
水稻苗期低温应答转录组分析
2018-11-01 08:24高红秀何沈雨王思宇张忠臣金正勋
华北农学报 2018年5期
朱 琳,袁 梦,高红秀,曲 悦,何沈雨,王 剑,王 珊, 王思宇,杨 玲,张忠臣,金正勋
(东北农业大学 农学院,黑龙江 哈尔滨 150030)
低温是影响农作物生产的重要危害因素之一。在前期育苗过程中,水稻经常会受到低温影响,导致水稻生长迟缓或停止生长[1-3]。早稻直播面积的逐渐扩大,水稻苗期受低温胁迫的可能性日益增加[4]。因此,提高水稻抗低温能力、深入研究水稻耐低温机制成为许多研究人员关注的热门话题[5]。水稻是世界上最重要的粮食作物之一,全世界超过1/3的人口以水稻为主食[6]。然而,水稻产量受到包括低温等不同非生物胁迫的影响很大[7]。水稻苗期在遭受低温胁迫后会打破细胞渗透压稳态、调节渗透压的物质积累及代谢异常等,导致叶片失绿卷曲、生长缓慢、育性下降和产量降低等[8]。在农业生产中,低温胁迫会致使幼苗枯黄、秧苗腐烂等现象,影响其穗发育和产量等[9]。低温冷害是水稻种植所面临的主要困难之一[10]。目前,关于作物耐冷性研究主要停留在生理阶段[11]。水稻抗逆相关功能基因组学的研究一直是水稻研究核心内容。分析低温胁迫下幼苗体内代谢物变化特征和研究水稻苗期耐低温机理有助于解析水稻应对低温胁迫的生理机制与分子机制,为以后水稻分子育种奠定基础,这对农业经济和社会都有着深远意义。水稻可以通过抵御代谢反应、改变体内激素的含量及活性来调节生理过程,使其在低温条件下能维持正常的生理活动[12-13]。转录水平的调控是生物体最重要的调控方式,与基因组学相比具有针对性强、研究范围小的优点[14]。因此,被认为是一种在转录水平上更精准的测定分析方法[15]。目前,利用高通量转录组测序技术已发现了细枝木麻黄[16]、玉米[17]、西葫芦[18]、油菜[19]等植物与耐低温相关的转录、病害防御、信号转导、转运、次级代谢等大量相关基因。
本研究采用高通量测序方法-转录组测序(RNA-Seq)技术,在转录组水平上研究水稻苗期应答低温差异基因的表达特性,为增强对水稻耐低温机理的认识,明确水稻应答低温的分子机制,以及培育水稻耐低温品种提供参考。
1 材料和方法
1.1 试验材料
供试材料为粳稻品种中花-11,由浙江大学生命科学学院植物生理与生化国家重点实验室提供。
1.2 试验方法
1.2.1 培养条件 种子催芽方法和水培方法参考国际水稻所的水稻水培方法[20]。幼苗长至一叶一心时,移栽到规格为6 cm×6 cm×6 cm的培养钵中,三叶一心时小心将生育进程一致的幼苗取出避免伤到根系。将其中1/2的幼苗放置在光温培养箱(HPG-280,哈尔滨市东联电子技术开发有限公司)作为对照组(WTCK-2),培养条件为:昼夜温度为28 ℃/22 ℃,12 h/12 h,相对湿度为80%;另1/2的幼苗作为低温处理组(WTCOLD-2),移入另一光照培养箱(HPG-280,哈尔滨市东联电子技术开发有限公司),昼夜温度保持在17 ℃,12 h/12 h,相对湿度80%,保持10 d。
1.2.2 RNA提取、文库构建 水稻幼苗在低温处理后,分别取对照和处理后的苗期叶片放入液氮速冻,存至-80 ℃冰箱保存备用。委托北京诺禾致源生物信息科技有限公司完成RNA的提取、质控、建库及Illumina HiSeqTM4000测序工作。
1.2.3 原始数据的组装 由高通量测序(Illumina HiSeqTM)得到的原始数据文件,利用CASAVA碱基识别(Base calling)分析转化成原始测序序列(Sequenced reads),即Raw reads。将低质量和带接头的reads进行过滤得到Clean reads,从而提高信息分析质量,Clean reads将作为后续分析的研究基础。本研究使用Q30(碱基准确率达到99.9%)的质量控制标准。
1.2.4 差异表达基因的筛选 通过使用DESeq自带的标准化方法对原始数据进行标准化处理。在差异分析过程中,使用负二项分布法对Read count的分布进行估计,评估且计算完Pvalue后,对Pvalue进行多重假设检验校正,来减少假阳性。差异基因筛选标准为:|log2(Fold Change)|>1并且校正的P值(Padj)<0.05。
1.2.5 GO基因功能注释分析 使用GOseq[21]将组装后的Unigene序列按照Corrected_Pvalue<0.05筛选差异基因,为寻找出基因显著富集的GO Term,对筛选的差异基因进行GO基因功能注释分析,获得所有差异表达基因的功能注释、生物学功能以及相关的代谢途径,从而推测这些基因对应的特定过程。
1.2.6 KEGG pathway分析 使用软件KOBAS(2.0)进行特定基因的通路定位,找出显著富集的通路,最后对KEGG注释的基因功能信息进行生物学通路的注释与预测,解释低温胁迫下植物对应的调控机制。
1.2.7 聚类分析 使用聚类软件Pheatmap对差异基因的相对表达水平值log2(ratios) 进行聚类。以FPKM值的读数来计算差异表达水平,颜色从红到蓝,表示log10(FPKM+1)从大到小。使用与之匹配的距离算法,计算出基因间的距离,反复迭代,算出基因之间的相对距离,最后依据基因的相对距离远近来分成不同的Subcluster,从而实现聚类。
1.2.8 qRT-PCR验证 利用TRIzol Reagent提取对照组和处理组叶片的总 RNA,cDNA合成利用PrimeScriptTMRT reagent Kit with gDNAEraser(Perfect Real Time),荧光定量检测使用abm®EvaGreen qPCR MasterMix-no dye试剂盒。利用PrimerQuest Tool(http://sg.idtdna.com/Primerquest/Home/Index)设计引物,优先选择靠近基因3′端的引物并进行 Real-time PCR 分析。反应程序为95 ℃,10 min;反应循环数为45次,循环过程是94 ℃,15 s,60 ℃,1 min;72 ℃,20 s。用ABI StepOne Plus(ABI 公司,美国)进行荧光定量检测。利用2-ΔΔCt法,以对照组中各基因表达量为1,Log2FC表示log2倍,误差线表示标准偏差,以OsActin为内参基因对目的基因进行转录水平的相对定量分析。
1.2.9 新基因功能预测 将测序所得的全部reads数据和已知的基因模型进行比较,对比原有的基因注释文件,可以发现新基因。根据转录组数据所提供的新基因碱基序列信息,在MSU(http://rice.plantbiology.msu.edu/analyses_search_locus.shtml)网站上查找相关需要信息。
1.2.10 蛋白互作预测 应用STRING蛋白质互作数据库(http://string-db.org/)中的互作关系分析差异基因蛋白互作网络。利用数据库中提取目标基因集互作关系和目标基因集中的序列,应用BlastX比对到String数据库中包含的参考物种的蛋白质序列上来构建蛋白质互作关系。
2 结果与分析
2.1 低温影响水稻幼苗期的生长发育
17 ℃低温处理10 d后,水稻在正常条件和低温处理下,幼苗长势差异明显(图1)。低温胁迫后的幼苗叶片偏黄、叶尖变黄枯且株高较正常条件下偏低。表明低温胁迫严重抑制了水稻幼苗的生长发育。

图1 正常条件(左)与低温处理(右)下的中花-11表型Fig.1 Zhonghua-11 phenotype under normal condition (left) and low temperature (right) treatment
2.2 测序结果的质量分析
分别收集处理组和对照组水稻苗期叶片并制备其RNA样品。这些样品通过illumina HiSeqTM测序平台进行测序,一共生成约1.01亿条长度约100 bp的原始数据,2种条件下的原始片段分别是47 576 488,53 790 724条。根据生物信息学分析,舍弃低质量的读数,经过序列拼接,最后获得约0.99亿干净的高质量序列,分别得到46 384 074,52 483 052条干净的高质量序列,占原始数据的97.53%。其中,平均Q20、Q30都分别可达到96%,91%以上,GC含量分别为54.91%和55.65%,不确定碱基比例仅为0.02%。使用TopHat2将过滤后的数据定位到中花-11(对照组和处理组)的基因组。在2个样本组的高质量序列读数中,在参考序列上有唯一比对位置的分别占86.29%和84.95%、有多个比对位置的占1.57%和1.75%、总测序序列定位数占87.86%,86.70%。结合以上数据可以说明转录组测序质量较高,符合进一步的生物信息学分析(表1)。
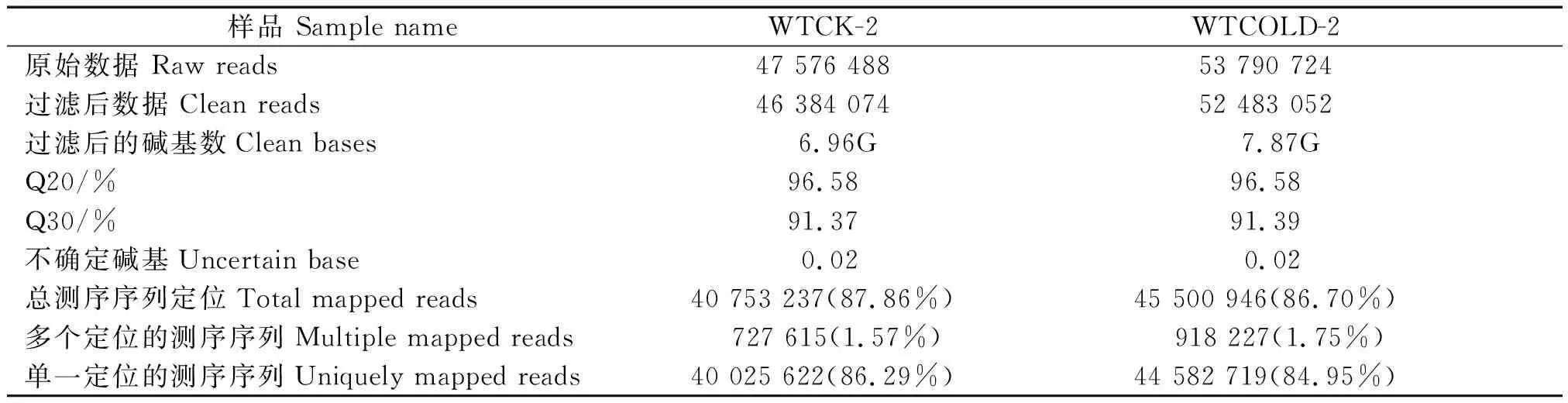
表1 测序数据统计Tab.1 Sequencing date statistics
注:Q20.质量值≥20的碱基所占比例;Q30.质量值≥30的碱基所占比例;GC含量.计算碱基G和C的数量总和占总的碱基数量的百分比。
Note:Q20.The proportion of the bases whose quality value≥20; Q30.The proportion of the bases whose quality value≥30; GC content.Calculate the percentage of the total number of bases G and C as a percentage of the total number of bases.
2.3 差异表达基因的GO功能注释分析
通过GO功能富集分析,对照组与处理组存在2 044个差异表达基因(Differentially expressed genes,DEGs),其中,1 252个基因表达上调,792个基因表达下调。被富集到3 405个GO terms中。其中生物学过程(Biological process,BP)占63.29%、细胞组分(Cellular component,CC)占8.81%、分子功能(Molecular function,MF)占27.90%。通过数据所占比例可以初步判断大多数差异表达基因与一些生物学功能显著相关。单一生物体代谢过程(9.55E-16)、质体(3.94E-21)和阳离子结合(4 .02×10-3)分别是BP、CC、和MF中最显著富集的功能类别。低温胁迫下DEGs涉及的20个最显著富集的功能如图2所示。
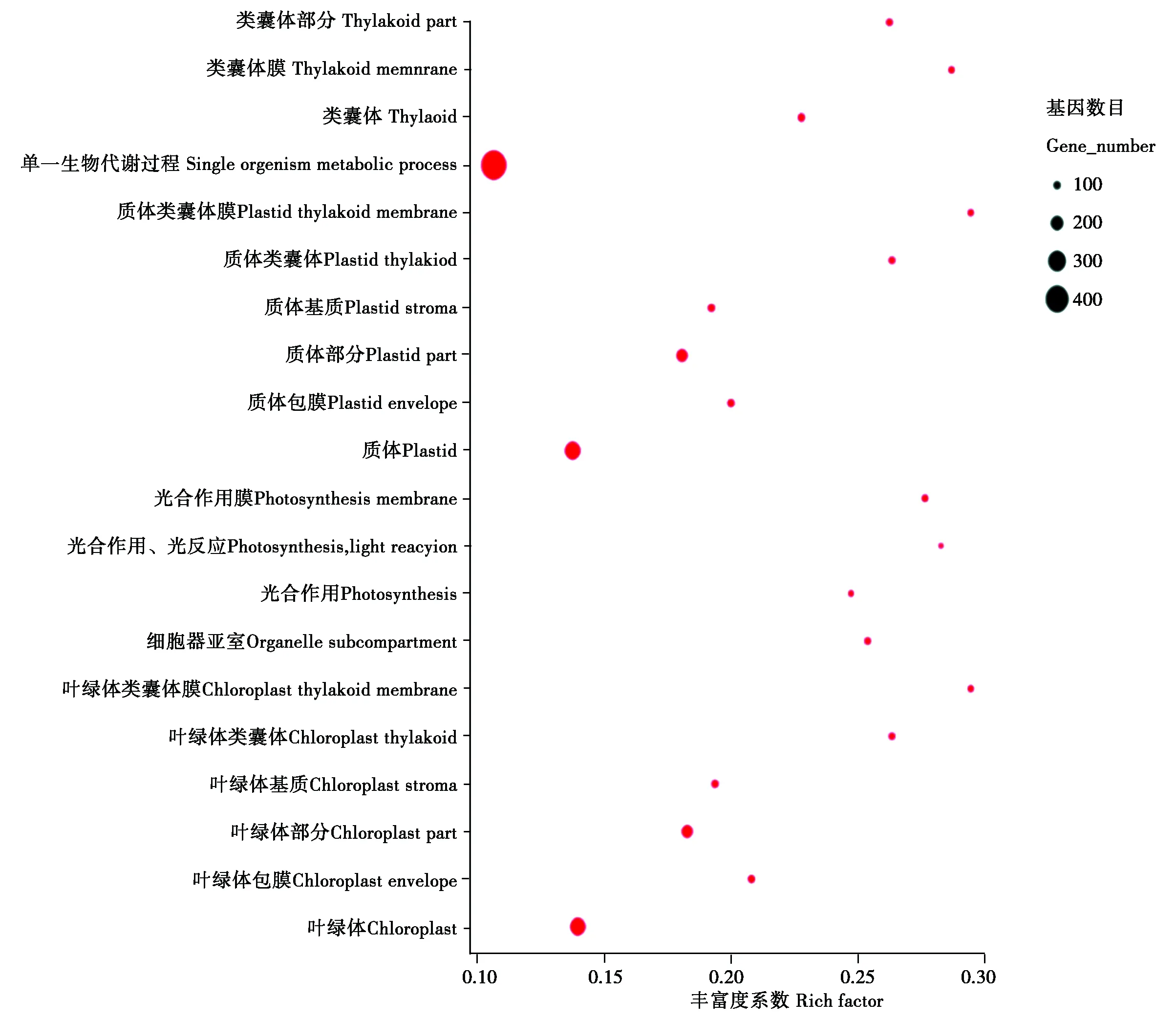
圆点所在的位置代表着富集的条目,圆点的大小表示差异基因的数量。 The position of the dot represents the enriched item,the size of the dot indicates the number ofDifferential genes.
2.4 低温胁迫下差异表达基因的鉴定与筛选
通过将表达模式相同或相近的基因聚集成类,用以推测已知基因的新功能或未知基因的功能[22]。为了鉴定具有功能富集的聚类,利用差异基因的相对表达水平值log2(ratios)进行层次聚类(图3),从图3可以看出,该材料在低温胁迫下基因表达量有变化。
基于以上发现,在对照组和处理组中鉴定了2 044个差异表达基因,其中有关于光合作用、苯丙氨酸代谢、WRKY转录因子等多种功能途径。其中右侧虚线的右侧圆点代表差异表达上调的基因有1 252个,左侧虚线的左侧圆点代表差异表达下调的基因有792个,两侧虚线中间表示无显著性差异表达的基因(图4)。
2.5 差异表达基因的KEGG通路分析
为了精准分析低温胁迫对水稻发育过程中参与的代谢途径发生的变化,对得到的Unigene进行KEGG代谢通路分析(表2)。结果表明,在代谢通路中,主要是光合作用天线蛋白(ko00196)、光合作用(ko00195)、类胡萝卜素生物合成(ko00906)、次生代谢产物的生物合成(ko01110)。利用KEGG数据库对差异基因进行数据分析,得到差异基因KEGG注释通路图,其中水稻苗期对受低温胁迫影响较大的代谢通路分析如下:
通路一:光合作用
低温胁迫下,此通路中的光合系统Ⅰ存在11个差异表达基因,且全部表现为下调;光系统Ⅱ中有24个差异表达基因,其中3个基因表现为上调,21个基因表现为下调;细胞色素b6/f复合体中含有1个差异表达基因且表现为下调;光合电子运输中有20个差异表达基因,有8个基因表现为上调,其余基因均表现为下调;F型ATP酶中有6个表现为下调基因(图5)。上述结果得出,在光合作用通路里这些差异表达基因可能是受低温处理影响较大的基因。
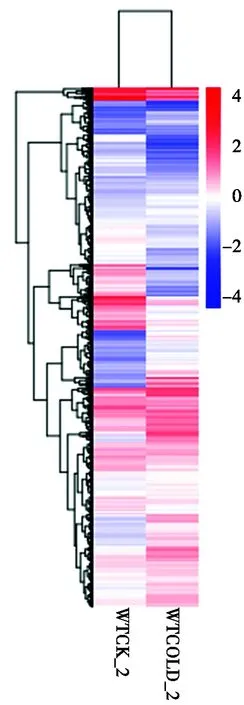
WTCK_2.对照条件下的中花-11;WTCOLD_2.低温胁迫下的中花-11。 WTCK_2.Zhonghua-11 under control conditions; WTCOLD_2.Zhonghua-11 under low temperature stress.
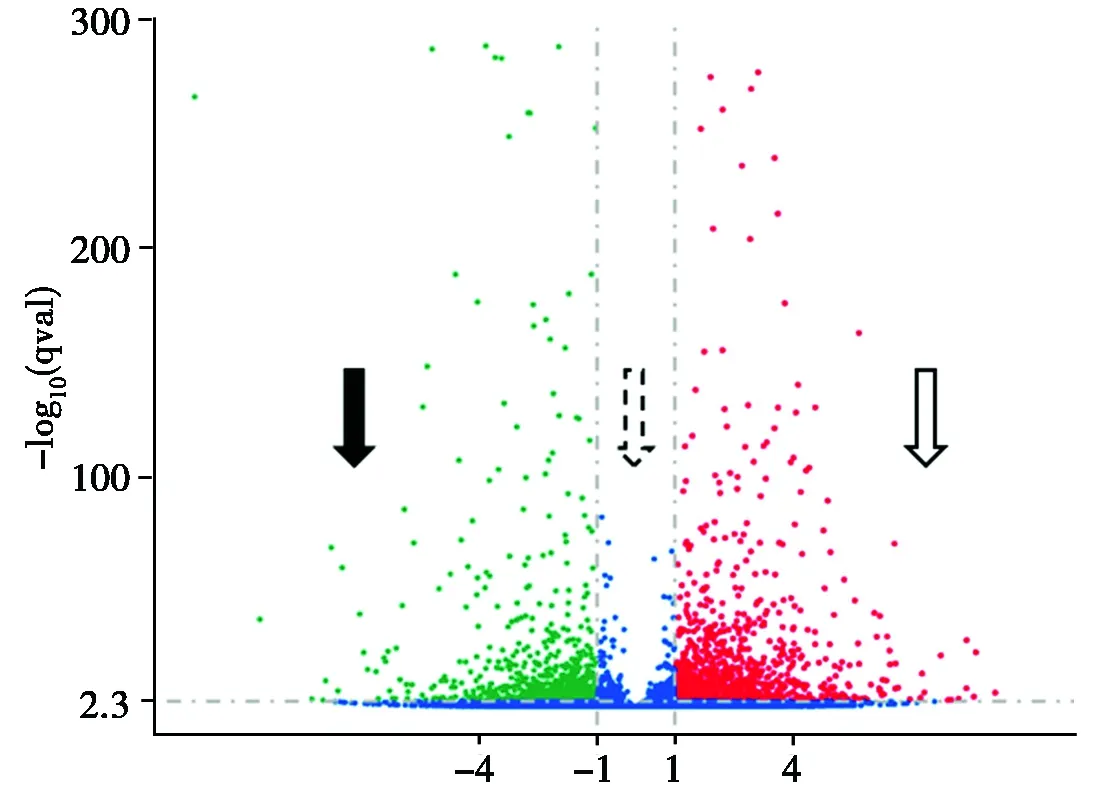
空心箭头.上调的显著差异表达基因;实心箭头.下调的显著差异表达基因;虚线空心箭头.无显著性差异表达的基因;横坐标.基因在不同样本中表达倍数变化;纵坐标.基因表达量变化差异的统计学显著性。
Hollow arrow.Significant Different expressed genes up-regulated; Solid arrow. Significantly Different expressed genes down-regulated; Dotted hollow arrow. Genes with no significant
Differential expression Abscissa. Gene expression fold change in Different samples; Ordinate.Statistically significant difference in gene expression changes.
图4差异基因的火山
Fig.4Volcanicmapofdifferentialgenes
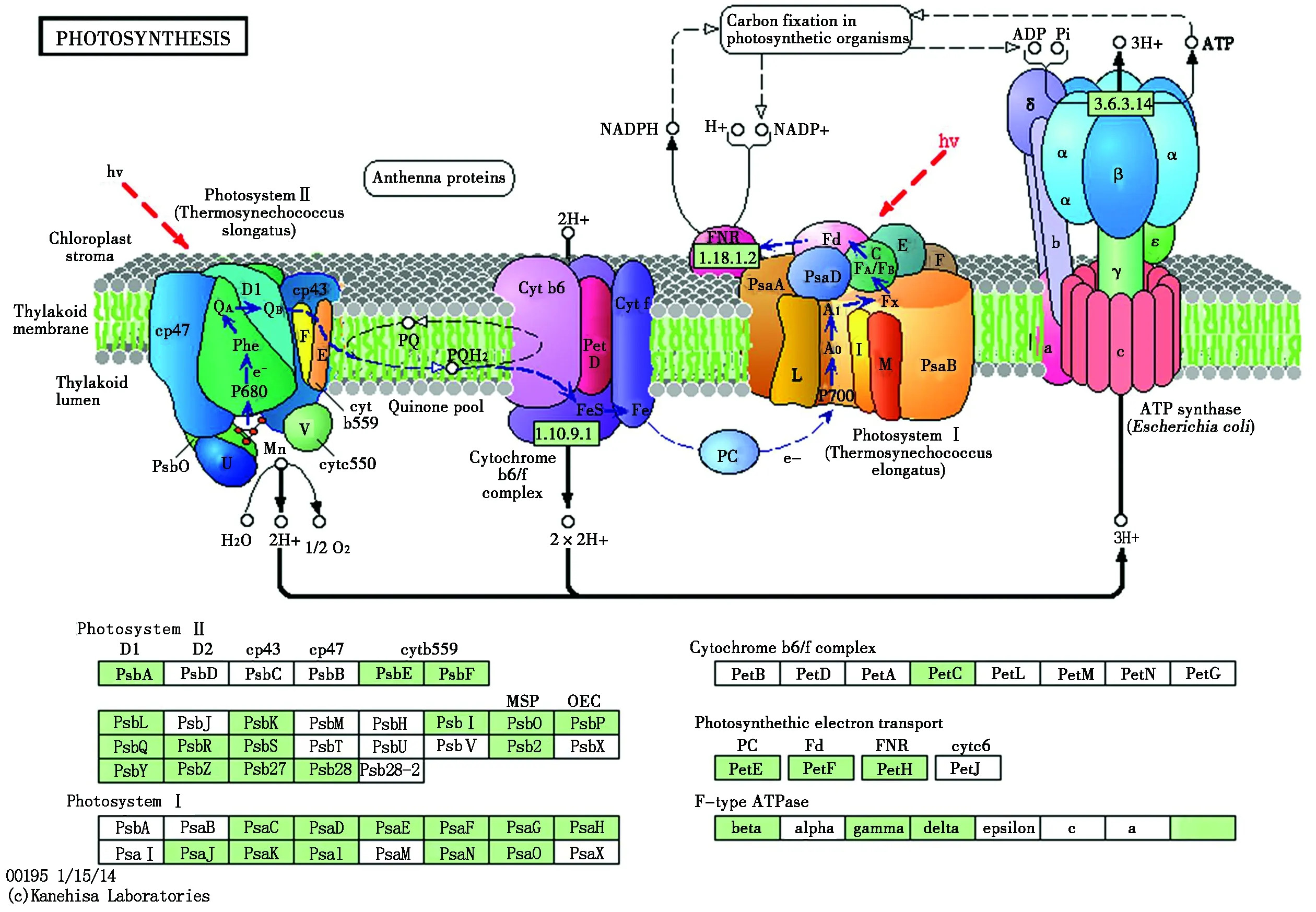
PhotosystemⅡ.光系统Ⅱ;PhotosystemⅠ.光系统Ⅰ;Cytochrome b6/f complex.细胞色素b6/f复合体;Photosynthethic electron transport.光合电子运输;F-type ATPase. F型ATP酶;Carbon fixation in photosynthetic organisms.光合生物中的碳固定;Chloroplast stroma.叶绿体基质;Thylakoid membrane.类囊体膜;Thylakoid lumen.类囊体腔。
图5差异表达基因参与的光合作用代谢途径
Fig.5Photosynthesismetabolicpathwaysinvolvedindifferentiallyexpressedgenes
通路二:苯丙氨酸代谢
苯丙氨酸代谢途径是植物最重要的次生代谢途径之一,苯丙氨酸解氨酶(Phenylalanine ammonia-lyase,PAL)基因表达量高,与之对应的次生代谢产物产量就高,水稻幼苗的整体抗逆能力就强[23-24]。低温胁迫下,在此通路中发现17个差异表达基因,8个DEGs表达上调,其余9个DEGs表达下调。分别在4.3.1.24(K01110)和4.3.1.25(K01110)处PAL相关基因OS02G0626400(1.654 5)、OS02G0626100(1.229 9)、OS04G0518100(1.544 4)上调表达,进而调节水稻次生代谢反应(图6)。从这个通路图可以看出,在低温胁迫下PAL基因表达上调,说明低温与苯丙氨酸代谢通路具有相关性,差异性表达的基因可能是低温应答的关键基因。
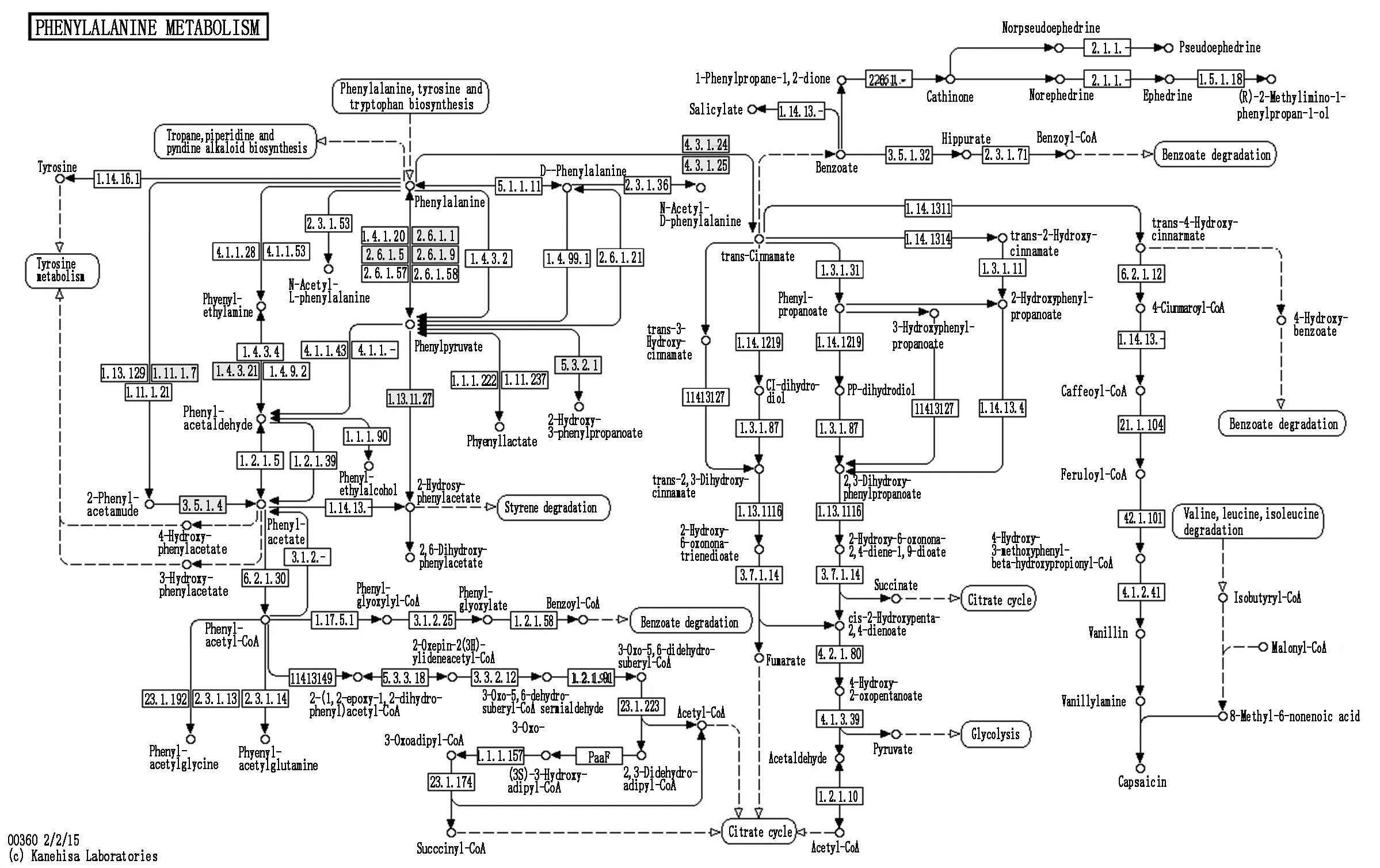
图6 差异表达基因参与苯丙氨酸代谢通路Fig.6 Differentially expressed genes involved in phenylalanine metabolic pathway
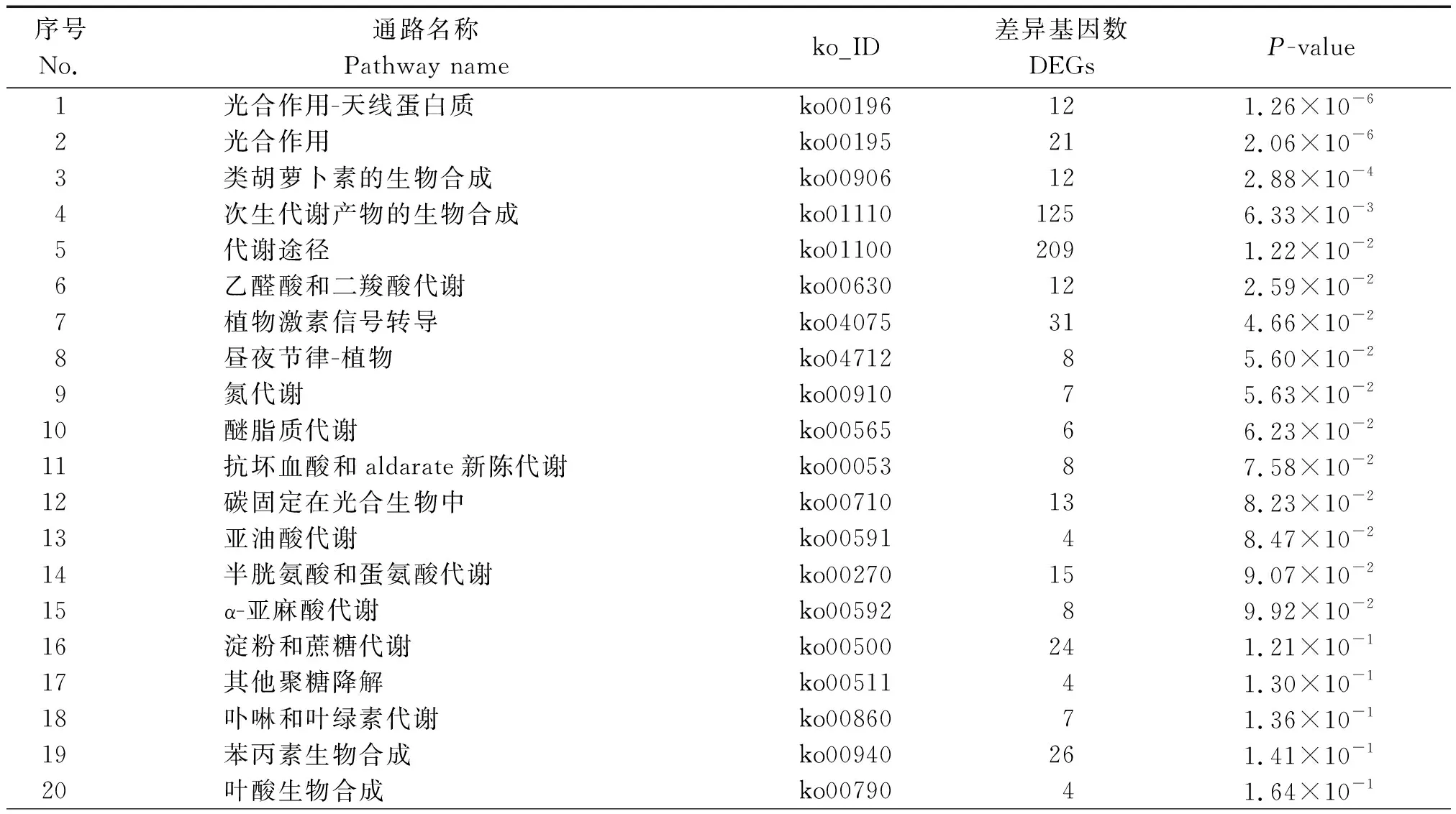
序号No.通路名称Pathway nameko_ID差异基因数DEGsP-value1 光合作用-天线蛋白质ko00196121.26×10-62 光合作用ko00195212.06×10-63类胡萝卜素的生物合成ko00906122.88×10-44次生代谢产物的生物合成ko011101256.33×10-35代谢途径ko011002091.22×10-26乙醛酸和二羧酸代谢ko00630122.59×10-27植物激素信号转导ko04075314.66×10-28昼夜节律-植物ko0471285.60×10-29氮代谢ko0091075.63×10-210醚脂质代谢ko0056566.23×10-211抗坏血酸和aldarate新陈代谢ko0005387.58×10-212碳固定在光合生物中ko00710138.23×10-213亚油酸代谢ko0059148.47×10-214半胱氨酸和蛋氨酸代谢ko00270159.07×10-215α-亚麻酸代谢ko0059289.92×10-216淀粉和蔗糖代谢ko00500241.21×10-117其他聚糖降解ko0051141.30×10-118卟啉和叶绿素代谢ko0086071.36×10-119苯丙素生物合成ko00940261.41×10-120叶酸生物合成ko0079041.64×10-1
表2(续)
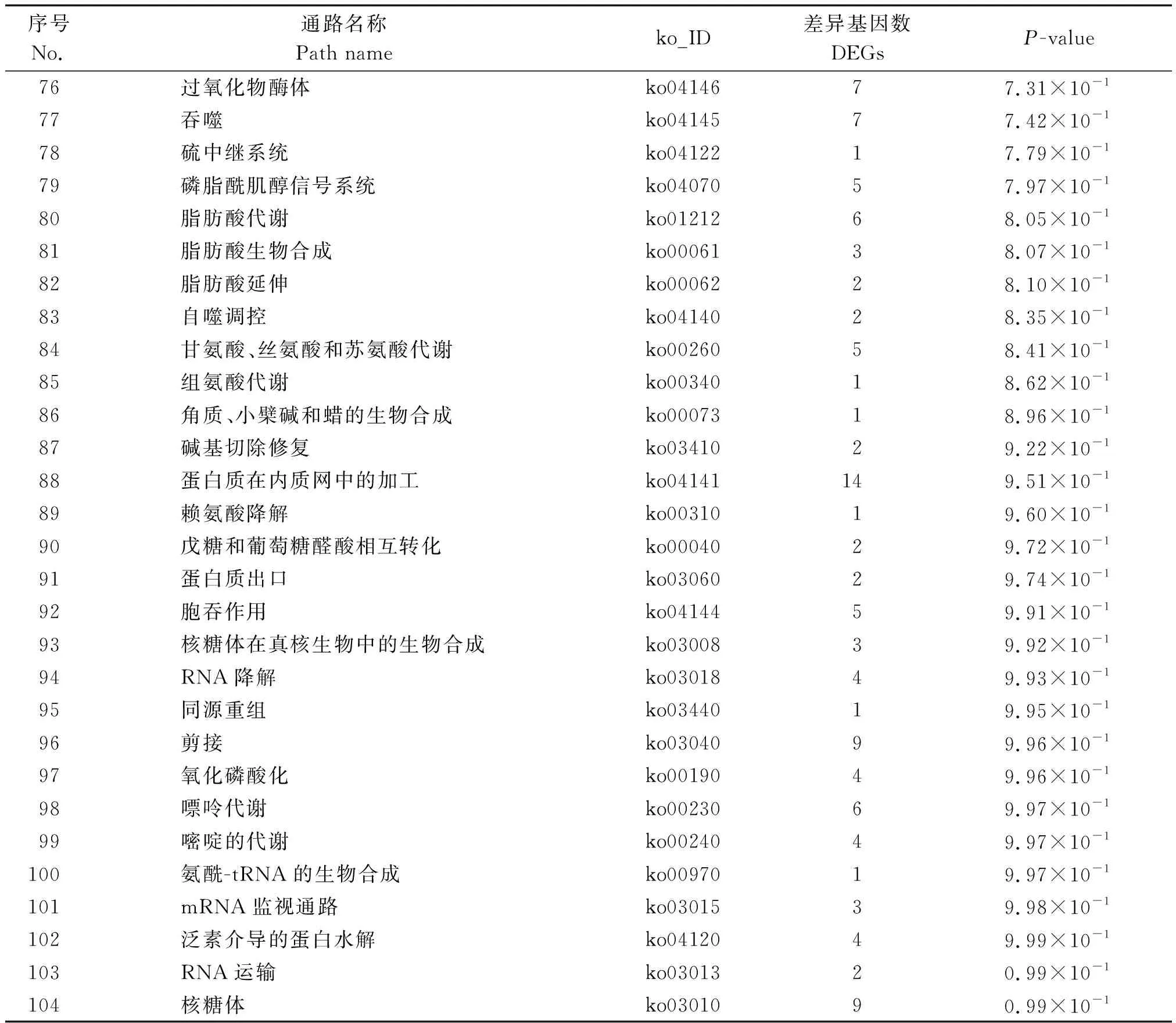
表2(续)
2.6 qRT-PCR验证基因表达
为了验证转录组分析结果中差异基因表达的可靠性,上调基因中选取了一个与植物病原体相互作用的基因OS05G0502000、一个与氮代谢相关的基因OS02G0770800、2个参与苯丙氨酸代谢的基因OS01G0869800、OS07G0677200;下调基因中挑选了4个与光合作用有关的基因OS07G0558400、OS01G0501800、OS03G0592500、OS02G0581100,利用qRT-PCR方法进行验证(表3)。使用RNA-Seq和qRT-PCR比较基因表达水平,以对照组中各基因表达量为1,统计低温处理下各基因的相对表达量。变化结果表明,qRT-PCR验证的基因表达量变化趋势和RNA-Seq差异基因表达趋势保持一致,如图7所示。表明转录组分析所得的数据具有较高的可靠性。

表3 实时定量PCR引物Tab.3 Primers used for qRT-PCR analysis
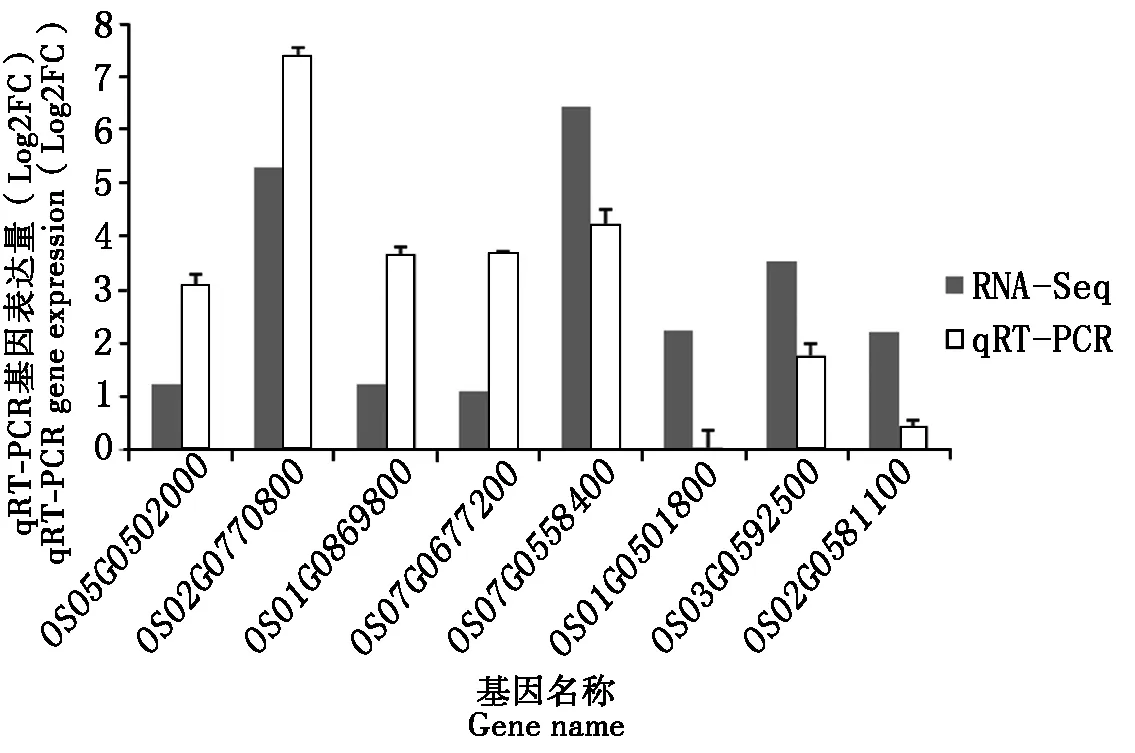
图7 RNA-Seq和qRT-PCR基因表达水平比较Fig.7 Comparison of RNA-Seq and qRT-PCR gene expression levels
2.7 新基因功能预测
综合各种外显子预测的算法和人们对基因结
构信息的认识,可预测出可能的完整基因[25]。新基因的预测有助于分析的完整性。转录组数据分析发现了25个新基因,它们的长度在779~6 551 bp不等,主要出现在1,2,3,4,5,6,7,8,10,12号染色体上,有6个基因没有具体的信息描述,其他的主要表现在线粒体前体、钙转运ATP酶、氯通道蛋白和转绿因子WRKY等方面(表4)。
水稻的WRKY、MYB、bHLH、bZIP、C2H2等家族调控水稻的生长发育和耐逆抗病过程[26-28]。本试验发现,2个WRKY转录因子,即Novel00699和Novel00786。这2个WRKY转录因子可能参与了应答低温环境的信号刺激,被诱导表达,提高作物抗逆能力。
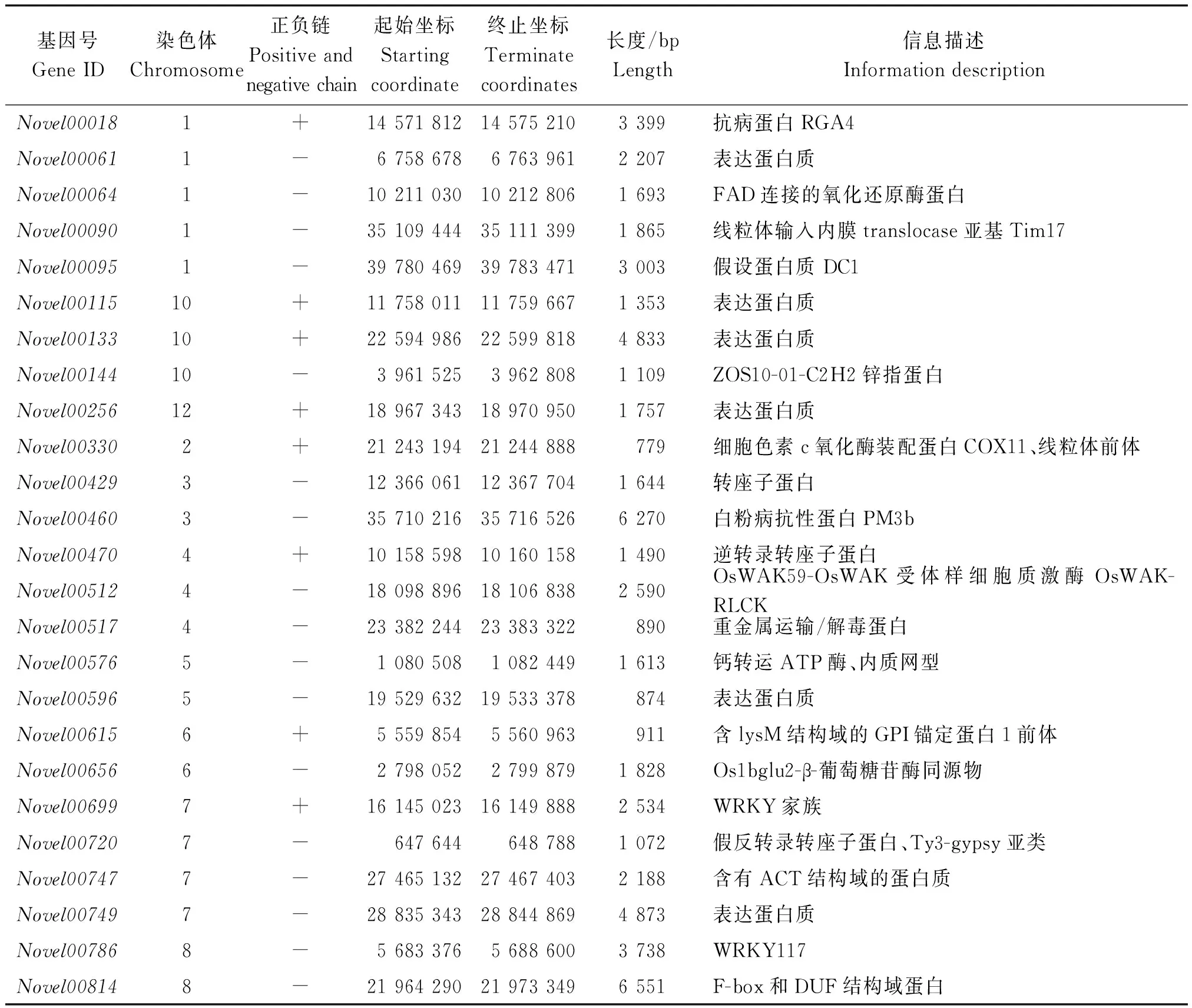
表4 新基因功能预测Tab.4 Novel gene function prediction
2.8 预测的互作蛋白
本研究分析得到5个与光合作用有关的基因,分别为LOC_Os01g31690、LOC_Os03g56670、LOC_Os04g33830、LOC_Os07g04840、LOC_Os08g44680,预测有16个基因与其互作;一个与PAL有关的基因LOC_Os02g41630,且预测到有3个基因与其互作(表5)。这些结果有助于对水稻幼苗响应低温胁迫的调节机制的进一步研究。
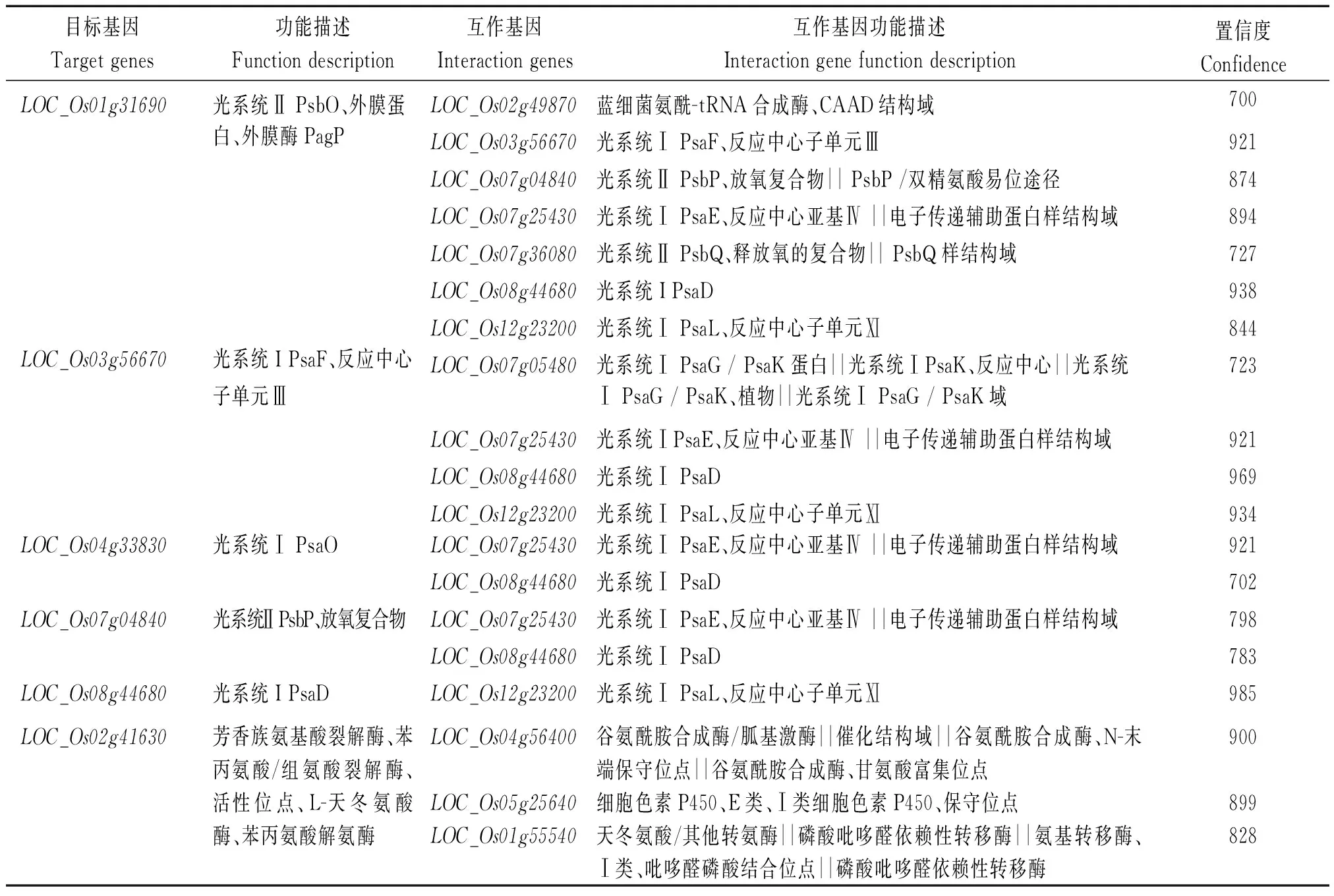
表5 预测的互作蛋白Tab.5 Predicted interaction proteins
3 结论与讨论
3.1 结论
本研究对正常条件和低温胁迫下水稻苗期叶片进行转录组测序。通过GO功能注释、KEGG Pathway、聚类分析等方法在正常条件和低温胁迫下进行差异基因表达水平上的比较,筛选出2 044个DEGs,其中,1 252个基因表达上调,792个基因表达下调;发现低温胁迫后差异表达基因主要参与光合作用、苯丙氨酸代谢等途径;在低温胁迫下,水稻幼苗的光合作用受到影响,其中该作用中光系统Ⅱ与光合运输电子受到的影响较大,基因变化趋势主要体现为下调;并且低温诱导了PAL活性升高,促进苯丙烷类次生代谢产物的合成,增强水稻幼苗在低温胁迫下的耐受力。通过新基因预测发现,差异表达基因可能参与转录因子的调控。本研究提供了水稻苗期低温研究方面一定的表达数据,同时也有助于对水稻苗期低温响应的分子调控机理的进一步研究。
3.2 讨论
国内外学者针对低温胁迫对植物光合作用的影响有着大量研究。王春萍等[29]研究了低温胁迫对水稻幼苗不同叶龄叶片叶绿素荧光特性,结果发现,低温胁迫对水稻幼苗三叶期的光合利用能力造成了一定影响。PSⅡ是植物光合作用过程中进行光反应的重要结构,低温胁迫会导致光能和光合系统Ⅱ(PSⅡ)电子传递转化效率降低,造成水稻的净光合速率、气孔导度与蒸腾速率均明显下降[30-32]。Kaniuga等[33-34]研究证明,低温条件下降低了叶绿体的希尔反应活性和类PSⅡ的电子传递活性。本研究发现,低温处理水稻幼苗后在光合作用通路上的差异表达基因,分别体现在光系统Ⅰ、光系统Ⅱ、细胞色素b6/f复合体、光合运输电子和F型ATP酶5个方面,受温度影响DEGs数量较多的为光系统Ⅱ和光合电子运输2个部分。光系统Ⅱ中的DEGs有21个表现下调,3个表现上调;光合电子运输中有20个差异表达基因,有8个表现上调,其余均为下调。通过这些数据可以补充说明:低温胁迫下,水稻幼苗的光合作用受到影响,且该作用中光系统Ⅱ与光合运输电子受到的影响较大,基因变化趋势主要体现下调。
PAL在植物生长发育和抵御环境胁迫等方面具有重要的生理意义[35-36]。黄小贞等[24]发现在不同的组织和时间段PAL基因的表达具有特异性,并且受内部发育信号等的调节。董春娟等[37]研究得出,黄瓜幼苗中的PAL基因表达和酶活性可在10 ℃低温条件下被诱导升高,证明了在低温胁迫下,黄瓜中的PAL基因参与黄瓜幼苗对低温胁迫的响应且存在基因功能的重叠。油菜在2 ℃处理后,叶片经PAL抑制剂氨基茚磷酸处理,叶片物质积累会发生迟缓,且Fv/Fm的值下降。证实PAL活性受到抑制会导致幼苗对低温胁迫的抗性降低[38]。本试验发现,水稻幼苗在低温胁迫下,有17个DEGs参与苯丙氨酸代谢通路,其中有3个注释到PAL功能,表达量均表现为上调。结合本试验结果说明,水稻幼苗为在低温条件下提高自身的抗逆性,保证其正常生长发育。所以,低温诱导了PAL的活性升高,促进苯丙烷类次生代谢产物的合成,缓解低温引起的不利因素。
众多研究表明,拟南芥与水稻 WRKY 基因家族的生物学功能和复杂相关特征已被广泛的挖掘并进行了功能鉴定[39-42]。目前,粳稻和籼稻分别鉴定出98,102个WRKY转录因子[41],生物和非生物逆境因子可大量诱导WRKY基因表达[43-45]。鄂志国等[46]将水稻WRKY家族按WRKY结构域基本结构特征的差异为3类,且得出它们通过调控生长调节物质所介导的信号途径,广泛的影响水稻的生长发育、生物与非生物胁迫应答。彭喜旭等[47]研究表明,WRKY80可能通过茉莉酸/乙烯介导的信号途径参与或调控防御反应。Shimono等[48-54]在水稻中找到WRKY45、WRKY13、WRKY03等多个WRKY家族基因被证明参与抗病防御调节。仇玉萍等[55]证实水稻中的7个WRKY因子(OsWRKY8、OsWRKY12、OsWRKY13、OsWRKY14、OsWRKY17、OsWRKY23 和OsWRKY45)能被4 ℃低温诱导表达;Zhou等[56]研究发现GmWRKY21过量表达可提高拟南芥抵抗低温的能力。本试验研究发现,对照组和处理组的水稻幼苗共有10个WRKY转录因子发生差异性表达,有8个DEGs表现上调,分别为WRKY21(OS01G0821600)、WRKY24(OS01G0826400)、WRKY121(OS03G0741400)、WRKY68(OS04G0605100)、WRKY7(OS05G0537100)、WRKY8(OS05G0583000)、WRKY69(OS08G0386200)、WRKY97(OS12G0116400);2个DEGs表现下调,分别是WRKY30(OS08G0499300)、WRKY90(OS09G0481700)。新基因预测中也发现了2个WRKY转录因子WRKY115(Novel00699)和WRKY117(Novel00786),经过低温处理后,水稻幼苗中部分WRKY转录因子的表达量发生变化,说明一些WRKY转录因子参与低温胁迫。
猜你喜欢
军事文摘(2021年16期)2021-11-05
今日农业(2020年23期)2020-12-15
心电与循环(2020年1期)2020-02-27
劳动保护(2018年8期)2018-09-12
江苏农业科学(2017年5期)2017-04-15
山东工业技术(2016年15期)2016-12-01
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
湖北农业科学(2014年3期)2014-07-21
