
云南鸡足山山桂花和野桂花群落的格局与环境解释
2018-11-23 02:36:20李韦鸿杨国栋李涌福王贤荣段一凡朱轶人
植物资源与环境学报 2018年4期
李韦鸿, 杨国栋, 李涌福, 王贤荣, 从 睿, 段一凡,①, 朱轶人
(南京林业大学: a. 南方现代林业协同创新中心, b. 生物与环境学院, c. 林学院, 江苏 南京 210037)
山桂花(OsmanthusdelavayiFranch.)和野桂花〔O.yunnanensis(Franch.) P. S. Green〕隶属于木犀科(Oleaceae)木犀属(OsmanthusLour.),均为中国特有香花植物,也是木犀属中少有的春花植物,四季常绿,花朵芳香洁白,食用和观赏价值较高,是优良的园林绿化乔木或灌木。山桂花仅分布在云南、四川和贵州海拔2 100~3 400 m的区域[1]109-111,已有研究集中在品种分类[2]、系统学[3]、花粉形态[4]及显微鉴别[5]方面,近期相关研究较少。野桂花资源稀缺,仅分布在云南、四川和西藏海拔1 350~2 800 m的区域[1]101-103,目前对其研究仅限于群落特征[6]和化学成分[7]等方面,有关野桂花分布的报道也较少。山桂花和野桂花分布范围狭窄但开发前景广阔,云南鸡足山山顶树种过于单一造成病虫害加剧,加之人类活动和环境变化造成山桂花和野桂花群落和分布受到极大的破坏,亟待采取有效措施对其加以保护,因此,开展群落研究十分必要。
生物群落常被用来衡量生态系统质量[8],群落结构取决于多种环境因子的综合作用[9]。分类和排序是植被生态学研究中用于研究群落与环境之间关系的基本手段[10],用以把对象分组或按照生态梯度排序,解释其生态相关性[11]。植物生态学中常用的分类和排序方法有双向指示种分析(two way indicator species analysis,TWINSPAN)、除趋势对应分析(detrended canonical correspondence analysis,DCCA)以及典范对应分析(canonical correspondence analysis,CCA)[12-13]等,但上述这些方法受二维限制对信息模糊或者多因子作用的情况会造成结果不精确或不适用。自组织特征映射网络(self-organizing feature map,SOM)可以实现对非线性、复杂数据的可视化,输入样本后可自动将高维数据点降维,将输入层和输出层神经元之间的连接强度组成权重矩阵,通过权值调整使最后得到的二维映射层可以比较合理地反映样本的分布特征[14]。SOM在足够多次迭代的情况下能实现很好的聚类效果[15-16],并已在生态学和航空航天等领域得到很好的应用[17-18]。苏日古嘎等[19]利用SOM对北京松山自然保护区山地草甸群落进行分类,发现群落类型主要受海拔等环境因子的影响。目前,SOM已广泛应用于群落分类和动态预测等方面[20-23]。针对云南鸡足山山桂花和野桂花分布区域环境因子多样的状况,SOM相较传统方法有更好的适用性。
云南鸡足山海拔2 300 m以上的区域集中分布较多的山桂花和野桂花,对研究二者群落特征有一定的代表性。笔者在调查中发现山桂花和野桂花在不同的海拔和土壤等环境条件下的分布区域和群落格局存在很大差异,2种植物群落的格局以及环境因子对其群落分布的影响尚不清楚。本研究采用SOM对云南鸡足山的山桂花和野桂花群落进行研究,以期揭示2种植物不同的群落特点及其与环境因子之间的关系,从而更好地了解其生存现状,为山桂花和野桂花的后续研究和资源保护以及引种培育提供理论参考依据。
1 研究地概况和研究方法
1.1 研究地概况
云南鸡足山位于云贵高原滇西北宾川县境内(北纬25°56′~26°00′、东经100°20′~100°25′),地处金沙江干热河谷流域,受印度洋暖湿西南季风和大陆气团影响,夏季暖湿多雨,冬季低温干燥,属典型的亚热带半湿润气候特征。山体海拔3 240 m,气候立体多样,垂直高度变化大,被3条较大的山谷割裂,形似鸡足,地形生境复杂多样,满足不同海拔、坡向和土壤的要求,对调查植物群落类型与环境的关系具有一定的代表性。山桂花和野桂花分布区域多为亚热带山地常绿-落叶阔叶混合林,鸡足山则以元江锥(CastanopsisorthacanthaFranch.)、高山锥(C.delavayiFranch.)和滇青冈(CyclobalanopsisglaucoidesSchott.)为上层优势种形成半湿润常绿阔叶林[24]。山体中下部年均温11.9 ℃,最热月气温17.3 ℃,最冷月气温4.7 ℃,年降水量1 192.3 mm,其中雨季降水量占89%[25]。
1.2 研究方法
1.2.1 样地设置和指标观测 采用经典样地法按照海拔从低到高(2 300~3 200 m)分别设置面积20 m×20 m的山桂花样地5个以及野桂花样地7个,总面积4 800 m2,每个样地再设置面积10 m×10 m的样方4个,并从左下开始按顺时针方向编号,按海拔从低到高编号1~48,另外,每个样方中设置1个面积5 m×5 m草本层小样方。样地设置遵循种分布均匀、结构完整、层次分明、环境条件一致、群落中心部位避免过渡的原则。
对上述48个样方内胸径大于1 cm的乔木进行每木检尺,分别测量其胸径、树高和冠幅,并统计各树种株数;灌木和胸径小于1 cm的小乔木统计株数、盖度和生长情况;草本统计种类、盖度和生长情况。对12个样地的海拔、坡度、坡位、坡向以及土壤的厚度和含水量进行统计和测量。采用经纬仪测量海拔。采用坡度仪测量坡度。按照Zhang等[26]的方法划分坡位和坡向,上坡位记为1,中坡位记为2,下坡位记为3;采用罗盘仪测量坡向,规定正北为0°,按照45°进行划分,北坡记为1,东北坡记为2,西北坡记为3,东坡记为4,西坡记为5,东南坡记为6,西南坡记为7,南坡记为8,其中,北坡为阴坡,东北坡、西北坡和东坡为半阴坡,西坡、东南坡和西南坡为半阳坡,南坡为阳坡。参考文献[27-28],分别在样地的四角以及中间取样测量土壤的厚度和含水量,采用挖剖面的方式测量土壤厚度;采用环刀法取土样,烘干后称取质量,根据公式“土壤含水量=〔(烘干前铝盒和土样的质量-烘干后铝盒和土样的质量)/(烘干后铝盒和土样的质量-烘干后铝盒的质量)〕×100%”计算土壤含水量。
1.2.2 群落物种组成 参照恩格勒系统和哈钦松系统统计各样地科、属和种的数量,分析群落物种组成,研究亲缘关系,然后确定群落类型和结构[29]。
1.3 数据处理
1.3.1 重要值分析 参考管杰然等[30]的方法分别计算乔木、灌木和草本种类的重要值。乔木种类的重要值=(相对密度+相对优势度+相对频度)/3,灌木种类的重要值=(相对密度+相对盖度+相对频度)/3,草本种类的重要值=(相对高度+相对盖度+相对频度)/3。其中,相对密度=(某植物种类个体总数/所有植物种类个体总数)×100%,相对优势度=(某植物种类胸径总和/所有植物种类胸径总和)×100%;相对频度=(某植物种类频度总和/所有植物种类频度总和)×100%;相对盖度=(某植物种类盖度总和/所有植物种类盖度总和)×100%;相对高度=(某植物种类高度总和/所有植物种类高度总和)×100%。
1.3.2 SOM分类排序 使用SOM对48个样方进行分类排序,输入样本后通过自主学习调整自动确定最佳匹配单元,将高维数据点降维、保序地映射到二维网格上[31],得到样方分类排序的神经网络蜂巢图。使用K-means算法进行运算,通过计算戴维森堡丁指数(Davies-Bouldin index,DBI)评估K-means算法中K值(聚类数)的取值,DBI值越小,说明在该K值下聚类效果越好[32]。SOM详细计算过程参考文献[33-34]。SOM分类和排序在Matlab 7.14平台上用SOM工具箱(SOM toolbox)完成。
1.3.3 多元回归树检验 参考赖江山等[35]的方法选择生境作为连续分布的森林群落的参考节点。采用多元回归树(multivariate regression trees,MRT)法对山桂花和野桂花群落的48个样方进行SOM分类验证,将环境因子梯度作为分类节点,每次将样方二歧式分割,直至满足某一条件或不能再分割,然后用物种数据定义的样方划分类别,并采用交叉验证确定分类结果,使误差和MRT的规模尽可能小。MRT采用R语言mvpart_1.6-1程序包中的mvpart函数进行运算。
2 结果和分析
2.1 山桂花和野桂花群落的种类组成
在调查的云南鸡足山山桂花和野桂花群落中分布植物167种,隶属于68科114属,其中,蕨类植物5科8属10种,裸子植物2科2属2种,被子植物61科104属155种。壳斗科(Fagaceae)、杜鹃花科(Ericaceae)、蔷薇科(Rosaceae)、菊科(Asteraceae)、忍冬科(Caprifoliaceae)、卫矛科(Celastraceae)和山茶科(Theaceae)等植物在群落中较为常见,在科组成中所占比例较大。在山桂花和野桂花群落中单种科所占比例较大,为39.7%,如川续断科(Dipsacaceae)、杜英科(Elaeocarpaceae)、马桑科(Coriariaceae)和安息香科(Styracaceae)等。
2.2 山桂花和野桂花群落中重要值排名前10的乔木和灌木种类
在调查的云南鸡足山山桂花和野桂花群落的12个样地中,山桂花和野桂花的数量优势明显。5个山桂花样地中,山桂花在3个样地的相对密度大于30%,在2个样地的相对密度甚至达到60%,为绝对的优势种;7个野桂花样地中,野桂花在5个样地的相对密度大于40%,远高于其余植物种类。山桂花和野桂花在各样地的相对频度为15%~17%,分布较为均匀,且高于其余植物种类。5个山桂花样地中,山桂花在其中2个样地的相对优势度分别仅5.1%和1.9%,在其余3个样地的相对优势度较高,均在20%以上;7个野桂花样地中,野桂花在其中2个样地的相对优势度较高,分别为17.6%和8.1%,在其余5个样地的相对优势度较低,均在2%左右。
云南鸡足山山桂花群落中重要值排名前10的乔木和灌木种类见表1。由表1可见:山桂花群落中山桂花的重要值最高。重要值大于2.0%的9种乔木中,除短翅卫矛(EuonymusrehderianusLoes.)和绢毛稠李(PaduswilsoniiSchneid.)外,其余全部为常绿乔木。重要值大于2.0%的灌木种类中,山桂花和滇青冈为常绿灌木,其余3种植物为落叶灌木种类。说明除松科(Pinaceae)植物外,山桂花群落中壳斗科、木犀科(Oleaceae)和山茶科等科的常绿乔木和灌木种类较多,常绿乔木种类在建群乔木种类中占比较大。
表1云南鸡足山山桂花群落中重要值排名前10的乔木和灌木种类
Table1ToptenarborandshrubspeciesaccordingimportantvalueinOsmanthusdelavayiFranch.communityinJizuMountainofYunnanProvince
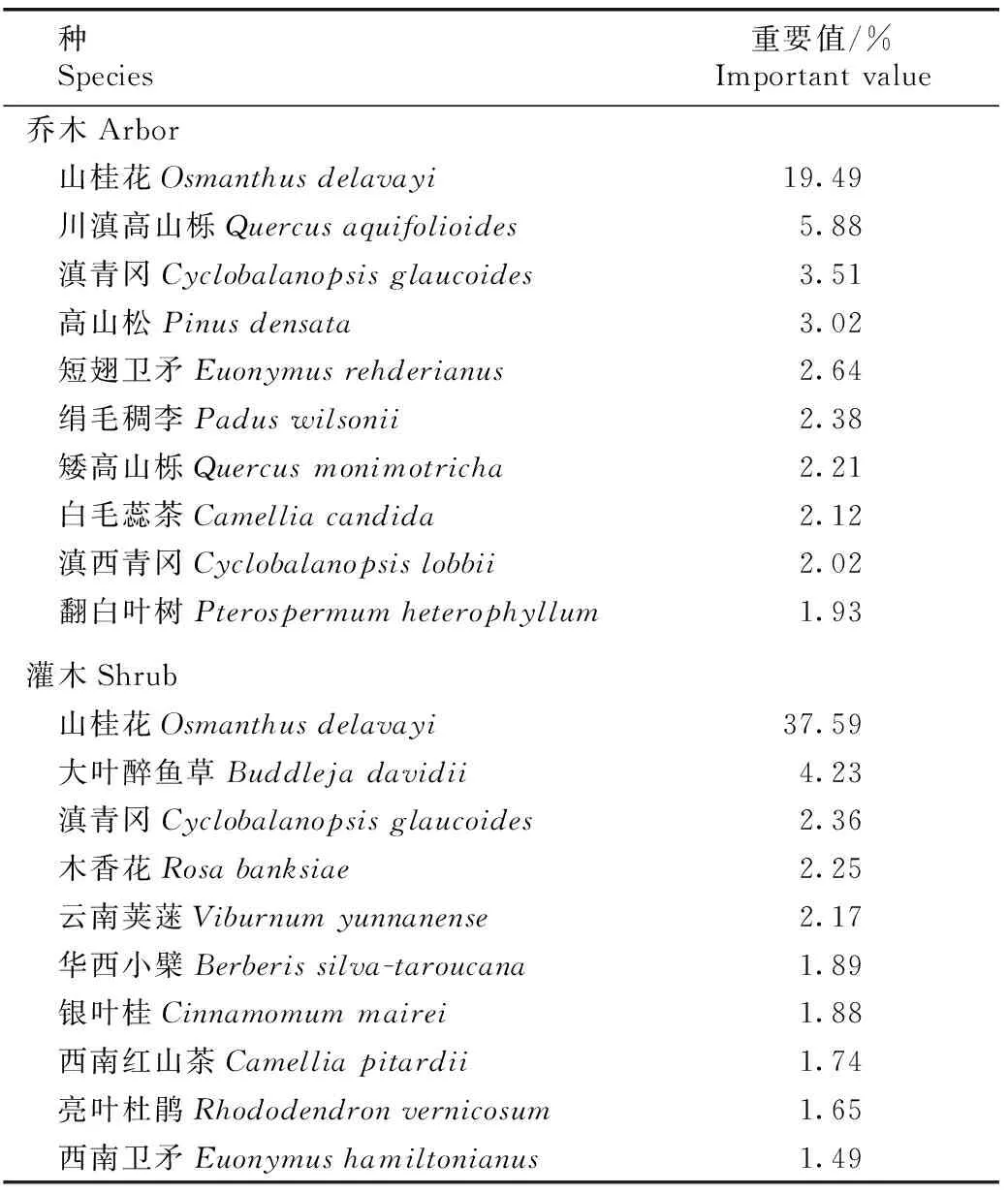
种 Species重要值/%Important value乔木Arbor 山桂花Osmanthus delavayi19.49 川滇高山栎Quercus aquifolioides5.88 滇青冈Cyclobalanopsis glaucoides3.51 高山松Pinus densata3.02 短翅卫矛Euonymus rehderianus2.64 绢毛稠李Padus wilsonii2.38 矮高山栎Quercus monimotricha2.21 白毛蕊茶Camellia candida2.12 滇西青冈Cyclobalanopsis lobbii2.02 翻白叶树Pterospermum heterophyllum1.93灌木Shrub 山桂花Osmanthus delavayi37.59 大叶醉鱼草Buddleja davidii4.23 滇青冈Cyclobalanopsis glaucoides2.36 木香花Rosa banksiae2.25 云南荚蒾Viburnum yunnanense2.17 华西小檗Berberis silva-taroucana1.89 银叶桂Cinnamomum mairei1.88 西南红山茶Camellia pitardii1.74 亮叶杜鹃Rhododendron vernicosum1.65 西南卫矛Euonymus hamiltonianus1.49
云南鸡足山野桂花群落中重要值排名前10的乔木和灌木种类见表2。由表2可见:野桂花群落中野桂花的重要值最高。重要值排名前10的乔木中,野桂花、高山锥、滇青冈、茶叶卫矛(EuonymustheifoliusWall.)和高山松(PinusdensataMast.)的重要值大于2.0%,且除茶叶卫矛外,其余均为常绿乔木。重要值排名前10的灌木有野桂花、毛果珍珠花〔Lyoniaovalifoliavar.hebecarpa(Franch. ex Forb. et Hemsl.) Chun〕和茶叶卫矛等,这10种灌木的重要值均大于2.0%,且除野桂花和滇青冈外,其余均为落叶灌木种类。说明野桂花群落中壳斗科、木犀科和松科等科的常绿乔木和灌木种类较多,常绿乔木种类在建群乔木种类中的占比较大。
表2云南鸡足山野桂花群落中重要值排名前10的乔木和灌木种类
Table2ToptenarborandshrubspeciesaccordingimportantvalueinOsmanthusyunnanensis(Franch.)P.S.GreencommunityinJizuMountainofYunnanProvince

种 Species重要值/%Important value乔木Arbor 野桂花Osmanthus yunnanensis22.05 高山锥Castanopsis delavayi4.58 滇青冈Cyclobalanopsis glaucoides2.43 茶叶卫矛Euonymus theifolius2.13 高山松Pinus densata2.11 绒毛杜鹃Rhododendron pachytrichum1.96 亮叶厚皮香Ternstroemia nitida1.68 银叶桂Cinnamomum mairei1.39 灯台树Bothrocaryum controversum1.34 宽叶杜鹃Rhododendron sphaeroblastum1.33灌木Shrub 野桂花Osmanthus yunnanensis10.30 毛果珍珠花Lyonia ovalifolia var. hebe-carpa5.21 茶叶卫矛Euonymus theifolius5.13 绢毛稠李Padus wilsonii3.14 滇青冈Cyclobalanopsis glaucoides3.08 四蕊槭Acer tetramerum2.74 乌桕Sapium sebiferum2.68 铁仔Myrsine africana2.59 云南荚蒾Viburnum yunnanense2.40 青榨槭Acer davidii2.22
云南鸡足山山桂花和野桂花群落中菊科、蔷薇科、景天科(Crassulaceae)、报春花科(Primulaceae)和虎耳草科(Saxifragaceae)的草本种类均较多。
2.3 山桂花和野桂花群落分类
2.3.1 自组织特征映射网络(SOM)分类结果 经SOM处理后确定量化误差和拓扑误差最小的最终输出层为8×7的拓扑结构,得到山桂花和野桂花群落48个样方的拓扑映射图。根据K-means聚类规则,选择戴维森堡丁指数(DBI)最小处的聚类数6,据此将拓扑映射图划分为6块区域,对应6个群落类型(图1)。
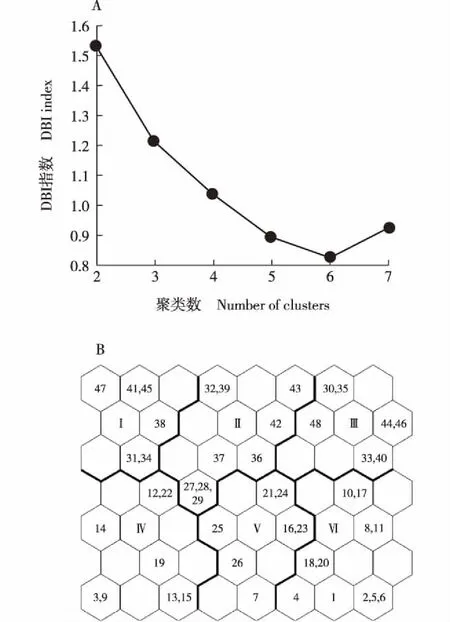
Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ,Ⅵ: 群落类型Community type; 1-48: 样方编号No. of quadrat.图1 云南鸡足山山桂花和野桂花群落48个样方分类的戴维森堡丁指数(DBI)(A)和拓扑映射图(B)Fig. 1 Davies-Bouldin index (DBI) (A) and topology map (B) of classification of 48 quadrats of Osmanthus delavayi Franch. and O. yunnanensis (Franch.) P. S. Green communities in Jizu Mountain of Yunnan Province
6个群落类型在拓扑映射图上界限明显,构成各群落类型主体的植物种类及各群落类型包含的样方如下:
1)群落类型Ⅰ的群落主体由山桂花+绒毛杜鹃(RhododendronpachytrichumFranch.)-大花醉鱼草(BuddlejacolvileiHook. f. et Thoms.)-火绒草〔Leontopodiumleontopodioides(Willd.) Beauv.〕构成,包含样方31、34、38、41、45和47。
2)群落类型Ⅱ的群落主体由山桂花+川滇高山栎(QuercusaquifolioidesRehd. et Wils.)-高山栎(QuercussemicarpifoliaSmith)+滇青冈-高秆薹草(CarexaltaBoott)构成,包含样方27、28、29、32、36、37、39、42和43。
3)群落类型Ⅲ的群落主体由川滇高山栎+山桂花-木香花(RosabanksiaeAit.)-变豆菜(SaniculachinensisBunge)+藏报春(PrimulasinensisSabine ex Lindl.)构成,包含样方30、33、35、40、44、46和48。
4)群落群落类型Ⅳ的群落主体由野桂花+滇青冈-头状四照花(CornuscapitataWall.)-华南铁角蕨(AspleniumaustrochinenseChing)构成,包含样方3、9、12、13、14、15、19和22。
5)群落类型Ⅴ的群落主体由野桂花+大果冬青(IlexmacrocarpaOliv.)-四川卫矛(EuonymusszechuanensisC. H. Wang)-黄水枝(TiarellapolyphyllaD. Don)构成,包含样方7、16、21、23、24、25和26。
6)群落类型Ⅵ的群落主体由滇青冈+野桂花-乌桕〔Sapiumsebiferum(Linn.) Roxb.〕+野桂花-凹叶景天(SedumemarginatumMigo)+野菊〔Dendranthemaindicum(Linn.) Des Moul.〕构成,包含样方1、2、4、5、6、8、10、11、17、18和20。
2.3.2 多元回归树(MRT)检验 MRT采用二歧式分割法将山桂花和野桂花群落的48个样方分为6类,交叉验证符合“1-SE”原则。从6个环境因子中选择分异较大的海拔和坡度作为分类节点用于构建MRT,分类节点分别为海拔2 815、2 605、2 370和2 045 m以及坡度31.4°。6个群落类型的样方数分别为6、9、7、8、7和11(图2)。
MRT输出层为6×8的结构图,48个样方在输出图左下角开始按“之”字形排列,编号为1~48,分为6个群落类型(图3-A)。参考赖江山[32]的方法,根据群落空间分布的连续性,对分类结果进行修订,结果见图3-B。修订样方为26和31,占总样方的比例较小,基本不影响MRT的主体分类结果。修正后6个群落类型分别包含6、9、7、8、7和11个样方,
各样
方的分布和环境因子与SOM分类各类型包含的样方数目和特征一致,说明SOM对环境因子和群落物种重要值的分类排序结果可靠。
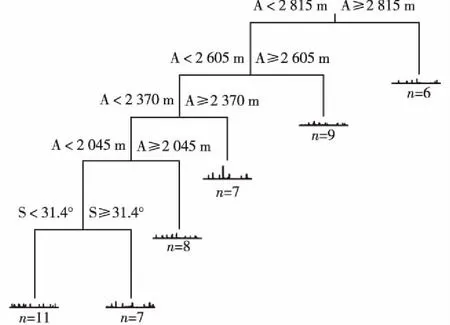
A: 海拔Elevation; S: 坡度Slope; n: 样方数 Number of quadrats.图2 云南鸡足山山桂花和野桂花群落48个样方的多元回归树分类图Fig. 2 Multivariate regression trees classification figure for 48 quadrats of Osmanthus delavayi Franch. and O. yunnanensis (Franch.) P. S. Green communities in Jizu Mountain of Yunnan Province
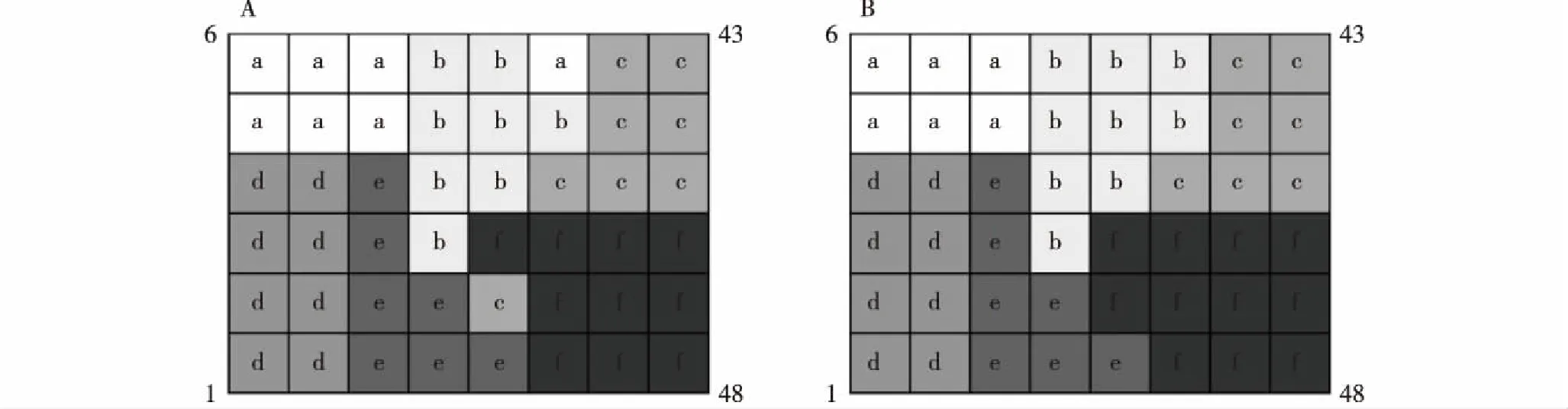
样方1~48从左下角开始按“之”字形排列Quadrat 1-48 are arrayed from left bottom according to zigzag shape. a,b,c,d,e,f: 群落类型Community type.A: 修正前Before modification; B: 修正后After modification.图3 云南鸡足山山桂花和野桂花群落6个群落类型样方分布图Fig. 3 Distribution map of quadrats with six community types of Osmanthus delavayi Franch. and O. yunnanensis (Franch.) P. S. Green communities in Jizu Mountain of Yunnan Province
2.4 山桂花和野桂花群落环境因子聚类
云南鸡足山山桂花和野桂花群落环境因子SOM排序图见图4。由图4可见:SOM排序图纵轴基本呈现了气候和环境在垂直方向上的变化,从下到上由暖湿逐渐向干冷的梯度变化,表现海拔、土壤含水量、土壤厚度、坡度和坡向的垂直变化;横轴反映了坡位的变化过程,从左下至右上坡位从下坡位逐渐变为上坡位。SOM排序图从下到上,海拔逐渐升高,土壤含水量逐渐降低,坡度逐渐增大,土壤厚度逐渐减小,最厚处位于排序图左下角,坡位从右到左数值逐渐减小,即从下位坡变为上位坡,坡向多为阴坡或半阴坡,只有中上部的部分样方处于阳坡或半阳坡。
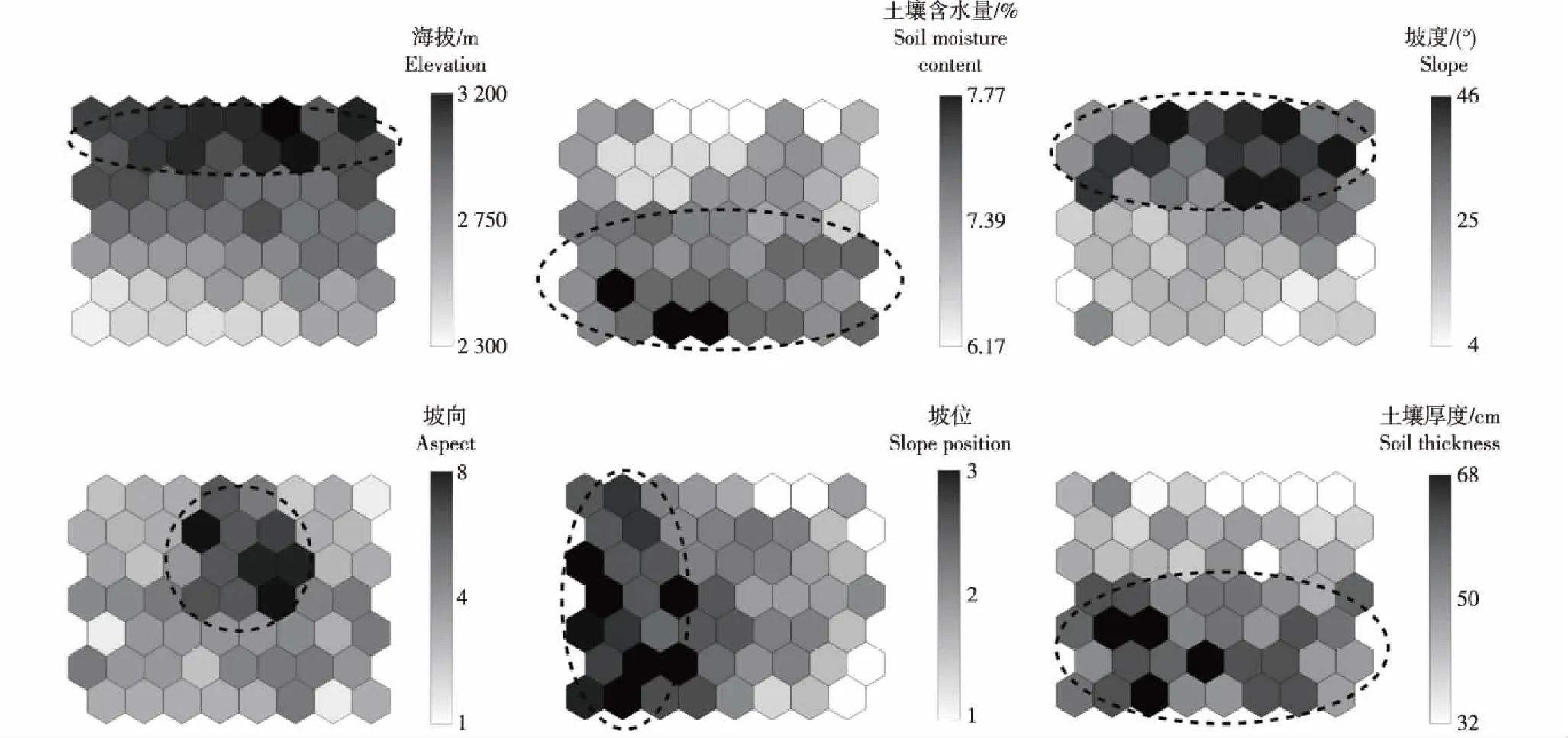
坡向中1、2、3、4、5、6、7和8分别表示北坡、东北坡、西北坡、东坡、西坡、东南坡、西南坡和南坡1, 2, 3, 4, 5, 6, 7, and 8 of aspect represent north, northeast, northwest, east, west, southeast, southwest, and south slopes, respectively. 坡位中1、2和3分别表示上坡位、中坡位和下坡位 1, 2, and 3 of slope position represent the top, middle, and bottom of slopes, respectively. 虚线部分为各环境因子的较高值在拓扑映射图上的集中分布区Broken line part is concentrated distribution area of relatively high value of each environmental factor on topology map.图4 云南鸡足山山桂花和野桂花群落环境因子SOM排序图Fig. 4 SOM ordination map of environmental factors of Osmanthus delavayi Franch. and O. yunnanensis (Franch.) P. S. Green communities in Jizu Mountain of Yunnan Province
2.5 山桂花和野桂花群落的优势种排序
云南鸡足山山桂花和野桂花群落48个样方中优势种在SOM排序图上的梯度分布见图5。结果显示:山桂花、川滇高山栎、绒毛杜鹃和火绒草主要分布于排序图上半部分,分布中心位于群落类型Ⅱ;而野桂花、华南铁角蕨、滇青冈和黄水枝主要分布于排序图的下半部分,分布中心位于群落类型Ⅳ;亮叶杜鹃(RhododendronvernicosumFranch.)、头状四照花和银叶杜鹃(RhododendronargyrophyllumFranch.)的分布则较为均匀,但也有各自的分布中心;大果冬青主要分布于排序图下半部分,分布中心位于群落类型Ⅴ。头状四照花的分布中心对应的是位于群落类型Ⅳ的样方12和22,与该类型的灌木优势种和位置十分吻合。绒毛杜鹃的分布中心对应群落类型Ⅱ的样方36,黄水枝的分布中心对应群落类型Ⅳ的样方3和9以及群落类型Ⅱ的样方27、28和29,大果冬青的分布中心对应群落类型Ⅴ的样方7,说明优势种的分布在很大程度上决定了不同群落类型的范围。
综合图4和图5可以看出:优势种的分布在海拔上两极分化,山桂花是所有高海拔样地的优势种,其伴生种主要为川滇高山栎、绒毛杜鹃、火绒草、亮叶杜鹃以及较少的银叶杜鹃,且伴生种多属壳斗科和杜鹃花科;而低海拔样地则主要以野桂花、滇青冈和华南铁角蕨为优势种,伴生种主要为头状四照花、大果冬青、亮叶杜鹃、银叶杜鹃和黄水枝,且伴生种多属杜鹃花科。山桂花偏好高海拔和较大坡度,而野桂花适宜生长在土壤含水量较高和土壤厚度较厚的环境,不宜生长在坡度过大和海拔过高的环境。黄水枝分布在排序图左侧的样方,只在下坡位以及很少的中坡位有分布,在土壤含水量较低的下坡位分布极少。山桂花和野桂花的分布也一定程度上受到坡度和坡位的影响,环境因子排序图显示从左到右坡位是从下到上,坡度逐渐增高,二者的分布中心偏左侧,说明高坡度和上坡位在一定程度上不利于山桂花和野桂花生长。坡度和坡位对川滇高山栎、绒毛杜鹃和火绒草的影响非常小(可忽略),海拔对这三者的影响较大。

图5 云南鸡足山山桂花和野桂花群落48个样方中优势种在SOM排序图上的梯度分布Fig. 5 Gradient distribution of dominant species in 48 quadrats of Osmanthus delavayi Franch. and O. yunnanensis (Franch.) P. S. Green communities in Jizu Mountain of Yunnan Province on SOM ordination map
3 讨论和结论
群落结构受多种非生物环境因子影响[36-37],如温度、水分和养分等,这3个环境因子在海拔上有综合体现[38]。鸡足山海拔较高,气候季节和垂直变化明显。本研究对该地山桂花和野桂花2个群落物种组成的比较结果表明:山桂花和野桂花群落物种丰富度较高,保护和开发前景较好。野桂花群落植物科、属、种的数量均高于山桂花群落。这可能是由于野桂花群落分布于海拔2 385~2 820 m的常绿阔叶林内,此段山势变化较为平缓,土壤厚度较厚,空间结构合理,而山桂花群落分布于海拔2 810 m的北坡至3 205 m的山顶,此段山势陡峭,土壤贫瘠,由小乔木和灌木组成,层次结构较差,物种丰富度较低。海拔和坡度的差异均导致土壤和植被表现出明显的垂直分布差异,从而影响群落的物种组成[23]。
SOM优势种排序结果说明优势种的分布中心与群落类型分布位置一致,证明优势种是决定群落分布格局的主要因子。48个样方被分成6个群落类型,各类型的优势种和样方间差异明显。山桂花在群落中占明显优势,且在山体上部和山顶均呈聚集分布,推测原因为山桂花群落为高山灌丛,林分密度较低且无遮挡,加之特殊的气候条件,使得山桂花成为绝对的优势种。相反,野桂花则仅在样方1和2中为优势种,通常在群落中位于滇青冈和元江栲等壳斗科大乔木之下,这与赵宏波等[39]对桂花〔Osmanthusfragrans(Thunb.) Lour.〕的调查结果一致。野桂花常呈斑块状分布于常绿阔叶林内,虽然其数量较多且分布均匀,在群落中重要值较高,但由于群落中大乔木所占比例较高,林分密度大,导致野桂花实生苗和根蘖苗较为少见,因此在灌木层中的重要值低于乔木层,且低海拔林窗中物种多样性较高[40],种间竞争较强,其重要值很少存在绝对优势。SOM分类结果显示:6个群落类型在拓扑映射图上界线明显,并且有不同的分布范围和优势。其中,属于山桂花群落的样方29以及属于野桂花群落的样方27和28均被分在群落类型Ⅱ,有一定的重叠,这可能是因为山桂花和野桂花虽然有分布,但3个样方的优势种均是黄水枝和绒毛杜鹃,伴生种也较相似(包括华南铁角蕨、银叶杜鹃和川滇高山栎),加之优势种对群落分类的影响较大,因此被分到同一群落类型。虽然山桂花和野桂花群落间存在海拔落差,但是从群落分类结果来看,2个群落很有可能存在一定交叉,说明二者对环境因子的适应性有相似之处,但是在分布上是逐渐融合还是有远离的趋势还需进一步研究探明。
群落格局受海拔、坡度和坡位等因子的影响较大,其中海拔的影响最大,通过间接影响水热等环境条件来影响高山地区群落格局。本研究调查的6个环境因子中,海拔对山桂花和野桂花群落格局的影响最显著。高山地区温度较低,虽然降水较多,但由于坡度大,水分流失较快,土壤受到冲刷也较为瘠薄,加之坡位和坡向等共同影响高山地区水热资源的分配,使得高山地区对植物的适应性要求较高。山桂花主要在土壤较为贫瘠的高海拔山区分布,此外,坡度和坡向对其分布的影响也较大;野桂花分布的主要影响因子为海拔、土壤含水量和坡位。山桂花、绒毛杜鹃、川滇高山栎和火绒草等种类更适应较为干冷的环境,在高海拔、坡度大的地区是主要优势种;而野桂花、滇青冈、黄水枝和华南铁角蕨等种类要求较为湿润温暖和土层较厚的环境,因而在低海拔地区形成优势种。绒毛杜鹃为喜阳植物[41],受坡向影响较大,其分布中心的坡向为西南坡向,接受的日光照射最多,有利于绒毛杜鹃合成叶绿素和花青素等色素。坡位主要影响黄水枝和华南铁角蕨的分布,二者主要分布在下坡位的非阳面,同时还受到土壤因子的影响,由于这2种草本植物是长在群落近地的阴生植物,对水分和土壤要求较高,且蕨类虽然有根、茎、叶的分化[42],但输导组织欠发达,需要生活在阴暗潮湿的环境,便于吸收水分和无机盐以及孢子萌发,上坡位则不能满足这一条件。坡度和坡位对川滇高山栎和绒毛杜鹃几乎没有影响,说明这2种植物较耐高山贫瘠陡峭的环境,较适应高海拔环境,且其硬革质叶,较耐寒、抗旱,因而占据了高海拔的生态位。
本研究探明了云南鸡足山山桂花和野桂花群落类型的分布格局及其与环境因子的关系,有助于了解这2种中国特有香花木犀属植物的生境情况,为其园林应用和引种培育提供参考。山桂花四季常绿,早春开花,且对土壤水分要求不高,喜高海拔、坡度较陡的上坡位,要求排水良好,对土壤厚度要求较低,是园林绿化的良好树种;而野桂花分布于土层较厚、土壤含水量高、低海拔、坡度较小的下坡位,对水热要求较高。在对山桂花和野桂花的引种栽培和研究中需注意选择适宜的环境及保护自然萌发的根蘖苗,从而保护其遗传多样性。此外,人为活动对其自然演替的影响也待进一步研究。
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
林业勘查设计(2022年1期)2022-02-15 05:35:12
现代园艺(2021年23期)2021-12-01 07:47:44
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
林业调查规划(2020年3期)2020-06-03 07:02:36
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
