
基于SSR分子标记的青檀自然居群交配系统分析
2018-11-23 02:36范佳佳盛继露李晓红张小平
植物资源与环境学报 2018年4期
范佳佳, 盛继露, 李晓红, 张小平
(安徽师范大学生命科学学院, 安徽 芜湖 241000)
从保护遗传学角度来看,保护物种的遗传和进化潜力是其保护和复壮的重点。交配系统(mating system)是植物种群内2代个体间遗传联系的有性系统,决定了其后代居群的基因型分布,与植物演化密切相关,对植物的分布、群体有效大小、遗传结构、基因流和选择、种群演化等均有影响[1],因此,深入了解植物的繁殖信息对遗传育种方法选择和植物保护策略制定具有重要的指导意义[2]。
青檀(PteroceltistatarinowiiMaxim.)隶属于青檀属(PteroceltisMaxim.)[3],为中国特有的单种属植物,雌雄同株异花,风媒传粉。该树种虽然在中国温带和亚热带地区广泛分布,但由于其对生长条件要求严格,再加上人为干扰的影响,现存种群多呈间断岛屿状分布[4],种群数量和规模呈下降趋势。目前,虽然关于青檀的研究较多[5-7],但尚未见其交配系统类型方面的研究报道,不利于制定其有效保护策略。
SSR分子标记已被广泛用于植物交配系统研究[8]。作者利用12对多态性较高的SSR引物对山东枣庄青檀寺内青檀自然居群的10个家系子代进行了PCR扩增,在此基础上对其交配模式进行了分析,以期分析其交配系统的影响因子,预测其原位保护居群的生存风险,从而为青檀的原位和异位保存以及其遗传多样性保持和繁育体系制定提供科学依据。
1 材料和方法
1.1 材料
在山东枣庄青檀寺内的青檀自然居群中随机选10株青檀作为母本,株距1~50 m;分别采集各母株饱满成熟种子50~100粒,同株种子为1个家系,置于硅胶中带回实验室;采用变温层积法[9]进行催芽,种子发芽后移到花盆中;幼苗具5枚以上真叶时,采集所有叶片,置于硅胶中保存、备用。
1.2 方法
采用改良CTAB法[10]提取10个家系子代237株单株叶片的基因组DNA,置于-20 ℃冰箱中保存、备用。
利用12对多态性较高的SSR引物(表1),采用TP-M13-SSR技术[11]进行PCR扩增反应。扩增体系总体积15.0 μL,包括模板DNA 2.5 ng、2.5 mmol·L-1dNTPs Mix 1.2 μL、25 μmol·L-1MgCl21.2 μL、10×PCR buffer (Mg2+Free)1.5 μL、5 U·μL-1TaqDNA聚合酶0.1 μL、10 μmol·L-1TP-M13荧光引物0.3 μL、10 μmol·L-1正向引物0.4 μL、10 μmol·L-1TP-M13反向引物0.1 μL,ddH2O补足体积。扩增程序:94 ℃预变性4 min;95 ℃变性30 s、相应温度退火30 s、72 ℃延伸30 s, 27个循环;95 ℃变性30 s、53 ℃退火30 s、72 ℃延伸30 s,10个循环;72 ℃延伸8 min。
将质量体积分数1%琼脂糖凝胶电泳检测合格的扩增产物交上海点晶生物科技有限公司进行测序。
表1用于青檀各家系叶片DNA扩增反应的SSR引物序列及退火温度
Table1SequenceandannealingtemperatureofSSRprimersusedforamplificationreactionofDNAfromleafofeachlineofPteroceltistatarinowiiMaxim.

引物编号No. of primer引物序列(5'→3') Primer sequence (5'→3')正向引物 Forward primer反向引物 Reverse primer退火温度/℃Annealing temperatureQT-1CATATTTCCTCTTCCCCTAAACAGCTCACCCATACCTTC55QT-2CACCTTTGCTTACTCCCTGAATGTACTCGCTAATGAACC56QT-3AGCGACTGAGGGTTTCATGGCTTCTGCTCCGCCTTTCT62QT-4CAGGGCACTCCAATAGAATAGATGGTGCTGGGATGGGAAG60QT-5CATTTGGATACACCAGGAAGGCAGCCATTGATGCTTAGTCC60QT-6TCTAGGCTGTATAAAGGGACGATGAAGTAAATGGGGAATC54QT-7CATGTCACCATTACCGAACACACAGTAAGAAAACACACC55QT-8TGGCGATGTGAAGCCCTAAGTCATTTCAACGGTCAAGATTAC56QT-9CAATAATAGCCTTGCATCTCCTCCCTTTGAACAAACCTC56QT-10CCTGTCCAGCTACTAATTTGGTCTGCGATGGTATCTGTT56QT-11CAGGTCCAGAGGGAGAAACCCCAGGGTCAAATAGGTAAT60QT-12GCTTCCTTGGGTCTCATCCTCCACAGACGAGTAGTTCTCC56
1.3 数据分析
以LIZ500为内标,用GeneMarker软件判读扩增产物测序结果,获得微卫星基因型数据,并进行人工校正。基于最大似然法,用MLTR v3.2软件[12]计算交配系统参数,包括单位点异交率(ts)、多位点异交率(tm)、双亲近交系数(tm-ts)和亲本近交系数(F),重复取样1 000次。假定条件:1)每个交配事件为随机的远交(t)或自交(s=1-t);2)异交率与母本基因型独立;3)花粉库与母本同质;4)受精和子代基因型的检测时间无选择;5)各位点等位基因随机分离。近交衰退值(δ)为1减去自交种子相对于异交种子的相对适合度,当不同世代间的亲本近交系数基本不变时,δ=1-2F·t/〔(1-t)·(1-F)〕。
2 结果和分析
实验结果(表2)表明:供试10个青檀家系的双亲近交系数均值为0.028,大于0,表明该青檀自然居群存在一定的近交。
表2基于SSR分子标记的青檀各家系的交配系统参数和近交衰退值
Table2MatingsystemparametersandinbreedingdepressionvalueofeachlineofPteroceltistatarinowiiMaxim.basedonSSRmolecularmarker
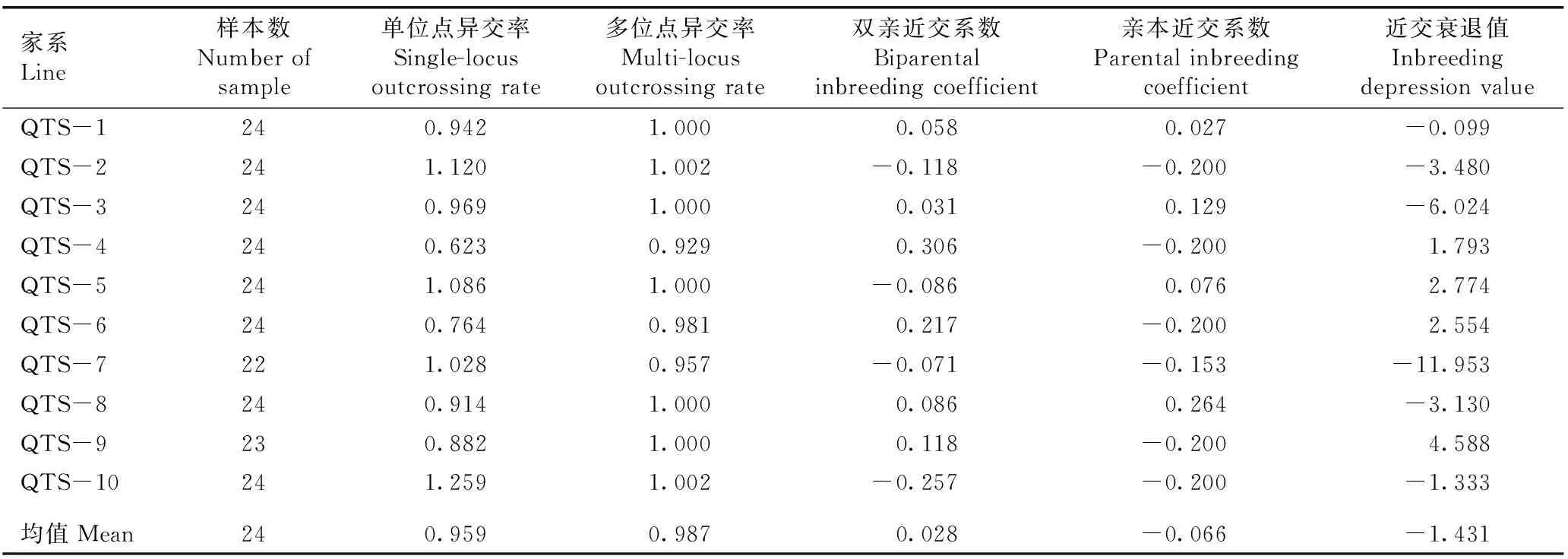
家系Line样本数Number of sample单位点异交率Single-locusoutcrossing rate多位点异交率Multi-locusoutcrossing rate双亲近交系数Biparental inbreeding coefficient亲本近交系数Parental inbreeding coefficient近交衰退值Inbreedingdepression valueQTS-1240.9421.0000.0580.027-0.099QTS-2241.1201.002-0.118-0.200-3.480QTS-3240.9691.0000.0310.129-6.024QTS-4240.6230.9290.306-0.2001.793QTS-5241.0861.000-0.0860.0762.774QTS-6240.7640.9810.217-0.2002.554QTS-7221.0280.957-0.071-0.153-11.953QTS-8240.9141.0000.0860.264-3.130QTS-9230.8821.0000.118-0.2004.588QTS-10241.2591.002-0.257-0.200-1.333均值Mean240.9590.9870.028-0.066-1.431
10个家系的单位点异交率和多位点异交率均较高,其中,家系QTS-2、QTS-5、QTS-7和QTS-10的多位点异交率低于单位点异交率,双亲近交系数小于0,说明这4个家系存在较高比例的异交;其余6个家系的多位点异交率高于单位点异交率,双亲近交系数大于0,说明这6个家系存在亲缘关系较近个体间的异交,即存在程度较低的近交。家系QTS-2、QTS-4、QTS-6、QTS-7、QTS-9和QTS-10的亲本近交系数小于0,说明这6个家系有过剩的杂合子,但缺少纯合子,异交优势较大;而家系QTS-1、QTS-3、QTS-5和QTS-8的亲本近交系数大于0,说明这4个家系有过剩的纯合子,近交优势较大。虽然实际上世代间亲本近交系数不变的情况不存在,但近交衰退值仍能在一定程度上反映各家系的近交衰退程度。计算结果显示:家系QTS-4、QTS-5、QTS-6和QTS-9的近交衰退值较高,分别为1.793、2.774、2.554和4.588。
3 讨论和结论
本研究结果表明:供试青檀自然居群各家系间存在一定的近交(双亲近交系数均值为0.028),这可能是因为青檀的无性生殖能力较强,如根基萌蘖(克隆),由此产生的个体可与母株交配,进而提高自交比例。但是,供试青檀自然居群家系间单位点异交率和多位点异交率的均值均较高(分别为0.959和0.987),这可能是家系间随花粉传播的较强基因流所致,并且,基因流取决于青檀典型的风媒传粉生物学特征(如先叶开花、雌雄同株异花、花药伸出花被片、花粉粒轻且量大、雌蕊柱头呈羽毛状以及不具香味和蜜腺等)。供试青檀自然居群的亲本近交系数均值为-0.066,小于0,说明该青檀自然居群的杂合子过剩。综合上述研究结果及青檀的生物学特征,认为青檀是以异交为主的树种。近交衰退程度、授粉成本和交配机会是影响植物种群自交和异交程度的决定因子[13],并且,较高的近交衰退可能使植物形成适应近交的机制[14]。从生物学特征来看,单性花为青檀避免近交的机制,但雌雄同株却导致这一机制不完全。在供试的10个青檀家系中,有4个家系的近交衰退值较高,因此,虽然该青檀自然居群的杂合子过剩,异交率较高,但是仍需对其各原位保护居群的遗传多样性进行长期监测。
总体来看,防止青檀种群衰退的重要途径为防止近交。首先应对已有种群开展原位保护,设立自然保护区,有效保护适宜青檀生长的石灰岩生境,减少人为破坏和干扰;其次应加强青檀造林技术研究和管理(如引种其他种群的青檀植株,人为加强各种群间的基因交流),提高种群的遗传多样性和活力,最大限度地保存青檀的遗传和进化潜力;再次在引种育苗时广泛收集青檀种子,建立优质种质资源库,尽量种植不同植株或种群的子代,从而更好地开展异地保护。
猜你喜欢
广西林业科学(2022年6期)2023-01-16
植物研究(2022年5期)2022-10-12
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
东方少年·布老虎画刊(2020年3期)2020-06-09
家教世界·创新阅读(2018年9期)2018-12-17
郑州大学学报(医学版)(2015年2期)2015-02-27
