
黄鳝肠道假单胞杆菌铁蛋白基因的克隆及融合表达
2018-12-04 03:37:58臧彧伟王全禾
浙江农业学报 2018年11期
臧彧伟,王全禾,杨 龙,李 伟
(长江大学生命科学学院,湖北荆州434025)
铁离子是细菌生长的重要营养元素,但过量铁会引起细胞毒性[1]。保持铁离子的储存与代谢平衡对细菌至关重要[2]。细菌可能会通过铁蛋白或铁蛋白样分子储存胞质中过量的铁来达到平衡铁离子利用与减少细胞毒性的目的[3]。研究发现,细菌中共存着两种铁储存蛋白,一种为可结合血红素的Bfr型铁蛋白,另一种为不能结合血红素的Ftn型铁蛋白[4]。Ftn通常是一个由24个亚基组装的球形蛋白,其中央为一个可大量储存三价铁的空腔[5]。这种特殊的结构使得铁离子一直保持着可溶性状态并阻止其参与氧化还原反应从而造成活性氧胁迫[6]。Bfr型的铁蛋白也是一个24亚基的球形结构,不过其每两个亚基可以结合一个血红素分子,并且每个亚基中心都具有一个铁氧化酶中心[7]。球形结构中这些铁氧化酶中心高度对称[8]。研究发现,绿脓杆菌主要的储铁蛋白是Bfr,Ftn只能收集极少量的铁。尤其是在绿脓杆菌生长受到铁离子极限缺乏条件时,从Bfr中获得铁离子是维持生长的主要途径[2]。绿脓杆菌的Ftn型铁蛋白中铁离子释放需要铁氧还蛋白NADP还原酶(Fpr)提供电子;而Bfr铁蛋白不仅需要Fpr提供电子而且需要脱铁状态的Bfr相关铁氧还蛋白(apo-Bfd)协同作用才能实现铁离子的释放[2,9]。相反地,Ftn是大肠埃希菌的主要铁储存蛋白,其Bfr蛋白的功能尚不清楚[10]。尽管植物、动物和微生物的Ftn型和Bfr型菌铁蛋白亚基折叠方式和四级结构都很类似,它们的序列相似性却很低,这就极大地影响了其电荷交换的能力并造成了功能的巨大差异[3,11]。因此,开展不同生物铁蛋白基因分离和功能鉴定对于深入了解这些基因在铁离子代谢中的作用非常必要。黄鳝肠道假单胞杆菌是笔者实验室分离到的一种非致病微生物。该菌分类地位上与Pseudomonas sp.TKP菌株非常相似,具有产表面活性剂的能力[12]。对于该菌铁蛋白Bfr基因克隆及功能的研究国内外尚没有相关报道。因此,笔者拟通过克隆其Bfr基因,构建原核表达载体,进行重组蛋白的分离纯化及鉴定;分析重组蛋白铁离子结合能力并探讨其对该菌营养生长的影响,以期为深入了解假单胞杆菌Bfr基因的功能提供理论基础。
1 材料与方法
1.1 材料
1.1.1 质粒和菌株
原核表达载体pGEX-4T-1购买自Amersham Bioscience公司;大肠埃希菌感受态DH5α、BL21(DE3)、pEASY-T1载体等购自北京全式金公司;黄鳝肠道假单胞杆菌菌株由长江大学生物医药研究所鉴定并保存。
1.1.2 主要试剂
限制性内切酶、T4 DNA连接酶等购自Thermo Scientific公司;质粒DNA提取试剂盒、琼脂糖胶回收试剂盒、DNA聚合酶等购自北京全式金公司;异丙基硫代-β-D-半乳糖苷(IPTG)购自 Invitrogen公司;GST-resin纯化柱购自上海七海生物;DAB显色试剂盒购自北京中衫金桥生物科技公司;硝酸纤维素膜购自武汉谷歌生物,菲洛嗪购自上海生工。
1.2 方法
1.2.1 Bfr基因的克隆及序列分析
根据GenBank数据库已知假单胞杆菌属Bfr基因的序列设计一对基因特异性引物Bfr-F1(5’-ATGAAAGGCGACATCTCA-3’) 和 Bfr-R1(5’-TTATTCACCCATCTGCG-3’),以黄鳝肠道假单胞杆菌gDNA为模板,进行PCR扩增,扩增后产物用凝胶回收试剂盒进行纯化后克隆至pEASY-T1载体,转化大肠埃希菌DH5α感受态细胞后,选取阳性菌落进行序列测定。
测序所得序列在NCBI数据库进行核苷酸及氨基酸同源性分析。用DNAMAN软件进行多序列比对并分析其相似性和限制性酶切位点信息。
1.2.2 原核表达载体的构建
根据测序所得Bfr基因序列设计一对特异性表达引物rc-F(5’-TTAGAATTCATGAAAGGCGACATCTCAG-3’)和 rc-R(5’-GAACTCGAGTTATTCACCCATCTGCG-3’),以黄鳝肠道假单胞杆菌基因组为模板,进行PCR扩增。上、下游引物分别增加了 EcoRⅠ和 XhoⅠ限制性酶切位点。PCR产物经1%琼脂糖凝胶电泳后并用胶回收试剂盒回收纯化。将纯化产物及pGEX-4T-1载体用EcoRⅠ内切酶和XhoⅠ内切酶进行双酶切,电泳、回收纯化后加入T4 DNA连接酶于16℃进行连接反应。连接产物转化DH5α感受态细胞,经菌液PCR及双酶切验证后送往济南博尚生物技术有限公司测序。
1.2.3 重组蛋白的诱导表达、纯化及Westernblot检测
将测序正确的pGEX-4T-1-Bfr重组质粒转化BL21(DE3)感受态细菌,涂布在含有氨苄青霉素(50 μg·mL-1)的 LB 固体培养基上,37 ℃条件下倒置培养过夜。挑取单克隆接种于含氨苄青霉素(50 μg·mL-1)的 LB 液体培养基中,37 ℃,220 r·min-1振荡培养菌液 D600至 0.5 ~ 0.6,加入IPTG(125 μg·mL-1)继续于 37 ℃、220 r·min-1振荡培养3 h,取40 μL菌液,进行SDS-PAGE电泳检测蛋白表达情况。
将阳性菌株按照体积比1∶1 000接种于新鲜LB液体培养基中扩大培养,用IPTG诱导蛋白表达。于4 ℃、10 000 r·min-1离心 10 min 收集菌体,沉淀用Tris-HCl缓冲液(pH 8.0)重悬,冰上超声裂解细菌,超声5 s,间隔10 s,总时间15 min,4 ℃,10 000 r·min-1离心 10 min,取上清,用0.45 μm滤膜过滤后将上清加到GST-resin纯化柱中,用还原型谷胱甘肽(6 mmol·L-1)洗脱,获得GST-Bfr融合蛋白。
将纯化的融合蛋白重新加到GST-resin纯化柱中,用含有凝血酶(终浓度为3 U·mL-1)的Tris-HCl缓冲液酶切过夜,然后用Tris-HCl缓冲液洗脱Bfr蛋白,利用SDS-PAGE凝胶电泳检测Bfr蛋白酶切纯化情况。
将纯化得到的 GST-Bfr蛋白和Bfr蛋白经12%SDS-PAGE凝胶分离,转移到硝酸纤维素膜上,用5%脱脂牛奶4℃封闭过夜,再用Tris缓冲液-吐温(TBST)洗膜3次,每次10 min。随即加入小鼠抗GST-tag单克隆抗体(1∶5 000),37℃孵育2 h,用TBST洗膜6次,每次5 min。最后,加入辣根过氧化物酶(HRP)标记的山羊抗小鼠二抗(1∶10 000)于37℃孵育2 h,用DAB显色试剂盒显色。
1.2.4 重组蛋白铁离子螯合能力分析
在两支含 150 μL 浓度为 1 mmol·L-1的FeCl3(或FeCl2)试管中分别加入160 μL的GSTBfr融合蛋白(113 μg·mL-1)或 Bfr蛋白(113 μg·mL-1),补充Tris-HCl缓冲液至3 mL后于25℃、100 r·min-1振荡孵育 60 min,随即加入 120 μL的1%抗坏血酸,于室温孵育20 min,再加入120 μL 的菲洛嗪(5 mmol·L-1)室温孵育 10 min,最后用紫外分光光度计测定混合液在562 nm处的吸光值。以加入160 μL Tris-HCl缓冲液替代重组蛋白的作为空白对照,实验重复3次。
1.2.5 重组蛋白对假单胞杆菌生长的影响
挑取假单胞杆菌单克隆于液体LB培养基中30 ℃、220 r·min-1振荡培养 16 h。取 400 μL 培养液于40 mL新鲜 LB培养基中,加入2 mL FeCl3(1 mmol·L-1)和 100 μL Bfr蛋白(113 μg·mL-1),于30 ℃、220 r·min-1振荡培养12 h;摇动培养3 h后每间隔1 h取样一次,重复3次,用紫外分光光度计测定各时间点样品在600 nm处吸光值表征其生物量。以既不添加FeCl3也不添加Bfr蛋白的为阴性对照,以只添加FeCl3不添加Bfr蛋白的为阳性对照,实验重复3次。
2 结果与分析
2.1 黄鳝肠道假单胞杆菌Bfr基因的获得及序列分析
利用引物对Bfr-F1/R1从黄鳝肠道假单胞杆菌gDNA中扩增获得了一条长约450 bp的条带。将该PCR产物测序后发现该基因全长471 bp,编码一个长155 aa的多肽链(图1)。

图1 黄鳝肠道假单胞杆菌Bfr基因的核苷酸序列及推断的氨基酸序列Fig.1 The nucleotide sequence and deduced amino acid sequence of Bfr from Pseudomonas spp.
将测序所得的核苷酸序列和推断的氨基酸序列进行NCBI数据库Blast比对,发现该基因与其他不同微生物来源的铁蛋白基因具有较高的同源性。同时利用DNAMAN软件进行的多序列比对结果显示:该基因推断的氨基酸序列与来源于荧光假单胞杆菌的Bfr蛋白具有83.3%的同源性,而与耶尔森氏菌、双歧杆菌、荚膜红细菌、淋球菌、绿脓假单胞杆菌和沃尔巴克氏体来源的Ftn蛋白分别具有 49.4%、43.7%、40.2%、37.6%、45.9%和31.6%的同源性(图2),证明实验已成功克隆得到黄鳝肠道假单胞杆菌的Bfr基因。
2.2 原核表达载体的构建、表达纯化及鉴定
用基因特异性引物对rc-F/R从假单胞杆菌gDNA中扩增获得的片段,双酶切后与用同样限制性内切酶双酶切的pGEX-4T-1连接,经测序验证的pGEX-4T-1-Bfr重组质粒转化BL21(DE3)感受态细菌,挑取阳性菌株用IPTG进行诱导重组蛋白的表达。SDS-PAGE电泳结果显示,相对于诱导前和转空白质粒的菌株,诱导后产生了一条分子量约为50 ku的条带,符合预期大小(图3-A)。
通过GST-resin纯化柱结合带有GST标签的蛋白,用还原型谷胱甘肽洗脱GST-Bfr蛋白,将洗脱的蛋白重新上柱后,再用凝血酶进行酶切,经Tris-HCl缓冲液洗脱获得了Bfr蛋白,SDS-PAGE电泳检测显示纯化后基本无杂蛋白,酶切后得到的Bfr蛋白大小约24 ku,比用软件预测的略大,可能是凝血酶酶切后多余的氨基酸导致。
将SDS-PAGE凝胶中蛋白转移至硝酸纤维素膜上,分别用小鼠抗GST-tag单克隆抗体为一抗和辣根过氧化物酶(HRP)标记山羊抗小鼠二抗进行Western blot免疫印迹的结果显示,带有GST标签的融合蛋白被特异性识别,而酶切后的Bfr蛋白不能与抗体特异性识别,说明已成功实现了融合蛋白GST-Bfr和重组蛋白Bfr的纯化(图3-B)。
2.3 GST-Bfr蛋白和重组 Bfr蛋白与 Fe3+和Fe2+的螯合能力检测
利用铁螯合试剂菲洛嗪,笔者采用比色法检测了融合蛋白GST-Bfr蛋白和重组Bfr蛋白与Fe3+和Fe2+的螯合能力。结果显示,GST-Bfr蛋白在加入Fe3+和Fe2+后D562吸光值与对照均无明显差别;而Bfr蛋白在加入Fe3+后D562吸光值显著变小,加入Fe2+后D562吸光值与对照无显著差异。证明融合蛋白GST-Bfr既不能螯合Fe3+也不能结合Fe2+;而Bfr蛋白可以螯合Fe3+,但不能螯合Fe2+。
2.4 重组蛋白对假单胞杆菌生长的影响
在外源添加Fe3+情况下通过测定假单胞杆菌在不同培养时间后的生物量,分析重组蛋白有无对假单胞杆菌营养生长的影响。结果表明:相对于阴性对照和阳性对照,只有当Fe3+和Bfr蛋白同时存在时,才会促进假单胞杆菌的生长。这一定程度上说明Bfr蛋白对假单胞杆菌获得铁营养元素具有重要作用。

图2 Bfr推断的氨基酸与其他微生物铁蛋白序列比对结果Fig.2 Multiple alignment of ferritins from other species
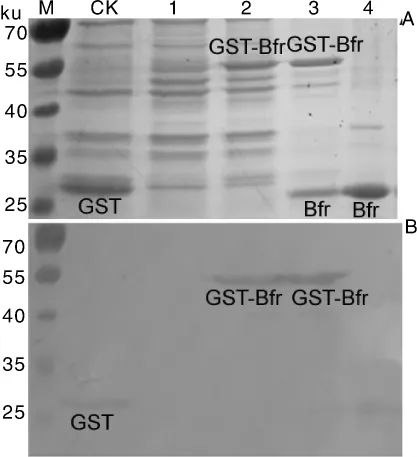
图3 融合蛋白的SDS-PAGE检测(A)与Western-blot分析(B)Fig.3 Expression(A)and detection of the recombinant proteins(B)
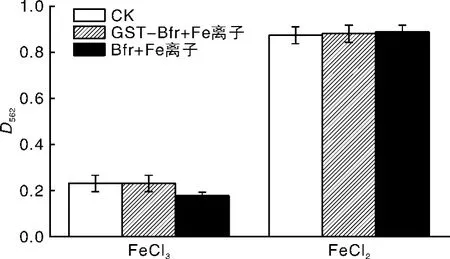
图4 Bfr蛋白与FeCl3、FeCl2的螯合能力分析Fig.4 Iron chelation detection of Bfr proteins
3 讨论
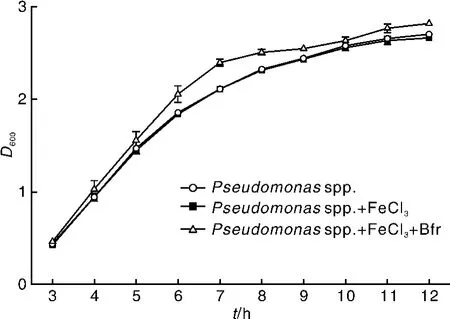
图5 不同培养时间假单胞杆菌的D600值Fig.5 D600values of Pseudomonas spp.at different time point
细菌铁蛋白Bfr是铁蛋白家族中的一员,同时也是一种重要的储存铁和解毒蛋白[13-14]。细菌铁储存蛋白是由24个亚基和12个血红素分子组成的球形蛋白,可以通过铁结合位点将Fe2+氧化[15],随后 Fe3+被移到中央内腔储存起来[16]。因此,铁蛋白可以在铁过量的条件下起一定的作用[6,17]。目前已对绿脓假单胞杆菌[2,7]和大肠埃希菌的[10]铁蛋白基因研究得比较透彻,而对其他假单胞杆菌的铁蛋白基因的功能不甚清楚。本研究成功克隆了黄鳝肠道假单胞杆菌的Bfr基因,序列分析发现该基因同目前已报道的Bfr类型的铁蛋白基因同源性较高,而同Ftn型铁蛋白同源性较低,是一个新的Bfr型铁蛋白家族的成员。在此基础上,笔者构建了原核表达载体,并实现了融合蛋白的表达;通过亲和层析获得了较纯的Bfr蛋白。铁离子螯合实验发现Bfr蛋白可以结合Fe3+离子但不能螯合Fe2+,这说明黄鳝肠道假单胞杆菌Bfr蛋白可能是一种重要的储铁蛋白。在外源添加Fe3+和Bfr蛋白时,能够一定程度上促进假单胞杆菌的生长,这可能是因为多余的Bfr蛋白结合Fe3+离子后起到了解毒效果并使得Bfr蛋白与细菌内的铁氧还蛋白相互作用形成复合物有利于保持细胞质铁稳态所致[18]。该研究为进一步分析不同微生物铁蛋白的功能提供了基础资料,应深入研究该类型细菌铁蛋白与黄鳝体内转铁蛋白受体相互作用获铁及转运机制。
猜你喜欢
食品工业科技(2023年4期)2023-02-14 10:12:58
医药前沿(2020年29期)2020-12-04 20:21:25
中国食品学报(2020年9期)2020-01-17 16:11:07
中国食品学报(2019年10期)2019-11-12 11:31:36
当代水产(2019年5期)2019-07-25 07:50:32
工友(2017年11期)2017-12-01 06:35:17
湖南农业(2016年3期)2016-06-05 09:37:36
中国洗涤用品工业(2015年2期)2015-02-28 19:02:00
黑龙江科学(2013年7期)2013-09-03 10:51:18
黑龙江八一农垦大学学报(2013年1期)2013-08-06 03:28:46
