
广东惠州莲花山木本植物的多样性和生物量沿海拔的变化及相关性分析
2018-11-23 02:36:20林谕彤李海滨黄潇洒吴林芳朱娘金林永标刘占峰旷远文
植物资源与环境学报 2018年4期
林谕彤, 李海滨, 黄潇洒, 吴林芳, 朱娘金, 林永标, 刘占峰, 旷远文,①
(1. 中国科学院华南植物园 广东省应用植物重点实验室, 广东 广州 510650; 2. 中国科学院大学, 北京 101408;3. 广东惠东莲花山白盆珠自然保护区, 广东 惠州 516003; 4. 广州林芳生态科技有限公司, 广东 广州 510520)
植物的生存以及分布高度依赖于环境,生境不同,植物聚生的种类可能各异[1]。在不同海拔条件下,气温、水分和光照等生态因子的变化对植物生理生态过程,甚至植物群落的组成和分布均具有显著的影响[2],因此,森林群落特征和生物多样性沿海拔梯度的变化成为了植物生态学研究的热点之一[3]。植物种类多样性与海拔主要存在3种关系:一是种类多样性与海拔呈负相关,主要存在于大部分热带湿润地区[4]、温带湿润山区[5]以及部分干旱地区[6];二是随海拔升高,种类多样性指数呈先升高后降低的“中间高度膨胀(mid-altitude bulge)”模式[7],该模式主要受降水和热量影响[8];三是种类多样性与海拔呈正相关[9],存在于极少数地区。研究植物种类多样性的海拔特征对揭示植物分布、共存及其与生态因子的相互关系,了解森林群落动态和维持生态系统结构以及种类多样性保护和管理均具有重要的理论和实践意义。
植被生物量能直接反映植被的生长状况及其对生态因子变化的适应程度[10],是衡量植被生态系统生产力的重要指标之一[11],而探讨生物多样性和生态系统功能的关系是生态学研究的核心内容之一[12]。相关研究结果表明:植物多样性的提高能增加植物生产力和土壤固碳能力[13],但不同生态系统的种类多样性与其生产力存在正相关[14]、负相关[15]或单峰格局[16]等关系。如青藏高原山地植物地上生物量与物种多样性呈负相关,但青藏高原高山草本植物地上生物量与物种多样性则呈“S”曲线模式[17];在青藏高原高寒草甸区,位于中间海拔的植物群落多样性最高,中间海拔以上的群落地上部分生物量随海拔升高而逐渐降低[18];但青藏高原高寒草甸的植物多样性随海拔升高呈显著的单峰模式,植物多样性最高时植物生产力和丰富度均处于中等水平[12];而在热带雨林中,植物种类多样性和生物量随海拔升高呈相反的变化趋势[19]。作为陆地生态系统的重要组成部分,亚热带常绿阔叶林森林生态系统具有重要的生态、社会和经济价值,当前,对亚热带常绿阔叶林的植物种类多样性和生物量随海拔的变化趋势以及二者间的关系尚不明确。
广东惠州的莲花山白盆珠省级自然保护区属南亚热带季风气候,海拔高差大且土壤发育呈明显的垂直带谱,从低到高分布着沟谷阔叶林、针阔混交林、常绿阔叶林、半落叶阔叶林和灌丛草甸森林植被,保护区内保存有完好的南亚热带次生常绿阔叶林和针阔混交林,保护区山脉为粤东沿海第一道生态屏障。目前,对莲花山植被的研究主要侧重于蕨类分布特征[20]、观赏植物资源保护[21]和兰科(Orchidaceae)植物多样性分析[22]等方面,对区内木本植物的种类多样性和生物量分配及其与海拔的关系尚缺乏深入和系统的研究。鉴于此,作者对莲花山不同海拔样地的木本植物种类多样性及生物量变化进行分析,以期为南亚热带常绿阔叶林生态系统的保护、管理及生态系统结构变化的预测提供科学依据。
1 研究地概况和研究方法
1.1 研究地概况
研究地位于广东惠州的莲花山白盆珠省级自然保护区,具体地理坐标为北纬23°02′44″~ 23°11′10″、东经115°02′02″~115°15′38″,主要保护对象为南亚热带次生常绿阔叶林。该区域属南亚热带季风气候,年平均气温22.0 ℃,年均降水量1 935.7 mm,常年基本无霜,气候垂直差异大。莲花山呈东北—西南走向,主峰海拔超过1 300 m,山地地势东北高,坡度较大,西南部为谷地;土壤发育具明显的垂直带谱,从低到高依次为赤红壤、山地红壤、山地黄壤、山地灌丛草甸土。
1.2 研究方法
于2016年11月至2017年10月,在莲花山北坡海拔100~900 m区域,沿海拔每升高100 m设置1个面积40 m×60 m的固定监测样地,共9个样地,各样地的位置信息见表1;并在每个样地内设置6个面积20 m×20 m的样方。海拔900 m 以上区域的植被主要为灌丛和草甸,乔木稀有且山体陡峭,山顶有观音石和观音塑像供人瞻拜,人为干扰过大,故未设固定样地。样地设置规则及样地代表性的保证均参照《中国生态系统研究网络观测与分析标准方法:陆地生物群落调查观测与分析》[23]。
对样地内所有胸径(DBH)大于或等于1.00 cm的木本植物个体进行挂牌,测量其DBH、树高和冠幅,记录种类、株数、分枝情况和生长状态以及相对多度、相对频度和相对优势度等,用GPS定位仪对样地的位置信息进行测定。为了减少DBH测量误差,对DBH>5.00 cm的木本植物,用胸径尺(精度0.1 cm)测量;对DBH≤5.00 cm的木本植物,用游标卡尺(精度0.01 cm)测量。
基于上述的样方调查数据,并依据木本植物个体DBH将样地内所有存活的木本植物分为4个径级:Ⅰ(1.00 cm≤DBH≤5.00 cm)、Ⅱ(5.00 cm
对野外无法确定的种类,采集符合种类鉴定要求的标本2份,由中国科学院华南植物园标本馆的植物分类学家进行种类鉴定,凭证标本保存于中国科学院华南植物园标本馆(IBSC)。
表1广东惠州莲花山不同样地的位置信息
Table1LocationinformationofdifferentplotsofLianhuaMountaininHuizhouofGuangdongProvince

样地编号No. of plot海拔/mAltitude纬度Latitude经度Longitude110023°08'15″115°09'13″220023°04'04″115°11'57″330023°04'06″115°12'13″440023°03'56″115°12'31″550023°03'50″115°12'40″660023°03'40″115°12'54″770023°03'39″115°13'06″880023°00'32″115°13'21″990023°03'29″115°13'28″
1.3 数据处理和分析
根据公式“重要值(IV)=相对多度+相对频度+相对优势度”计算重要值,其中,IV值大于0.400的种类为优势种;采用Shannon-Wiener多样性指数(H′)和Simpson多样性指数(D)分析各样地的种类多样性[24];根据公式“个体百分数=(某样地中各径级的个体数量/该样地的个体总数)×100%”计算该样地各径级的个体百分数。根据参考文献[25-26]计算各样地木本植物各器官的生物量,并根据各器官的生物量总和计算各样地木本植物的生物量。
采用EXCEL 2010和SigmaPlot 12.5软件对数据进行处理和分析。
2 结果和分析
2.1 木本植物种类组成及其与海拔的相关性
调查结果显示:莲花山不同样地中共分布有木本植物181种(变种),隶属于54科109属(见附录Ⅰ)。其中,海拔100 m样地为人工马尾松(PinusmassonianaLamb.)+大叶相思(AcaciaauriculiformisA. Cunn. ex Benth.)林,人为干扰严重。海拔200 m样地分布有木本植物72种(变种),隶属于35科58属,优势种为鹅掌柴〔Scheffleraoctophylla(Lour.) Harms〕和水东哥(SaurauiatristylaDC.)。海拔300 m样地分布有木本植物38种(变种),隶属于23科31属,以樟科(Lauraceae)植物为主,优势种为厚壳桂〔Cryptocaryachinensis(Hance) Hemsl.〕、鹅掌柴、肉实树〔Sarcospermalaurinum(Benth.) Hook. f.〕和华润楠〔Machiluschinensis(Champ. ex Benth.) Hemsl.〕。海拔400 m样地分布有木本植物48种(变种),隶属于25科36属,优势种为厚壳桂、华南青冈〔Cyclobalanopsisedithiae(Skan) Schott.〕、木荷(SchimasuperbaGardn. et Champ.)和鹅掌柴。海拔500 m样地分布有木本植物47种(变种),隶属于23科37属,优势种为红花荷(RhodoleiachampioniiHook.)、鹅掌柴、罗浮柿(DiospyrosmorrisianaHance)和密花山矾(SymplocoscongestaBenth.)。海拔600 m样地分布有木本植物47种,隶属于27科37属,优势种为猴欢喜〔Sloaneasinensis(Hance) Hemsl.〕、红花荷和华润楠。海拔700 m样地分布有木本植物44种,隶属于17科28属,乔木层和灌木层层次明显,优势种为密花山矾、红花荷、罗浮柿和罗浮杜鹃(RhododendronhenryiHance)。海拔800 m样地分布有木本植物16种(变种),隶属于11科12属,优势种为鼠刺(IteachinensisHook. et Arn.)。海拔900 m样地分布有木本植物25种(变种),隶属于16科19属,优势种为罗浮柿、密花山矾和罗浮杜鹃。
对莲花山不同样地中木本植物科数、属数和种数与海拔的相关性进行分析,结果见图1(海拔100 m样地因人为干扰严重,数据未纳入)。结果显示:莲花山海拔900 m以下样地的木本植物种类丰富度随海拔升高而降低,其中,海拔200 m样地的木本植物种类最丰富,其次为海拔400 m样地;整体上看,海拔200~700 m样地的木本植物种类较为丰富,海拔800 m以上样地的木本植物种类丰富度最低。从相关系数看,莲花山海拔900 m以下样地的木本植物的科数与海拔极显著(P<0.01)负相关,属数和种数与海拔均显著(P<0.05)负相关,表明海拔对莲花山木本植物的分布有显著影响。
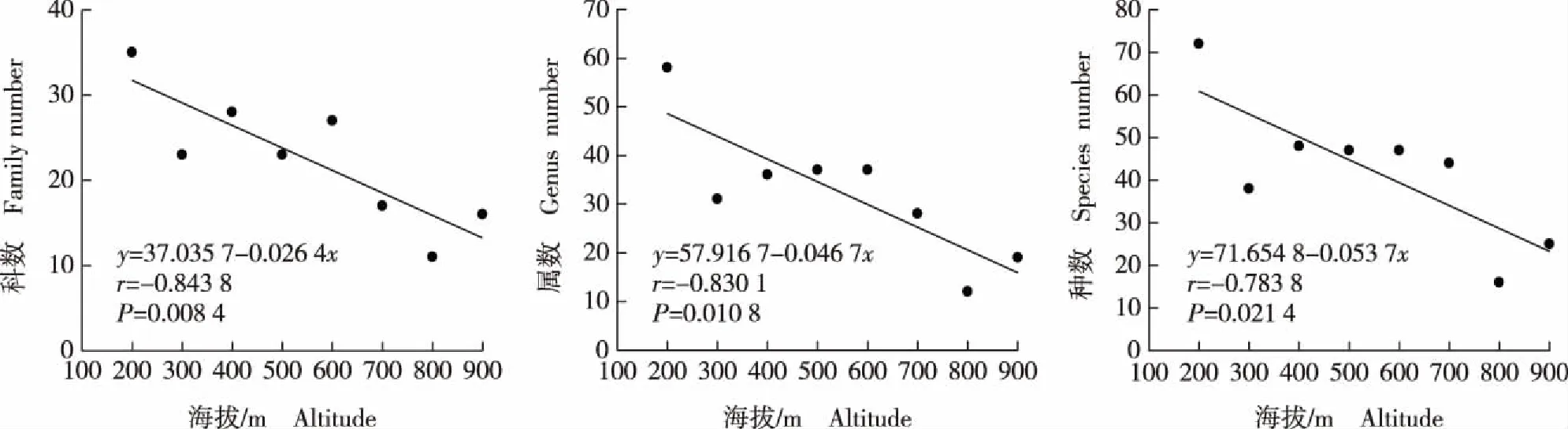
图1 广东惠州莲花山木本植物的科数、属数和种数与海拔的相关性Fig. 1 Correlation of numbers of family, genus and species of woody plant with altitude of Lianhua Mountain in Huizhou of Guangdong Province
2.2 木本植物种类多样性及其与海拔的相关性
莲花山不同样地中木本植物的Shannon-Wiener多样性指数(H′)和Simpson多样性指数(D)见表2;H′和D值与海拔的相关性分析结果见图2。
表2广东惠州莲花山不同样地的木本植物种类多样性1)
Table2SpeciesdiversityofwoodyplantindifferentplotsofLianhuaMountaininHuizhouofGuangdongProvince1)

样地编号No. of plot海拔/mAltitudeH'D1100——22004.6790.98433004.7170.98544005.2460.99255005.0020.99166004.9160.98877004.5310.98088003.8800.97099003.9460.970
1)H′: Shannon-Wiener多样性指数 Shannon-Wiener diversity index;D: Simpson多样性指数 Simpson diversity index. —: 无数据No datum.
由表2可见:在莲花山海拔900 m以下范围内,海拔400 m样地的木本植物种类多样性最高,其H′和D值分别为5.246和0.992,海拔400 m以上样地的木本植物种类多样性随海拔升高以及海拔400 m以下样地的木本植物种类多样性随海拔降低均呈降低趋势,其中,海拔800 m样地的木本植物种类多样性最低,其H′和D值分别为3.880和0.970。
由图2可见:莲花山海拔900 m以下样地的木本植物H′和D值随海拔升高均呈“低—高—低”的抛物线型分布模式,当海拔大于700 m时,H′和D值均明显降低。从相关系数看,H′值与海拔显著(P<0.05)相关,D值与海拔极显著(P<0.01)相关,表明海拔对莲花山木本植物的种类多样性有显著影响。

图2 广东惠州莲花山木本植物种类多样性与海拔的相关性Fig. 2 Correlation of species diversity of woody plant with altitude of Lianhua Mountain in Huizhou of Guangdong Province
2.3 木本植物生物量及其与海拔的相关性
莲花山木本植物生物量与海拔的相关性分析结果见图3。结果显示:莲花山木本植物的生物量随海拔升高呈“低—高—低”的抛物线型分布模式,其中,海拔400 m样地木本植物的生物量最高,为134.68 t·hm-2;海拔800和900 m样地木本植物生物量明显降低,分别为71.43和77.87 t·hm-2。从相关系数看,木本植物生物量与海拔极显著(P<0.01)相关,表明海拔对莲花山木本植物的生物量有显著影响。
2.4 木本植物个体数量及其与生物量的相关性
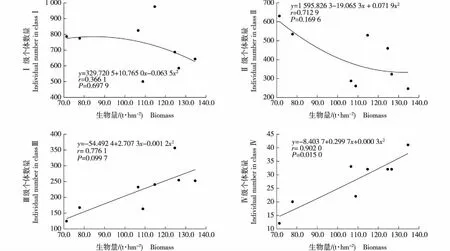
莲花山不同样地中各径级木本植物的个体数量及其个体百分数见表3;各径级木本植物的个体数量与生物量的相关性分析结果见图4。
由表3可见:莲花山不同样地的木本植物中Ⅰ(1.00 cm≤DBH≤5.00 cm)级个体数量均最多。海拔200 m和600~900 m样地中Ⅰ和Ⅱ(5.00 cm
由图4可见:莲花山Ⅰ、Ⅱ和Ⅲ级木本植物的个体数量与生物量均不显著(P>0.05)相关,但Ⅳ级木本植物的个体数量与生物量显著(P<0.05)相关,说明莲花山木本植物的生物量主要受大径级个体数量的影响。
表3广东惠州莲花山不同样地中各径级木本植物的个体数量及其个体百分数1)
Table3NumberandpercentageofindividualofwoodyplantindifferentdiameterclassesofdifferentplotsofLianhuaMountaininHuizhouofGuangdongProvince1)

样地编号No. of plot海拔/mAltitude各径级的个体数量Individual number in each diameter class各径级的个体百分数/%Individual percentage in each diameter classⅠⅡⅢⅣⅠⅡⅢⅣ1100————————22005002601632252.927.517.32.333005853222543249.027.021.32.744006432462524154.420.821.33.555006874593563244.829.923.22.166009755282403254.929.813.51.877008242872323359.920.916.92.488007866301241250.640.68.00.899007745341672051.835.711.21.3
1)Ⅰ: 1.00 cm≤DBH≤5.00 cm; Ⅱ: 5.00 cm
Ⅰ: 1.00cm≤DBH≤5.00 cm; Ⅱ: 5.00 cm
2.5 木本植物种类多样性与生物量的相关性
莲花山不同样地的木本植物种类多样性与生物量的相关性分析结果见图5。结果显示:莲花山木本植物的Shannon-Wiener多样性指数和Simpson多样性指数与生物量均呈极显著(P<0.01)正相关,种类多样性越高,生物量就越大,表明莲花山木本植物生物量受种类多样性影响,种类多样性提高有助于生物量增加。

图5 广东惠州莲花山木本植物的种类多样性与生物量的相关性Fig. 5 Correlation of species diversity with biomass of woody plant of Lianhua Mountain in Huizhou of Guangdong Province
3 讨论和结论
植物分布和生长受诸多生态因子(如温度、湿度和土壤等)的影响。本研究结果表明:莲花山木本植物的科数、属数和种数随海拔升高而显著降低,表明研究区域木本植物的分布具有特定生态幅[27];海拔200~700 m样地的木本植物种类丰富度较高,可能是由于该区域土壤水热条件良好,适宜植物生长;而受降水和温度的影响,海拔800 m以上的区域可能不适合高大乔木生长,而以灌木为主,莲花山木本植物的这一分布特征与亚热带马尾松林相似[28]。
在同一地理范围内,影响陆地植物多样性沿海拔变化的非生物因子主要有土壤养分、水分、光照和温度,生物因子则包括竞争和捕食等[29]。莲花山木本植物种类多样性随海拔升高呈“低—高—低”的抛物线型分布模式,且与海拔显著相关。这种植被分布模式也存在于其他区域,如:白龙江上游地区的杜鹃-巴山冷杉(RhododendronsimsiiPlanch.-AbiesfargesiiFranch.)林和苔藓-巴山冷杉林群落的植物多样性随海拔升高呈“低—高—低”的分布模式[30];华北地区暖温带落叶阔叶林的植物多样性指数在海拔1 200 m以下呈递增趋势,在海拔1 200 m以上呈递减趋势[31];高寒草甸植物群落在中海拔区域的植物多样性最高[18]。低海拔处的人为干扰[30]和高海拔处水热条件的不足[19]可能是导致这种分布模式的原因,不同植物对资源利用效率的差异可能也是导致“中间高度膨胀”分布模式的原因之一[7],如低海拔处优势种对光和水资源的竞争性利用会对其他资源利用效率低的植物产生排斥作用[29]。
除人为干扰外,植物生物量主要受气候因子(如温度、水分和光照等)和地形因子(如海拔)等的影响[10],水热条件的变化是导致植物生物量变化的外在原因,降水量和温度的变化导致土壤水分的变化,并影响植物地上部和地下部生物量的变化[32]。Guo等[16]也认为,不同海拔区域的水热条件差异是影响种类多样性和生产力关系的主要因子。热带森林的高降水量加速了土壤与大气中CO2的交换速率,提高了地上净初级生产力(ANPP)并加快了有机物质的分解速率[33];降水量年际变化是影响中国陆地生态系统生产力年际变化的主要因子,二者之间显著正相关[34]。本研究中,莲花山海拔300~700 m样地的木本植物生物量最大,个体径级大且数量多,且随海拔升高,高大乔木逐渐被罗浮柿和鼠刺等灌木替代,生物量随之降低,海拔对生物量有显著影响。在其他地区[19,35-36]也存在这种植物生物量与海拔的关系模式。
植物多样性与生态系统功能的相互作用是生物多样性研究的核心内容之一[24],且生态系统生产力是生态系统功能的一个重要指标。目前,对植物生物量(生产力)与其多样性之间的关系仍无普遍性认识。有些学者的野外观测和实验分析结果认为,植物种类多样性和生产力存在多种关系,如植物种类多样性增加则生态系统生产力增加[14,37];也有学者认为,植物种类丰富度和生物量的关系呈“驼峰型”[16,38],即随植物生物量增加,水热条件引起植物选择、资源竞争和生境变化,到一定程度时植物种类多样性下降,在未受人为干扰的亚高山地区[39]和高寒草甸地区[18]存在这一现象;还有学者认为,植物的生物量和多样性之间没有明显的关系[40],如随亚热带马尾松林植物种类多样性的增加其生物量没有显著变化[27]。在不同空间尺度上,植物种类丰富度和生产力之间关系不一[1],但无论植物种类多样性与生产力为何种关系,前提条件是排除人为干扰的影响[41]。本研究中,莲花山海拔100~900 m区域木本植物的种类多样性和生物量与海拔均有显著的相关性,与不同生态系统的实测结果和模型预测结果[13-14,37,42]类似。如果考虑各种生态因子的综合作用,植物生物量与种类多样性的关系沿海拔的变化趋势可能更明晰[41]。
综合上述研究结果,莲花山海拔100~900 m样地中共分布有木本植物54科109属181种,在海拔200~700 m样地的木本植物种类丰富度较高。该区域木本植物的种类多样性和生物量随海拔升高均呈“低—高—低”的抛物线型分布模式,在中海拔处(400 m)均最大;其中,Shannon-Wiener多样性指数、Simpson多样性指数和生物量与海拔均显著(P<0.05)相关,Shannon-Wiener多样性指数和Simpson多样性指数与生物量均极显著(P<0.01)相关。莲花山不同海拔区域的水热条件差异可能是其木本植物种类多样性及生物量变化的主要原因。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
作文小学中年级(2021年9期)2021-10-20 12:43:58
河北环境工程学院学报(2021年1期)2021-03-19 08:42:56
现代园艺(2018年1期)2018-03-15 07:56:10
西江月(2017年4期)2017-11-22 07:24:24
科学种养(2017年6期)2017-06-13 08:42:04
学生天地(2016年16期)2016-05-17 05:46:03
小学生作文·小学低年级适用(2016年2期)2016-03-07 11:43:55
