
楠木种群生命表分析*
2018-11-07 05:34:38林金锁
福建林业 2018年3期
林金锁
(邵武市林业局,福建邵武354000)
闽楠(Phoebe bournei),俗称楠木,为中亚热带常绿乔木,是我国江南区域合适的行道树、四旁树、用材树。楠木因其优良的材质,在上世纪中期被大量砍伐,种群数量锐减,群落斑块缩小,天然大面积的野生楠木林难以见到。不仅其种群数量资源日渐减少,其群落生长的天然环境也因人类的放牧、开垦等活动而受到强烈干扰,变得越来越脆弱。加上楠木种子存在休眠,在自然状态下解除休眠的时间较长,种群更新速度慢等原因,加上人为的采伐和干扰,使得楠木的数量越来越少,尤其是野生资源,当前已处于濒临灭绝状况[1-3]。种群是整个生态环境中一个不可缺少的成分,也是对整个生态系统研究的基础。研究种群数量在时空上的动态变化规律,要使用生命表以及损失率曲线等。绘制这一物种生长群落静态生命表的方式可以得到存活率、死亡率和亏损率等重要数据,进一步研究物种集体总数动态变化[4]。本文以建阳麻沙镇楠木林为对象,探讨楠木种群的物种种类、数量、群落生长,并以此提出对应的保护措施,对楠木资源培育和保护等方面都有重要意义。
1 研究区概况
研究地位于建阳市西部,与武夷山市、邵武市交界的麻沙镇,东经116°61′,北纬26°43′,海拔192.63m,面积470km2,其中林业用地3.3万hm2。境内山峰起伏,山沟山谷纵横,溪水河流相互交叉,属亚热带季风气候,年平均气温17.8℃,年均无霜期279d,年均降雨量1490~1720mm,土壤为红壤。
研究区为楠木保护小区,其楠木群落是楠木散布面积最大的一种常绿阔叶林。乔木层主要种类有苦槠(Castanopsis sclerophylla)、米槠(C.carlesii)、虎皮楠(Daphniphyllumoldhami)、冬青(Ilex chinensis)、杜英(Elaeocarpus decipiens)、青冈(Cyclobalanopsis glauca),以及枫香(Liquidambar formosana)和马尾松(Pinus massoniana)等,灌木层有杜茎山(Maesa japonica)、山血丹(Ardisia punctata)、草珊瑚(Sarcandra glabra)、狗骨柴(Diplospora dubia)、山矾(Symplocos sumuntia)、薄叶山矾(S.anomala)等,草本层以狗脊(Woodwardia japonica)为主,层间植物有野木瓜(Stauntonia chinensis)、藤黄檀(Dalbergia hancei)、网络崖豆藤(Millettia reticulata)、酸藤子(Embelia laeta)、络石(Trachelospermumjasminoides)等[5]。
2 材料与方法
2.1 调查对象和方法
采用样方法[6-7]进行调查,在麻沙镇的楠木种群研究区域内设置6块样本林地,大小为20m×20m,总样地面积为2400m2。进行每木检尺,记录下每个样本林地内楠木的数量、树高、胸径等。
2.2 龄级划分
一般情况下静态生命表都是用空间推测时间的方法,以楠木的胸径大小划分等级,将年龄结构替换为立木级结构分析种群动态[1,2]。根据楠木种群的生活史特点,将0~6cm定为第1等级,6~12cm为第2等级,12~18cm为第3等级,以此类推,将数目径级做时间对应关系,第1等级是1龄级的对应,第2等级是2龄级的对应。最后总计出各龄级楠木的株数,从而编楠木种群静态生命表。
2.3 生命表编制
静态生命表一般有以下参数:aα为在α龄级时的单体数;lα为在α龄级开始时标准化存活单体数;dα为从α到α+1龄级期间内标准死亡数;qα为从α到α+1龄级期间死亡率;Lα为从α到α+1龄级期间存活的单体数;Tα为从α龄级到超过α龄级的单体总个数;Tα为进入α龄级单体的生命期望或平均期望寿命;Kα为损失率。表中的所有参数均为互相联结,均可通过公式计算实际值得到,公式如下:
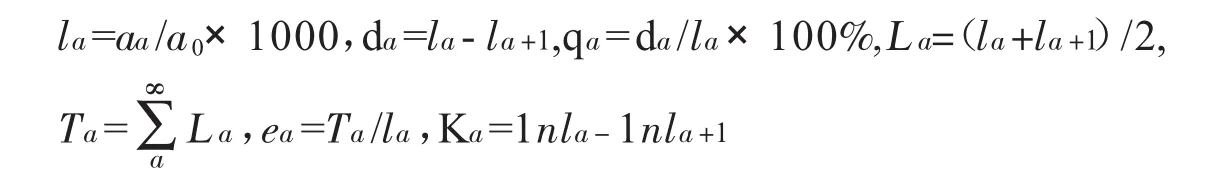
由于静态生命表并非为对这一物种集体的所有生命史的追踪,而是表现多代叠加的动态年龄过程的其中某个时段,且探究方法和行动可能有系统误差,数值记载也可能有误差,在编制过程中会出现亏损率等数值低于正数的状况,所以本文将选取匀滑技术对参数进行处理。
3 结果与分析
3.1 楠木种群生命表
研究区内的楠木林龄跨度很大,以中幼龄楠木为主,也有不少大径级的楠木存活,在保护区外甚至还发现很大龄级的独生楠木(D=200cm,约600~1000a)。楠木生长速率较为缓慢,根据人工速生楠木林生长速率为0.6~0.9cm/a。本研究样地内生长锥调查发现楠木需要生长多年径级才增加1cm(结合文献调查,大致推测楠木胸径年生长约0.20~0.33cm/a,与生长锥调查结果大致相符),因此在划分龄级时跨度较大,大致为1龄级对应大约8~22a,由此确定的楠木种群特定时段生命表见表1。
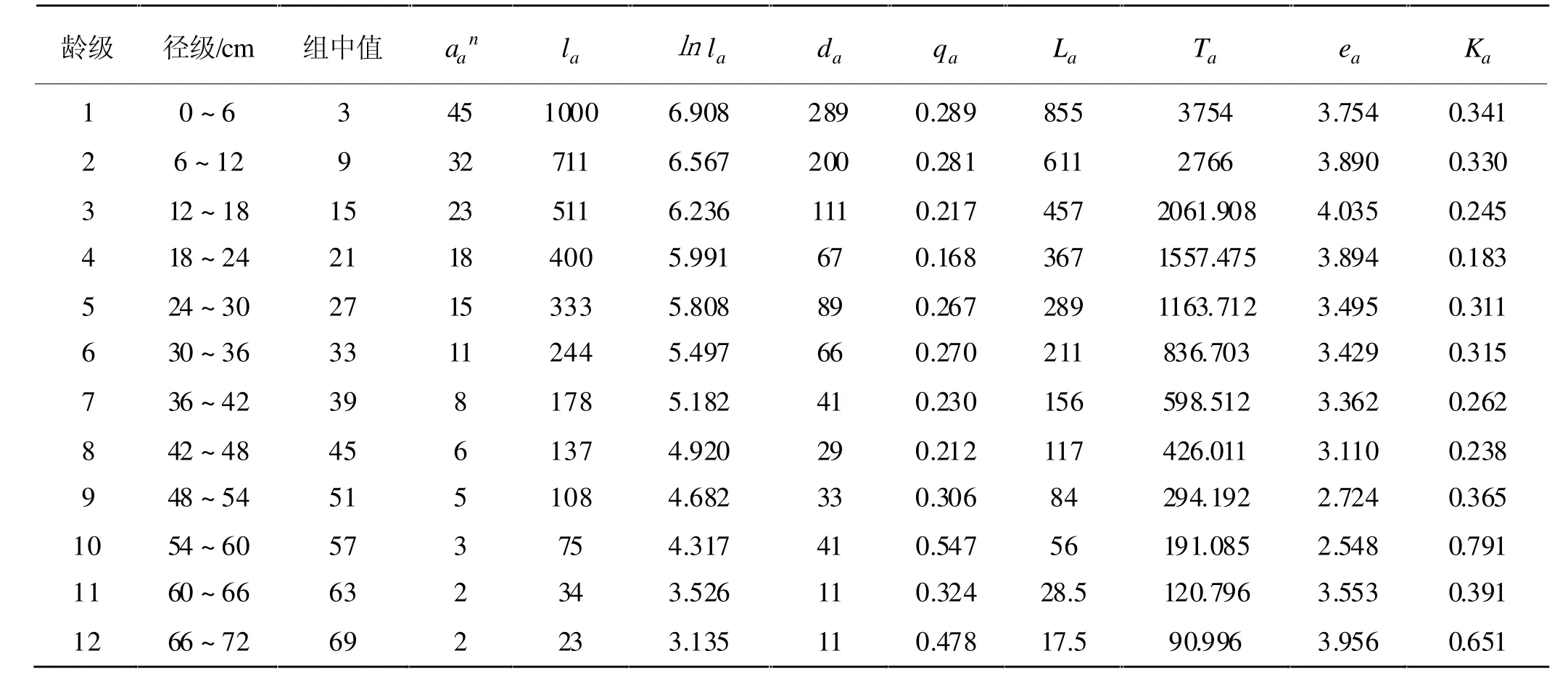
表1 楠木种群的静态生命表
3.2 楠木存活曲线
存活曲线为依靠生命表并凭借生存单体的多寡来表达某个时段死亡率,凭借将某个年龄组的单体存活的值的相对时间绘制成图而获得的表示存活情况的曲线。本文的横轴设定是径级相对年龄,纵轴设定是存活量的对数,绘制楠木物种集体的存活曲线(图1)。
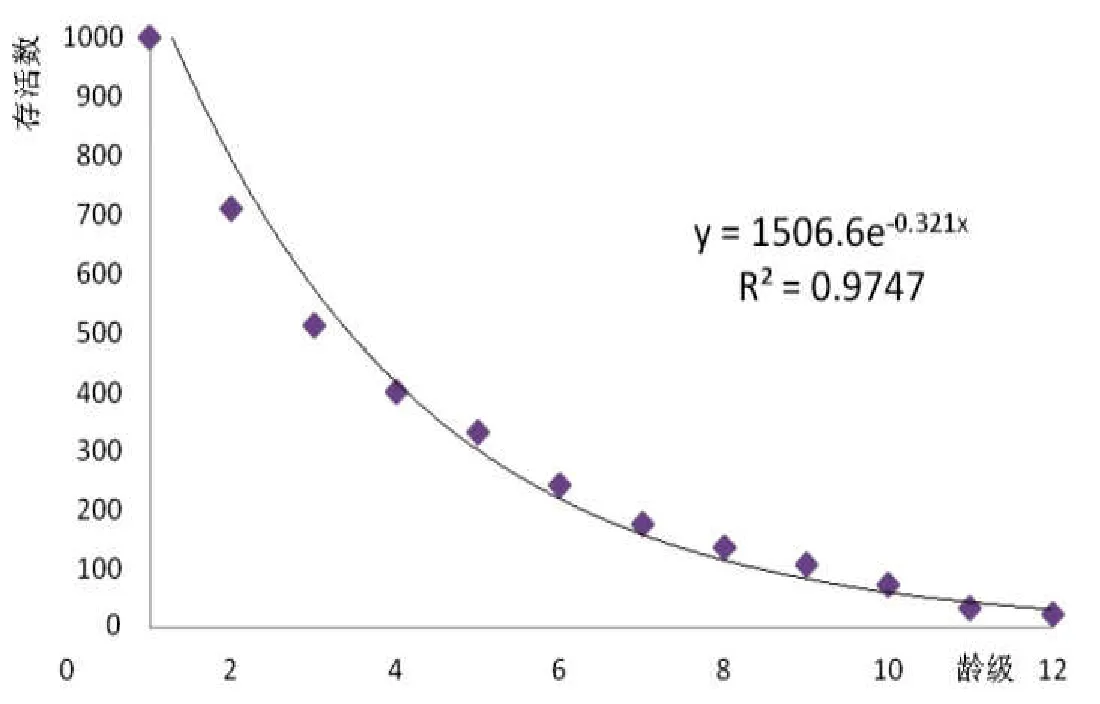
图1 楠木种群存活曲线
由表1和图1可看出,楠木种群的生存情况符合Deevey-Ⅰ型的描述,即凹型曲线,中幼林死亡相对较多,成熟后死亡相对稳定。楠木种群整体生存率较稳定,第1年龄级到第12龄级呈稳定下降趋势,第3~9龄级的生存率有小幅波动,由于直到第12龄级仍然有一定数量的楠木存活,因此无法由本研究确定楠木种群的生命周期及楠木的寿命极限。踏查中发现非样地内有一棵胸径近2m的楠木存活,印证了上述推论。
楠木种群生命表和存活曲线整体上体现了该物种集体的动态变化趋势和年龄层次特征。因为被调查的楠木林的人工干扰总体并不是很大,在存活曲线中出现了些许波动,可能是与当地的生长环境变化有关。
以龄级为横轴,以函数值为纵轴,制作楠木种群的死亡率和损失率曲线(图2)。根据Deevey的区分,存活曲线分为Ⅰ、Ⅱ、Ⅲ3个线型,其中Ⅰ型存活曲线为突起型,表明符合该线型的物种集体只有少量单体不能存活到该物种的生理年龄,一开始死亡率低,生长进入生理年龄后,在较短时段内全部死亡。Ⅱ型存活曲线大致为直线,表达为该物种集体在整个生命周期的死亡率都差不多一样。Ⅲ型存活曲线为下陷型,则该物种种群前期死亡率高,成活到一定的年纪后,死亡率变低。

图2 楠木种群死亡率和期望寿命
从图2可看出,楠木种群在第1~8龄级期间的亏损率总体较低,但有明显的波动,过了第8龄级后亏损率稍微增加;在死亡率部分,第1~8龄级的死亡率较小,可以看出第4龄级时的死亡率比其他龄级低,过了第8龄级后,死亡率开始提高。在楠木种群的存活过程中,随着时间推移,低龄级的楠木对于环境中的空间、光照、水等资源的需求日益增加,但不至于产生种内竞争,所以较高和稳定的存活率,与同一区域中的其他种群产生激烈的竞争,存活了一定的时间后,开始变老,生理机能减退,故死亡率上升。第4龄级的较高存活率,可能与楠木的生长特性有关,需要进一步研究。
在调查中发现,楠木幼树有较好的耐荫性,这与徐振东[8]对于楠木的研究结果一致,但伴随生长进程推进,楠木年龄开始增大,需光量也随之增大,需要全光照成林。整个楠木林中,最大的楠木胸径近2m,通过查找资料,历时千年左右,结合生命表、存活曲线以及死亡率和亏损率曲线,来表明楠木种群在生长周期因为环境选择造成存活数量的稳定减少,到了生理年龄,也就是第8龄级左右,楠木需要通过足够的光照条件才能进入主林层,而在人工干扰较少的楠木林内,只有少数楠木幼株能通过这种筛选,其他植株要么缓慢生长进入主林层,要么就在缓慢生长中逐渐死亡。
4 小结与讨论
通过对福建南平市麻沙镇楠木的地理散布以及该种群静态生命表、存活曲线等数据的分析,楠木物种的生存情况符合Deevey-Ⅰ型的描述,即中幼龄楠木有相对较高的死亡率,而中林龄以后的死亡率较为稳定。楠木的幼树具有一定的耐荫性,较稳定的存活率,由于当地居民的人为干扰,在初期有小幅的下降,直到第4龄级时,因为生产环境受到同类的竞争,需光率上升,需要通过光照条件的严格筛选才能进入上层林层继续成长,存活率开始迅速下降,死亡率急速上升。
在中幼龄期施以适当的人工干扰,达到成熟年龄的楠木可以接受更充足的光照,加强这一种群的自然更新能力,减小死亡率,进而扩展楠木人工林的规模,可以更好的达到保护的目的。
猜你喜欢
人类学学报(2023年5期)2023-04-29 19:50:21
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
运动(2017年1期)2017-09-03 10:39:20
绿色科技(2016年23期)2017-03-15 15:41:56
科教导刊·电子版(2016年25期)2016-11-16 10:24:35
江苏农业科学(2016年5期)2016-07-23 15:59:31
科技视界(2015年26期)2015-09-11 15:21:45
广西农学报(2015年4期)2015-09-11 02:55:46
