
云南稻微核心种质籼粳回交后代功能成分含量差异及评价
2018-11-30 06:46杨加珍杨晓梦普晓英刘建伟曾亚文
西南农业学报 2018年10期
杜 娟,杨加珍,杨晓梦,杨 涛,李 霞,普晓英,刘建伟,曾亚文*
(1.云南省农业科学院生物技术与种质资源研究所,云南 昆明 650223;2.云南省农业生物技术重点实验室,云南 昆明 650223;3.昆明田康科技有限公司,云南 昆明 650205;4.云南农业大学农学与生物技术学院,云南 昆明 650201)
【研究意义】稻米是全球一半以上人口的主食。每天食用富含生物活性物质的功能稻米有助于降低糖尿病、高血压和其他心脑血管疾病的发病率,有益于身体健康。开发富含黄酮类化合物、抗性淀粉、γ-氨基丁酸、生物碱、维生素、钙铁锌硒、膳食纤维、N-去氢神经酰胺等成分的功能稻米既能促进现代营养医学的进一步发展,还可提高稻米的附加值[1]。【前人研究进展】优异育种材料匮乏、遗传基础狭窄是作物育种水平停滞不前的主要原因之一,构建微核心种质及拓宽遗传背景是解决问题的关键[2]。微核心种质即核中之核,利用分子标记数据进行 DNA 水平遗传冗余压缩核心种质样本,用约 10 %最具遗传变异的核心种质代表整个核心种质,能够有效降低成本提高育种效率;并可通过比较测序进行关联分析,实现基因高通量挖掘。有关大豆[3]、小麦[4]、花生[5]、玉米[6]等作物微核心种质构建、抗病性鉴定及其种质资源表型评价进展显著,水稻[7-9]微核心种质也有诸多研究,但未有功能稻米育种及评价的研究。王明明等[7]用来源于南方、华中和北方稻作区的174 份水稻微核心种质资源对白叶枯病的抗性进行鉴定,微核心种质在3个不同稻作区之间及籼粳亚种之间对白叶枯病的抗性反应存在明显基因分化。Zhang等[8]用36 对SSR标记将4 310份中国稻核心种质压缩为189份微核心种质,保留了70.65 %的SSR变异和76.97 %的表型变异。金铭路等[9]以来源于26个省、市的204份中国水稻微核心种质为试验材料,进行耐冷性鉴定及其相关性分析,结果表明发芽期、芽期、幼苗期、孕穗期的耐冷性粳稻均明显强于籼稻。【本研究切入点】培育功能稻品种有效提高糙米中功能成分含量[10],是当前功能水稻选育的首要问题。云南是世界公认的亚洲栽培稻籼粳亚种分化中心[11],也是中国稻种最大的生态多样性中心和优异种质功能稻米资源的富集中心,经过长期的人工选择和栽培驯化形成云南稻种资源有利基因[12]。本项目已利用548份云南稻种核心种质及其改良后代进行糙米功能成分评价和鉴定[13-15],选育出米饭抗性淀粉含量超过10%的功米3、 4号及抗性淀粉、γ-氨基丁酸双高的云资粳82号(抗性淀粉14.1 %,发芽糙米GABA =59.30 mg/100g)。以108份云南稻微核心种质及其改良后代研究4种功能成分(总黄酮、γ-氨基丁酸、生物碱和抗性淀粉)差异及鉴定评价至今尚未有研究报道。【拟解决的关键问题】以云南稻微核心种质及其回交后代为研究材料,对4种功能成分差异进行评价分析,旨在为利用微核心种质有效降低成本、加快功能稻育种及推进产业化提供参考。
1 材料与方法
1.1 试验材料种植方法
以108份富含糙米总黄酮、γ-氨基丁酸、生物碱和抗性淀粉功能成分的云南稻微核心种质作为供体,其中籼稻48份,粳稻60份。以籼稻滇屯502和粳稻合系35为轮回亲本,按籼籼交、粳粳交配制,获得一一对应的回交后代127份,其中BC5F421份、BC4F412份、BC3F416份、BC6F335份、BC5F319份、BC4F324份。所有材料于2015年3月22日在籼粳交错区的玉溪市研和镇播种,5月18日移栽,水肥按照常规管理方案,9月25日成熟期收获株系,随机混收,用日本佐竹SATAKE砻谷机(THU35C)脱壳成糙米,高速万能粉碎机(DFT-50A)打成粉60 ℃恒温干燥后待测。
1.2 功能性成分测定
在云南省农业科学院云南省农业生物技术重点实验室,测定4种功能性成分含量。γ-氨基丁酸的测定方法参照INATOMI[16]方法进行,回归方程:Y=5.152x+0.003, 相关系数为0.9984;总黄酮参照申湘忠等[17]的方法,回归方程:Y=2.702x-0.0129,相关系数为0.9994;生物碱参照曾亚文[18]方法,回归方程:Y=6.204x-0.186,相关系数为0.9962;抗性淀粉参照Goni[19]方法。
2 结果与分析
2.1 微核心种质及其回交后代糙米功能成分含量变异
由表1可知,总体来看,微核心种质和其回交后代糙米4种功能成分含量的变幅和变异系数均较大,回交后代糙米的4种功能成分含量较轮回亲本(滇屯502、合系35)均有明显的提高。微核心种质供体变异系数最大的是粳稻回交后代抗性淀粉(70.2 %),粳稻供体黄酮含量次之(65.7 %);变异系数最小的是粳稻回交后代γ-氨基丁酸含量(8.8 %)。供体微核心种质糙米功能成分变异系数从大到小依次是:黄酮(69.2 %)>抗性淀粉(59.4 %)>γ-氨基丁酸(45.8 %)>生物碱(16.2 %);回交后代变异系数从大到小依次是:抗性淀粉(60.5 %)>生物碱(37.8 %)>黄酮(27.0 %)>γ-氨基丁酸(19.2 %)。粳稻供体黄酮、γ-氨基丁酸、抗性淀粉含量高于籼稻,而生物碱含量则为籼稻高于粳稻。表明微核心种质水稻4种功能成分含量存在明显的基因型差异,粳稻供体更有利于选育出富含黄酮、γ-氨基丁酸、抗性淀粉的育种材料,而籼稻供体更易于选择出富含生物碱的育种材料。
表1云南稻微核心种质供体、受体及回交后代功能成分含量的变幅和变异系数
Table 1 The ranges and variation coefficients of functional ingredients in different backcross populations of mini-core collections rice in Yunnan province

材料Materials类型Types黄酮Total flavonoidγ-氨基丁酸Gamma-aminobutyric acid生物碱(mg/100g)Alkaloid抗性淀粉Resistant starch变幅 (mg/100g)Ranges变异系数(%)Variation coefficient变幅(mg/100g)Ranges 变异系数(%)Variation coefficient变幅(mg/100g)Ranges变异系数(%)Variation coefficient变幅(%)Ranges变异系数(%)Variation coefficient供体Donor籼稻34.3~273.260.52.3~10.040.58.9~15.611.00.74~6.4260.3粳稻47.9~430.665.70.75~15.146.38.5~14.511.40.8~5.052.0总和34.3~430.669.20.75~15.145.88.5~15.616.20.74~6.4259.4轮回亲本Recurrent parentDT50250.63-6.82-12.62-0.63-HX3551.84-4.86-13.95-1.02-回交后代Backcross population籼稻31.5~130.735.84.3~12.218.39.5~38.040.51.2~3.626.5粳稻49.2~96.012.06.6~10.08.810.0~19.913.11.2~13.470.2总和31.5~130.727.04.3~12.219.29.5~38.037.81.2~13.460.5
2.2 微核心种质籼粳各回交后代糙米功能成分的含量差异
由表2可知,微核心种质及籼粳各回交后代糙米γ-氨基丁酸、总黄酮含量均为粳稻>籼稻,BC6F3、BC5F3籼粳稻差异达到极显著及显著水平;而生物碱含量为籼稻>粳稻,BC5F4、BC6F3之间差异达到显著和极显著水平;抗性淀粉籼粳差异不大,未达到显著水平。表明在水稻高功能成分材料选育过程中,粳稻回交后代具有选育富含黄酮、γ-氨基丁酸等成分材料的优势,而籼稻回交后代具有选育高生物碱含量材料的优势。
2.3 微核心种质籼稻各回交后代之间4种功能成分的含量差异
由表3可知,微核心种质籼稻各回交后代之间黄酮、γ-氨基丁酸含量差异不显著;生物碱和抗性淀粉含量在各回交后代之间差异较大,BC5F4含量最高且和其他回交后代间差异达极显著和显著水平。表明回交次数和自交代数对籼稻回交后代生物碱及抗性淀粉含量有显著影响,对总黄酮、γ-氨基丁酸含量无较大影响。进一步证明籼稻回交后代群体有利于筛选出富含生物碱的功能稻米材料。
在籼稻各回交后代中,筛选出供体亲本及其对应回交后代糙米总黄酮、γ-氨基丁酸、生物碱和抗性淀粉含量分别都较高的5对优良品系,其糙米总黄酮变幅为34.5~80.8 mg/100g;γ-氨基丁酸变幅为4.7~7.0 mg/100g;生物碱变幅为12.1~30.5 mg/100g;抗性淀粉变幅为2.5 %~6.2 %。这些株系可以作为功能稻米选育的桥梁材料及糙米功能成分基因定位的近等基因系材料。
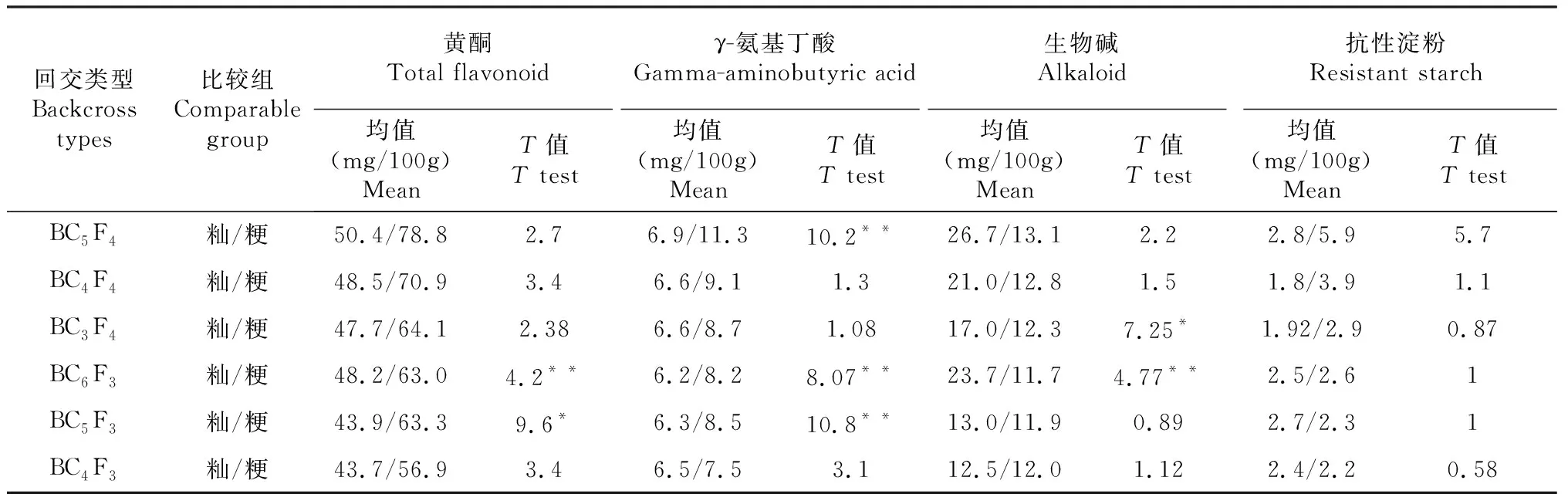
表2 微核心种质籼粳各回交后代之间糙米功能成分的含量差异
注:*和**分别表示0.05和0.01显著水平。
Note: * and ** indicate significant levels of 0.05 and 0.01, respectively.

表3 微核心种质籼稻各回交后代糙米功能成分的含量差异

表4 微核心种质粳稻各回交后代糙米功能成分的差异
2.4 微核心种质粳稻各回交后代之间4种功能成分的含量差异
由表4可知,微核心种质粳稻各回交后代之间生物碱含量差异不显著;总黄酮、γ-氨基丁酸及抗性淀粉在各回交后代含量差异较大,BC5F4含量最高和其他回交后代达极显著和显著水平。回交后代相同回交次数不同自交次数比较,总黄酮、γ-氨基丁酸、抗性淀粉随自交次数增加含量不断增加。BC5F4粳型回交后代中总黄酮、γ-氨基丁酸、抗性淀粉均值最高,说明粳型回交后代群体有利于筛选出富含黄酮、γ-氨基丁酸、抗性淀粉的功能稻米材料。
在粳稻各回交后代中,也筛选出供体亲本及其对应回交后代糙米总黄酮、γ-氨基丁酸、生物碱和抗性淀粉含量相对都较高的5对优良品系,其糙米总黄酮变幅为71.8~95.4 mg/100g;γ-氨基丁变幅为8.9~13.8 mg/100g;生物碱变幅为12.3~15.1 mg/100g;抗性淀粉变幅为3.2 %~5.1 %。
3 讨 论
水稻功能成分含量是与人类饮食健康密切相关的重要指标。因此在功能稻米选育中,对育种亲本材料的挑选应十分注重。孙丹等[14]对功米3 号和滇屯502 及其219 株F3家系糙米总黄酮和γ-氨基丁酸测定,F3群体糙米总黄酮平均值为130.15 mg/100g,变异系数18.83 %;F3群体糙米γ-氨基丁酸平均值为6.91 mg/100g,变异系数44.28 %。魏明亮[20]研究云南稻核心种质粳稻回交高代糙米的抗性淀粉含量变异系数高达59.2 %。本试验轮回亲本(合系35、滇屯502)糙米的功能成分含量较低,以108份云南稻微核心种质作为供体,整体来看,微核心种质和其回交后代糙米黄酮、γ-氨基丁酸、抗性淀粉、生物碱含量的变幅和变异系数均较大,四种功能成分4量存在较为明显的基因型差异,且回交后代中,4种功能成分较受体滇屯502和合系35均得到提高。说明以微核心种质及滇屯502及合系35作为受体培育具有高功能成分近等基因系的方案是可行的,可直接作为新的优良种质资源继续下一步的工作。粳型回交后代具有选育富含总黄酮、γ-氨基丁酸等成分材料的优势;籼型回交后代群体具有选育出富含生物碱的功能稻米的优势。这在高功能成分水稻育种工作方面也具有一定的参考价值。
另外,回交后代总黄酮含量高于轮回亲本,而低于微核心种质亲本。曾亚文[15]利用分光光度法检测云南栽培稻糙米发现紫米和红米的总黄酮、γ-氨基丁酸含量均极显著高于白米,日常饮食以白精米为主可能导致人类慢性病增加。杜娟[21]利用高效液相色谱法检测结果表明云南有色稻总黄酮极显著高于云南无色稻。控制总黄酮含量性状的基因,受细胞质遗传影响较大,与米色基因连锁[22]。有色稻米的种皮颜色是由类黄酮色素化合物沉积形成,回交后代材料经过多次回交配置,在育种过程中回交后代有色稻减少,白色稻增加可能是导致回交后代总黄酮降低的主要原因。因此育种中田间筛选环节应注意保留有色稻株系。
回交后代的生物碱含量的变异系数比微核心种质更大,且γ-氨基丁酸含量高于轮回亲本,也高于微核心种质,呈现超亲优势,与陈志远[23]的研究结果一致。稻米中γ-氨基丁酸含量受外界环境影响较大,干旱、低温、虫害、盐胁迫、机械刺激等逆境条件下,植物体内 γ-氨基丁酸大量富集而增加[24-25],因此在选育高 γ-氨基丁酸含量的功能稻米时,要关注外界环境对γ-氨基丁酸含量的影响。
4 结 论
本实验表明云南微核心种质供体及籼粳回交后代糙米的总黄酮、γ-氨基丁酸、抗性淀粉含量均存在丰富的变异,说明以云南核心种质及滇屯502、合系35培育具有高功能成分稻米的方案是可行的。
猜你喜欢
今日农业(2021年15期)2021-10-14
中国食品(2021年17期)2021-09-27
农民致富之友(2020年32期)2020-12-03
中国动物传染病学报(2020年2期)2020-04-02
中国粮油学报(2019年4期)2019-07-12
中国稻米(2018年5期)2018-01-21
中国酿造(2017年12期)2017-12-28
农村百事通(2017年8期)2017-04-26
亲子(2014年4期)2014-05-10
食品工业科技(2014年13期)2014-03-11
