],刺激后均逐渐恢复(图7)。
2.2.2 神经振荡的能量变化 分析大鼠夹尾刺激前后各神经振荡的能量,结果刺激时两核团delta和theta振荡的能量减弱(P<0.05),gamma和fast振荡的能量有增强趋势(P>0.05),且都在刺激后恢复(图8)。
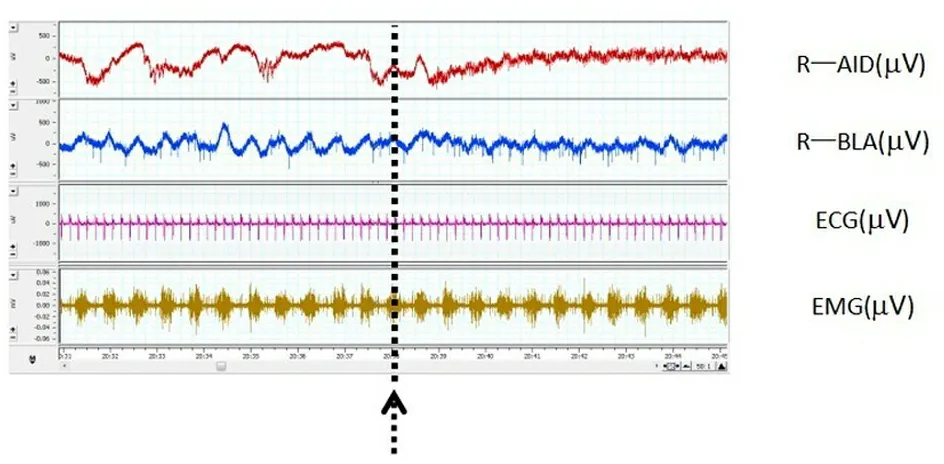
从上往下依次为右侧AID、BLA电信号(μV),心电(ECG,μV),呼吸肌肌电(EMG,μV);箭头表示夹尾刺激的开始,持续5 s。
图4 夹尾刺激对大鼠双核团细胞外电活动和外周生理指标的影响

从上往下依次为右侧AID、BLA的单位放电(μV);箭头表示夹尾刺激的开始,持续5 s。
图5 夹尾刺激对大鼠双核团单位放电的影响

从上到下依次为delta、theta、beta、gamma、fast神经振荡(μV);箭头表示夹尾刺激的开始,持续5 s。
图6 夹尾刺激对大鼠双核团神经振荡的影响

上图为AID(A)和BLA(B)的放电频率直方图,箭头表示刺激的开始,持续5 s;下图为AID(C)和BLA(D)的放电频率统计图 (n=12,配对t检验:*P<0.05,**P<0.01,***P<0.001)。
图7 夹尾刺激对AID和BLA放电频率的影响(bin=1s)

n=12,配对t检验:*P<0.05。
图8 夹尾刺激对两核团delta(A)、theta(B)、beta(C)、gamma(D)、fast(E)振荡能量的影响
2.2.3 神经网络振荡的相位锁值变化 计算分析大鼠夹尾刺激前后两核团间各神经振荡的相位锁值,结果刺激后两核团间delta、theta、fast振荡的相位锁值均降低(P<0.05),见图9。
2.3 夹尾刺激对外周生理指标的影响 分析大鼠夹尾刺激前后的心率、呼吸频率,显示刺激时心率升高(P<0.05),且刺激后并未立即恢复,呼吸频率升高(P<0.01),刺激后逐渐恢复(P<0.01),见图10。
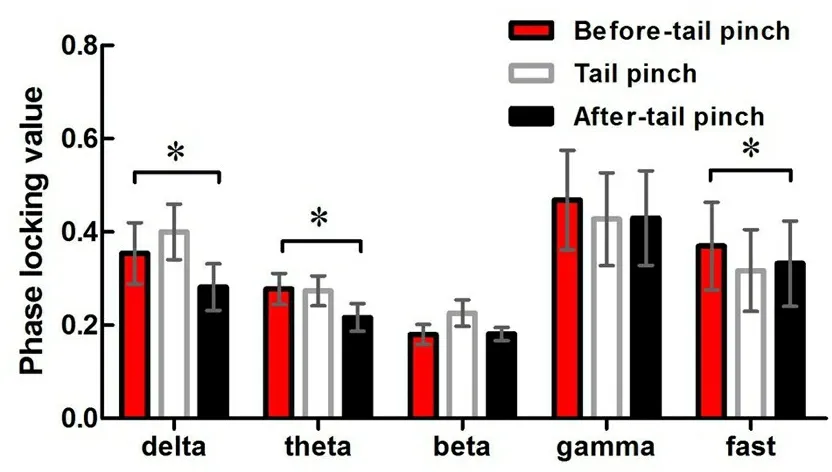
n=12,配对t检验:*P<0.05。
图9 夹尾刺激对AID和BLA间神经振荡相位锁值的影响

n=12,配对t检验:*P<0.05,**P<0.01。
图10 夹尾刺激对呼吸频率(A)和心率(B)的影响
3 讨论
杏仁核和背外侧前额叶都是负性情绪调控的关键脑区[3],而夹尾刺激可引起负性情绪的变化[7]。本实验同步记录和分析夹尾刺激前后正常大鼠AID、BLA的细胞外电活动、心电和呼吸肌肌电,结果刺激升高两核团的放电频率,并逐渐恢复,与之前夹尾刺激引起的变化相似[7];刺激降低了AID、BLA低频段(delta、theta)的能量,而高频段(gamma、fast)的能量有增高趋势,不同的神经振荡提示不同的生理状态,一般认为低频成分降低和高频成分增多反映脑电的去同步化状态[2],表明夹尾刺激对AID、BLA有激活作用。至于激活的具体通路,根据已有对BLA、内侧前额叶(mPFC)和DLPFC联系的研究[3,10],特别是有研究指出背外侧前额叶对杏仁核有自上而下的抑制作用[3]等,提示夹尾刺激影响AID、BLA放电有直接或间接通路的可能性,均有待进一步研究。另外,相位同步是相同节律的神经振荡在不同脑区间相互调制的表现,与脑区间的信息交流有密切联系[2,9],本实验中,刺激降低了AID和BLA间delta、theta振荡的相位锁值,说明夹尾刺激干扰了AID和BLA间低频段正常的信息交流。因此,夹尾刺激不仅改变了两核团delta和theta振荡的能量,而且还降低了核团间delta、theta振荡的相位锁值,同时结合反映情绪变化的外周生理指标[11],说明delta和theta振荡可能与伤害性刺激引起的相关负性情绪变化有联系,但是具体的过程和机制需进一步探索。
本实验运用脑内双核团和外周生理指标同步记录技术[4],记录了背外侧前额叶和杏仁核的细胞外电活动、心电和呼吸肌肌电,作为夹尾刺激引起情绪变化的指标,并分析两核团各神经振荡的特性,结果显示夹尾刺激引起了大鼠两核团神经振荡以及心率和呼吸频率的显著变化,将负性情绪调控的关键核团[3]的神经振荡与反应情绪变化的外周生理指标[11]结合分析,为进一步研究神经振荡在相关负性情绪调控中的作用奠定了基础。
猜你喜欢
江西农业大学学报(2022年6期)2023-01-31
磁共振成像(2022年6期)2022-07-30
中学生数理化(高中版.高考数学)(2022年1期)2022-04-26
中国医学影像技术(2022年2期)2022-03-01
房地产导刊(2022年1期)2022-02-28
海南师范大学学报(自然科学版)(2020年1期)2020-04-18
音乐生活(2020年12期)2020-01-05
中国老年学杂志(2017年13期)2017-07-18
计算机测量与控制(2017年6期)2017-07-01
中国实用神经疾病杂志(2012年14期)2012-12-17
