
利用同源重组技术构建DKK1、BMP4 真核表达载体及共转染成纤维细胞表达的研究
2018-11-01 08:30张梦瑶刘开东刘积凤贺建宁
华北农学报 2018年5期
张梦瑶,杨 峰,刘开东,刘积凤,贺建宁,柳 楠
(1.青岛农业大学 动物科技学院,山东 青岛 266109; 2.内蒙古农业大学,内蒙古 呼和浩特 010018; 3.青岛畜牧兽医研究所,山东 青岛 266121)
敖汉细毛羊论羊毛的品质比其他羊毛的品质要高,是我国最具有代表性的细毛羊之一[1]。羊的被毛是由毛囊来产生,同时羊毛产量是由毛囊的组织结构和性状共同决定[2]。探究毛囊发育的规律和结构特征,是研究毛囊生长发育调控机制的基础,同时对羊毛性状发育有重要影响[3]。目前,许多毛囊的研究主要集中在出生后以及成年绵羊身上,Stenn等[4]的研究就是针对成年绵羊毛囊的周期性生长变化过程。近年来,柳楠等[5]在分子水平上对敖汉细毛羊羊毛性状的形成机理进行了研究。前人对毛囊的研究大多集中在分子水平上,因此,在细胞水平上探究毛囊生长发育调控的基因已显得非常重要。
在DKK家族中,有一种分泌型的糖蛋白含有2个半胱氨酸[6],且保守区域Cys1和Cys2并肩相存,此蛋白则是DKK1(Dickkopf-1)。它是Wnt信号通路的拮抗物[7],通过抑制LRP5/6与Wnt形成一个三元复杂的跨膜蛋白来对抗Wnt信号,此复杂跨膜蛋白促进LRP5/6的内化,从而抑制 Wnt 信号通路的激活[8]。先前的研究显示[9],毛囊间的距离是由DKK和Wnt信号通过反应-扩散机制(Reaction-dufusion)决定的。DKK1则表达于毛囊发育的起始期,与Wnt信号通过反应扩散机制决定毛囊之间的距离[10]。DKK1在生长期中期表达较弱,在生长期末期开始表达增强[11],DKK1的表达受Wnts的直接控制[12-13]。有转染试验结果表明,DKK1的启动子受经典WNT信号通路调控[14-15]。BMPs也是一种具有分泌功能的蛋白,能使软骨和骨形态发生变化,其成员有很多种[16-17]。有研究表明,BMPs在毛囊发育过程中起到了重要作用,主要表现出对毛囊的抑制作用[18],而它的受体BMPRs类主要表现出对毛囊存在着促进的作用[19-20]。 毛青青等[14]通过对小鼠的研究结果表明,BMP4基因是影响皮肤和毛囊发育的一个关键性基因。BMP4也是TGF-β超家族中的一员[21],在TGF通路中扮演着重要的角色,BMP4的表达不仅对BMP通路中受体进行了控制,也对TGF通路中一些控制生殖发育、细胞增殖、分化的基因进行了控制[10-15]。现单个基因作用的研究已经明确,且与家畜皮肤毛囊关系的研究主要集中在分子水平上,DKK和BMP在毛囊的发育过程中均起到了抑制毛囊发育的作用,若二者同时的表达,表现出相互促进还是拮抗作用有待进一步研究。
本试验通过在细胞水平上研究两基因的相互作用关系,对其做了补充同时可为此基因在个体层次上的研究提供坚实的基础。
1 材料和方法
1.1 试验材料
选取健康40日龄的胎羊(由青岛畜牧所奥特种羊场提供)。
1.2 主要试剂及仪器
SoSo试剂盒购自青岛擎科,pcDNA3.1质粒、NdeⅠ、KpnⅠ限制性内切酶、标准胎牛血清、DPBS等均购自青岛赛尚科贸有限公司。Leica DMIRB 型倒置显微镜购于上海始恒仪器设备有限公司,Redbio CFX ConnectTM荧光定量PCR系统购自Bio-Rad公司。
1.3 质粒构建
1.3.1 引物设计 选取绵羊DKK1基因组(GenBank登录号为XM-012102454.2)CDS区 和BMP4基因组序(GenBank登录号为XM-012180609.2)CDS区,在CDS区外侧通过Primer 5.0设计引物在DKK1基因上下游引物5′端加入同源臂,完成后的引物序列为:
上游引物:5′-TTTAAACTTAAGCTTGGTACCATG
ACGGCTCTGGGCACAGCG-3′;
下游引物:5′-TGGATCCGAGCTCGGTACCTTAGTG
TCTCTGACAGGTGTGAAGCCTG-3′。
在BMP4基因上下游引物5′端加入同源臂,完成后的引物序列为:
上游引物:5′-TTTAAACTTAAGCTTGGTACCAT
GATTCCTGGTAACCGAATGCTG-3′;
下游引物5′-TGGATCCGAGCTCGGTACCTCAG
CGGCACCCACATCCCTCCACTA-3′。
并进行PCR扩增及琼脂糖凝胶电泳检测,扩增体系:cDNA 2 μL,上下游引物各1 μL,Gree Mix 21 μL。
1.3.2DKK1、BMP4基因与pcDNA3.1同源重组 pcDNA3.1质粒分别进行KpnⅠ、NdeⅠ单酶切,酶切体系:KpnⅠ1 μL;pcDNA3.1 5 μL;Buffer 5 μL ddH2O 39 μL;NdeⅠ1 μL;pcDNA3.1 5 μL;Buffer 5 μL ddH2O 39 μL;37 ℃ 2 h,对DKK1、BMP4基因及切开的pcDNA3.1质粒进行胶回收。SoSo连接体系:BMP4、DKK1基因各1 μL,pcDNA3.1 1 μL ,SoSo 5 μL,ddH2O 3 μL,转化DH5α感受态,摇菌,将阳性组送往公司测序,并通过ONZA质粒提取试剂盒获得阳性质粒。
1.4 敖汉细毛羊成纤维细胞培养
取40日龄胎羊用75%酒精和PBS清洗,去除头部和四肢,剩余躯干将其剪成1.0~2.0 mm3大小的组织块,滴加37.0 ℃预热的胎牛血清,放置到CO2培养箱(37 ℃,5.0% CO2)培养4 h,4 h后加入培养液。24 h换新鲜培养液,每天定时在倒置显微镜下观察细胞形态,当细胞密度达到约95%时进行传代。用0.25%的胰蛋白酶37.0 ℃消化,当细胞变圆,加入DMEM终止消化。按1∶3传代,待细胞密度达95%后,再次传代培养。
1.5 细胞转染
传代后细胞密度达到95%用Lipofectamine2000进行瞬时转染。共转染组和单转染组孵育体系:脂质体20 μL,培养液(不含双抗)480 μL,混匀,静置5 min。转染体系:共转染组DKK1、BMP4质粒各20 μL,培养液(不含双抗)460 μL;单转染组DKK1质粒40 μL,培养液460 μL;BMP4质粒40 μL,培养液460 μL,具体步骤参照Lipofectamine 2000说明书。
1.6 转染细胞的定量检测
挑取转染成功的细胞进行培养,待细胞密度达90%时用TRIzol(Roche)试剂裂解细胞提取RNA。后使用核酸测定仪测定浓度和纯度,并反转录成cDNA。根据绵羊DKK1、BMP4基因的全序列和GAPDH的全序列,使用PrimerPremier 5.0软件设计DKK1、BMP4和GAPDH基因的引物序列如表1所示。
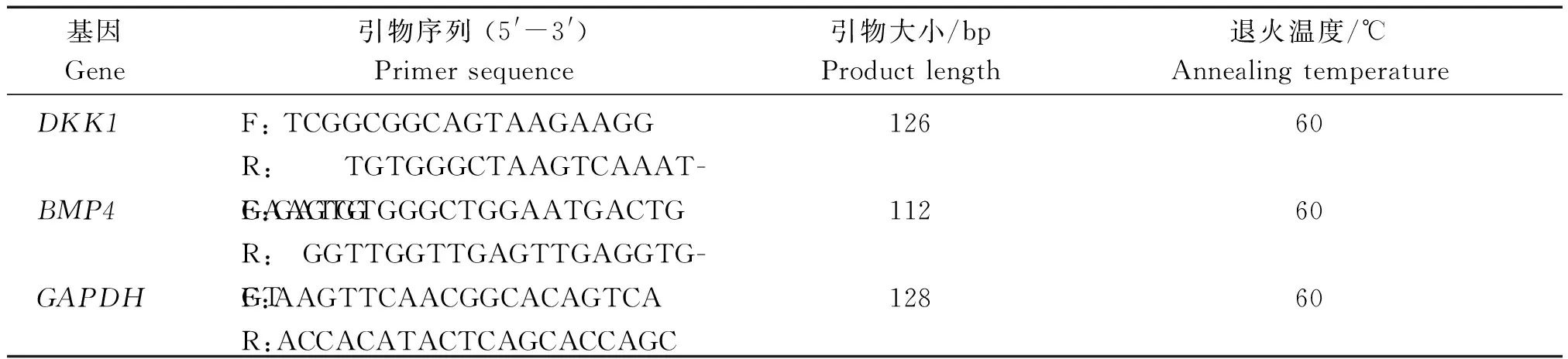
表1 引物信息Tab.1 Primers information
实时荧光定量PCR反应程序:95 ℃预变性7 min;95 ℃变性40 s,60 ℃退火40 s,72 ℃延伸30 s,共45个循环。试验组和对照组样品各进行3次重复,采用Ct(2-ΔΔCt)值法计算BMP4 、DKK1相对于内参基因GAPDH的表达量。利用SPSS 19.0软件中t检验对数据进行差异显著性分析,结果以平均值±标准误表示,以P<0.01作为差异极显著性判断标准。
1.7 Western Blotting检测DKK1、BMP4基因蛋白表达
样品分对照组和试验组。每个样品3个平行。首先用细胞裂解液RIPA提取细胞中的蛋白,β-actin作为内参。对提取的蛋白进行转膜,用5 % BSA对蛋白封闭2 h;将一抗用1∶2 000比例进行稀释,孵育PDF膜4 ℃ 12 h;山羊抗兔的二抗用以1∶2 000的比例稀释,孵育PDF膜1.5 h;曝光液1∶1的比例配置,在暗室中进行曝光拍照。所得结果用ImageJ 软件分析灰度值,灰度值的比值用SPSS统计分析软件进行t检验。
2 结果与分析
2.1 DKK1、BMP4基因PCR扩增
以cDNA为模板,应用设计的上下游引物进行PCR 扩增DKK1、BMP4基因目的片段。2%琼脂糖凝胶电泳分析结果如图1所示,结果表明,引物扩增的条带明亮无杂带,DKK1产物大小为753 bp,BMP4产物大小为1 230 bp,与已知的目的基因大小相同,可用于后期的试验。
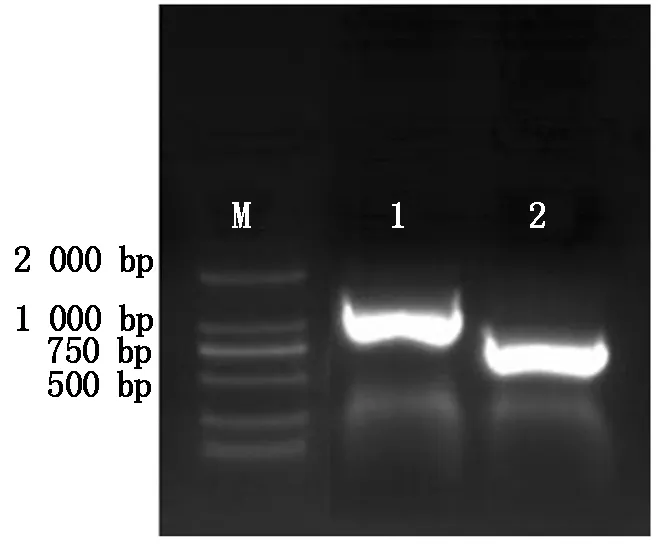
M.2000 Marker;1.BMP4基因扩增条带(1 230 bp); 2.DKK1基因扩增条带(753 bp)。 M.2000 Marker;1.BMP4 product(1 230 bp); 2.DKK1 Product(753 bp).
2.2 pcDNA3.1载体单酶切及纯化
对pcDNA3.1载体分别进行KpnⅠ、NdeⅠ单酶切,单酶切之后的载体进行胶回收用于后续的连接,结果如图2所示,1,4是对pcDNA3.1载体的KpnⅠ单酶切,2,3是对pcDNA3.1载体的NdeⅠ单酶切,pcDNA3.1载体5 428 bp,与图中条带所吻合。因此,1-4可进行目的基因与表达载体pcDNA3.1的连接。
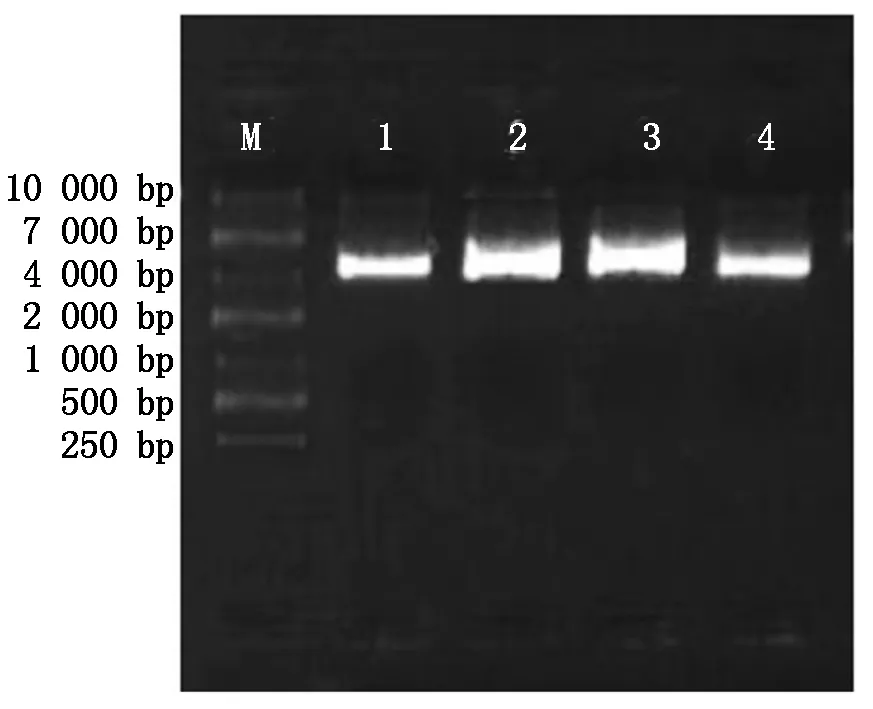
M.10000 Marker;1,4.pcDNA3.1载体KpnⅠ单酶切; 2,3.pcDNA3.1载体NdeⅠ单酶切。 M.10000 Marker;1,4.pcDNA3.1 plasmid KpnⅠ digested; 2,3.pcDNA3.1 plasmid NdeⅠ digested.
2.3 表达载体pcDNA3.1与目的片段重组及鉴定
用SoSo试剂盒将目的片段和pcDNA3.1载体进行连接,转化至大肠杆菌DH5α中,37 ℃过夜振荡培养,对菌液进行PCR电泳鉴定,如图3所示,在753 bp处有单一明亮的条带,为DKK1基因;在1 230 bp处有单一明亮的条带,为BMP4基因。说明目的基因与pcDNA3.1载体连接成功。同时对重组的质粒利用质粒提取试剂盒进行提取,如图4所示,1-2为pcDNA3.1-DKK1重组质粒,大小为6 181 bp,3-4为pcDNA3.1-BMP4重组质粒,大小为6 658 bp。
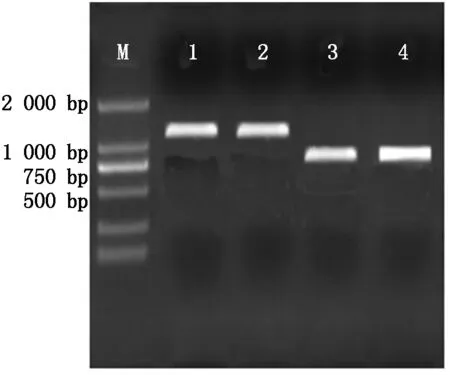
M.2000 Marker;1-2.BMP4菌液PCR; 3-4.DKK1菌液PCR。 M.2000 Marker;1-2.BMP4 microbial PCR; 3-4.DKK1 microbial PCR.
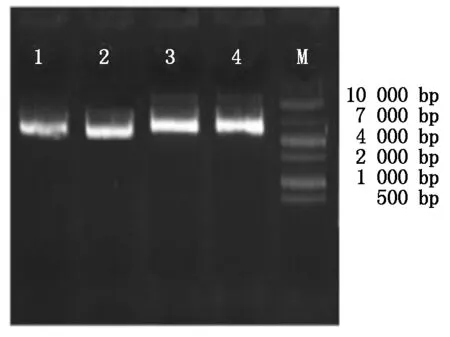
M.10000 Marker;1-2.pcDNA3.1-DKK1重组质粒; 3-4.pcDNA3.1-BMP4重组质粒。 M.10000 Marker;1-2.pcDNA3.1-DKK1 recombinant plasmid; 3-4.pcDNA3.1-BMP4 recombinant plasmid.
2.4 细胞形态观察
用电子倒置显微镜观察成纤维细胞的生长情况,记录成纤维细胞原、传代培养的形状变化。细胞生长24 h可观察到细胞慢慢贴壁,有圆形的,有梭形的,还有一些游离组织。培养至第5天,培养皿皿底基本长满细胞,即可进行传代培养。经传代后细胞开始生长速度稍慢,12 h后增殖迅速加快,细胞形态上仍然呈现梭形。如图5,6所示。

图5 成纤维细胞原代培养(×100) Fig.5 Primary culture of fibroblasts(×100)
图6 成纤维细胞传代培养(×100)Fig.6 Fibroblasts were subcultured(×100)
2.5 TA克隆测序比对
利用同源重组技术将DKK1、BMP4基因各自连接到pcDNA3.1表达载体中,并转化至大肠杆菌DH5α中,摇菌。将菌液送至生工公司进行测序,测序后的序列用Blast进行比对,比对结果如图7所示,在重组的过程中碱基没有发生突变,与已知目的基因完全一致。

图7 DKK1(A)、BMP4(B)TA克隆测序比对Fig.7 DKK1(A),BMP4(B) clone sequencing comparison
2.6 实时荧光qRT-PCR检测DKK1、BMP4基因表达
以cDNA为模板,对目的基因DKK1、BMP4和内参基因GAPDH进行PCR 扩增。2%琼脂糖凝胶电泳分析结果如图8所示,引物扩增的条带明亮无杂带,GAPDH产物大小为128 bp ,DKK1产物大小为126 bp,BMP4产物大小为112 bp,与已知的内参、目的基因大小相同,可用于实时荧光定量PCR试验。
由图 9可知,目的基因BMP4、DKK1和内参基因GAPDH的扩增曲线、熔解曲线清晰整齐,并且目的基因和内参基因的溶解曲线上没有杂乱的峰,表明在实时荧光定量PCR过程中引物特异性很高没有产生二聚体,因此,设计的引物可以用来进行定量分析。采用SPSS软件对单、共转染的细胞mRNA表达量进行分析,BMP4基因的表达量基本无变化,DKK1基因的表达量明显下降,共转染与单转染之间差异极显著(P<0.01)(图10)。
2.7 DKK1、BMP4基因蛋白表达的检测
以成纤维细胞的蛋白为模板进行Western Blot,试验结果显示,在DKK1、BMP4基因单、共转染中比较,DKK1的条带明显暗于单转染的对照组,BMP4的条带与单转染对照组比较亮度基本相同(图11)。经ImageJ和SPSS分析得共转染DKK1基因蛋白的表达极显著低于单转染组(P<0.01),共转染的BMP4基因蛋白的表达量与单转染组相同(图12)。
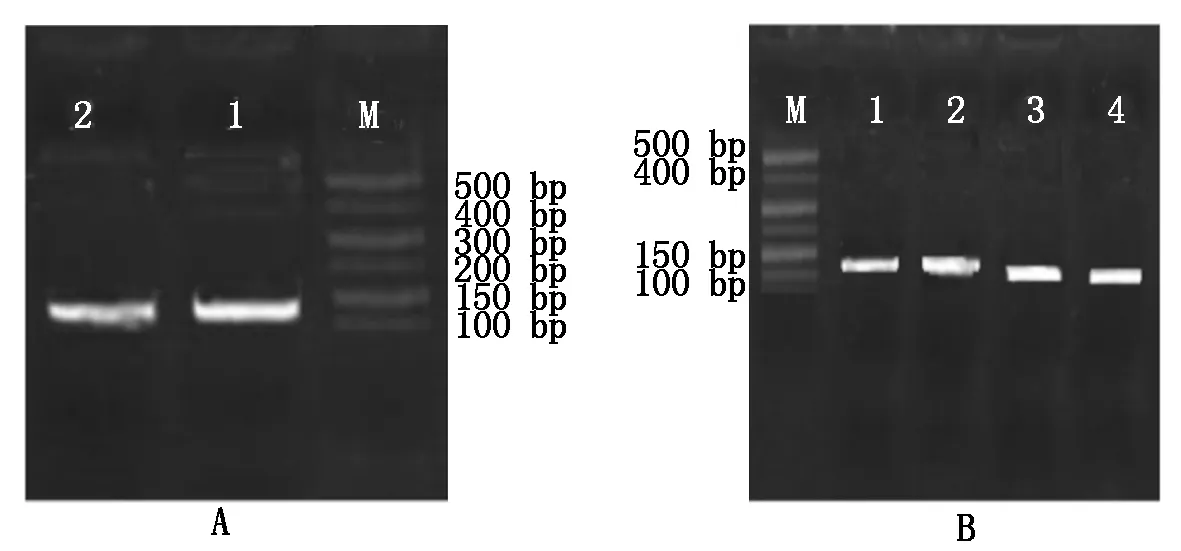
A:M.500 Marker;1-2.内参基因GAPDH扩增条带。B:M.500 Marker;1-2.目的基因DKK1扩增条带;3-4.目的基因BMP4扩增条带。
A:M.500 Marker;1-2. Internal reference geneGAPDHamplification products.B:M.500 Marker;1-2,3-4 .Objective geneDKK1,BMP4 amplification products.
图8内参基因GAPDH(A)目的基因DKK1、BMP4扩增产物(B)
Fig.8InternalreferencegeneGAPDHandobjectivegeneDKK1,BMP4amplificationproducts
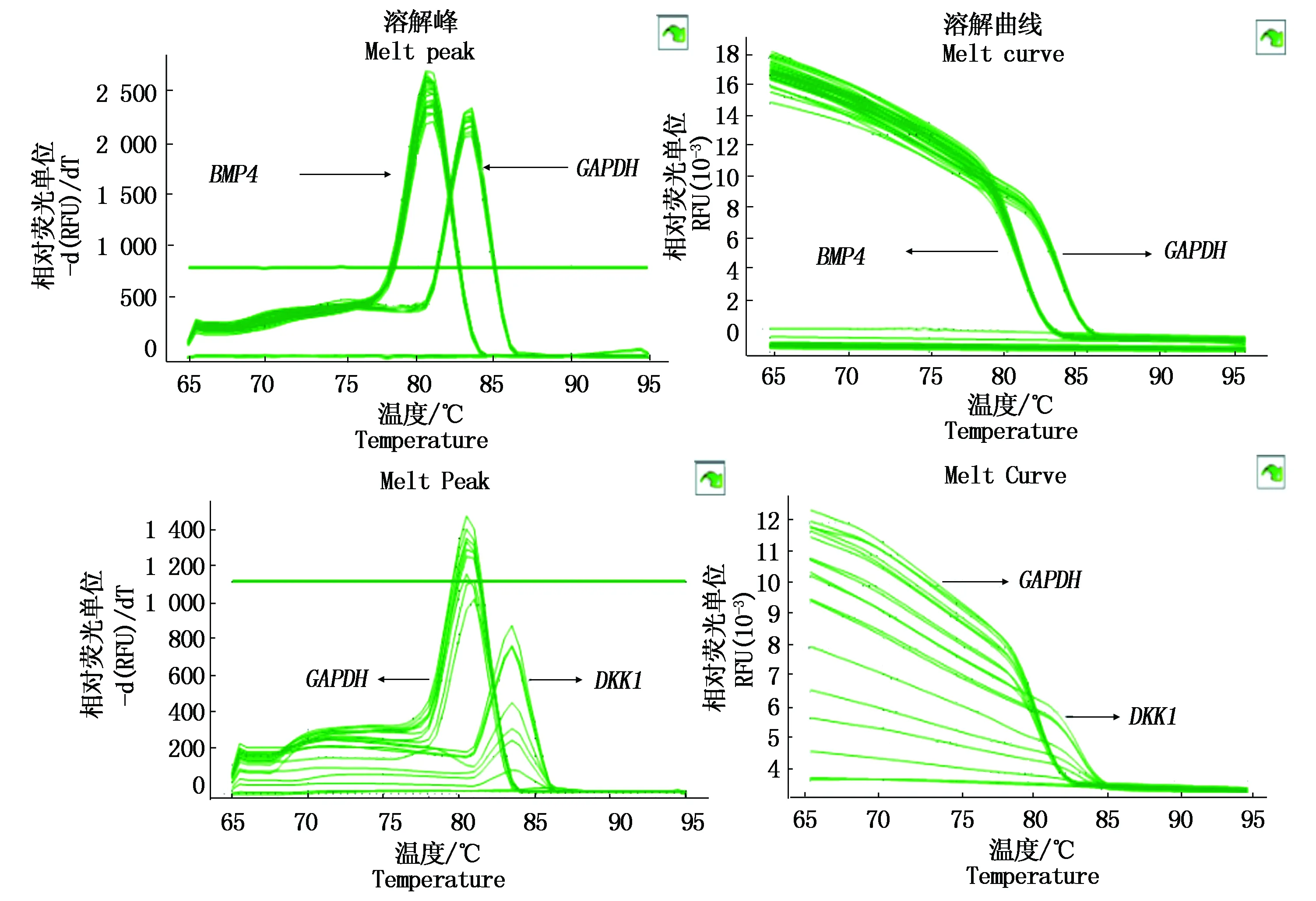
图9 BMP4、DKK1和GAPDH 熔解曲线峰值图(A)和扩增曲线图(B)Fig.9 BMP4,DKK1 and GAPDH peak curve of dissolution curve (A)and amplification curve(B)
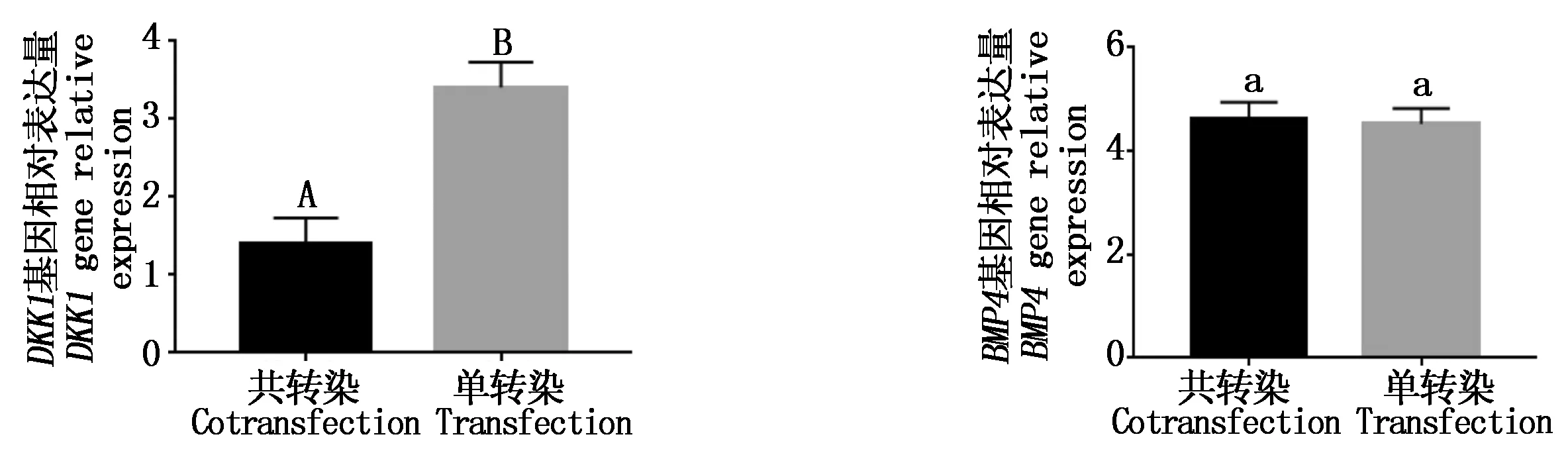
相同小写字母表示差异不显著;不同大写字母表示差异极显著。图12同。 The same small letters mean not significant;Different capital letters mean extremely significant difference.The same as Fig.12.

图11 DKK1、BMP4基因表达蛋白条带 Fig.11 DKK1,BMP4 gene expression protein product
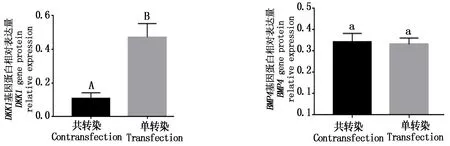
图12 DKK1、BMP4基因蛋白相对表达量Fig.12 DKK1,BMP4 gene protein relative expression
3 讨论与结论
BMP4基因是BMPs中的一员,已有研究表明,BMP4基因是影响毛囊发育的一个关键性基因。有研究结果表明[14],以小鼠为模型研究BMP4在胚胎期毛囊形成和出生后毛囊周期循环过程的毛球间质细胞和表皮里广泛表达[22],说明BMP4基因在毛囊的生长过程中能正常表达。同时将Noggin转入小鼠体内,严重地抑制了BMP4基因的表达,毛发生长异常且形态发生改变。在皮肤组织中利用基因敲除技术特异性敲除BMP4的受体BMPR1A基因的小鼠,毛发的外部形态发生了改变进一步证明了BMP4在毛囊发育过程中的作用[23]。研究证实,Wnt信号通路对于毛囊生长发育起着至关重要的作用。DKK1是Wnt信号通路的抑制剂,DKK1起作用是通过与Wnt信号通路的受体结合来抑制Wnt发挥作用[24]。而在毛囊生长期,过表达DKK1能抑制经典Wnt信号通路,从而促进毛囊外根鞘细胞的凋亡而使毛囊提早进入退化期。之前的研究对DKK1 基因和BMP4基因分别进行单基因的功能鉴定,结果鉴定出DKK1基因对毛囊的发育起到了一定的抑制作用,当其表达量超过一定强度时会抑制羊毛生长发育,导致某些区域羊毛生长不旺盛。而BMP4基因对毛囊的发育也起到了一定的抑制作用,它的过表达也不利于羊毛的生长。当二者进行共转染时,通过基因mRNA的表达量和蛋白表达量的变化来判断二者的相互作用,DKK1的表达量相对于单转染明显下降,而BMP4 的表达量相比于单转染没有发生明显的变化,因而简单地得出,BMP4 基因对DKK1基因起到了一定的抑制作用,二者同时过表达有利于羊毛的生长,以后的羊毛生产过程中可以筛选抑制性基因的过表达。
40日龄的胎羊培养的成纤维细胞生长能力强,活力高,容易获得,结果稳定性更高,试验周期短。因此,本研究选择成纤维细胞作为研究工具,选取子二代来转染,避免了原代细胞中纯度不高,减少了试验的误差。本试验创新点主要是利用同源重组技术构建表达载体,相比TA克隆技术更为简便快速,不会引入多余的碱基,目的基因可以更准确地定位在靶位点上。目前,也尚未发现对DKK1、BMP4基因二者之间作用的研究,特别是在细胞水平上的研究报道,本试验可对此起到补充作用。本研究通过同源重组技术,成功地构建了DKK1、BMP4基因表达载体,将载体转染到成纤维细胞中,采用实时荧光定量PCR、Western Blot来检测基因的表达,从而进一步研究基因的作用。经过单转染的细胞,DKK1、BMP4基因的mRNA和蛋白表达量明显的升高,这从细胞层次上研究了与羊毛有关的基因。经过共转染二者,BMP4基因本身具有抑制毛囊发育的作用,BMP4基因过表达的同时也将影响相关信号通路中DKK1基因的表达。这为以后的个体水平上的基因研究提供了坚实的基础,极大可能地解决了绵羊羊毛生长的问题。
本研究利用同源重组对DKK1、BMP4基因真核表达载体的构建、载体转染成纤维细胞后进行实时荧光定量PCR、Western Blot等,结果成功地构建了pcDNA3.1-DKK1/BMP4表达载体并转染成纤维细胞,转染后细胞生长良好,并且共转染成纤维细胞DKK1基因的表达量明显降低,共转染组的表达量极显著地高于单转染组(P<0.01),BMP4基因共转染的表达量等同于单转染组,因此,BMP4基因抑制了DKK1基因的表达,DKK1基因对BMP4基因的表达几乎没影响,为进一步在个体水平上研究其功能奠定了基础。
猜你喜欢
中国现代医学杂志(2022年16期)2022-08-31
实用医学杂志(2022年11期)2022-07-18
计算机应用与软件(2022年6期)2022-07-12
自然灾害学报(2022年2期)2022-05-10
感染、炎症、修复(2021年1期)2021-07-28
中国学校体育(2021年10期)2021-04-26
中国预防兽医学报(2020年1期)2020-06-05
中国实验诊断学(2019年11期)2019-11-26
文萃报·周二版(2019年37期)2019-09-10
课堂内外(高中版)(2019年8期)2019-09-02
