
苎麻脱胶细菌菌株的筛选、分类鉴定与多样性分析
2018-11-01 06:03:00郑科段盛文成莉凤冯湘沅刘正初曾洁郜明强彭源德
中国麻业科学 2018年5期
郑科,段盛文,成莉凤,冯湘沅,刘正初,曾洁,郜明强,彭源德
(中国农业科学院麻类研究所,长沙410205)
苎麻(Boehmeria niveaL.)等麻类韧皮的纤维细胞被大量键合型非纤维素物质包被或镶嵌,很难采用简单的物理加工方法提取[1]。以大量酸、碱处理为技术核心的化学脱胶方法,存在对环境污染严重等弊端。现代生物脱胶技术是麻类加工技术的发展方向,主要分酶脱胶和菌脱胶两类[2]。生产试验中,酶制剂脱胶成本相对较高,微生物脱胶技术更加成熟和实用[3]。但生物脱胶技术在生产中大规模稳定应用依然少见,主要归因于缺少脱胶能力强、性状稳定的菌种,故脱胶微生物已成为麻类脱胶技术发展的关键[4]。国内外报道的脱胶微生物主要有Erwinia carotovora、Pec-tobacterium carotovorum、Bacilluspumilus、Bacillussubtilis、Aspergillusniger等细菌和真菌[5-6]。有研究人员通过基因克隆定位诱变[7]、构建工程菌株等现代生物技术手段来提高脱胶菌种选育的效率[8],或者以混合菌的方式进行脱胶研究[9],这两种途径均需要合适的脱胶菌株作为支撑。因此,在微生物脱胶研究中,发掘生长条件简单、生长周期短的可培养细菌更具实践意义。
本研究从不同生境中采集样品,以苎麻韧皮原料为基质,通过富集、分离与纯培养,进行分离筛选,获取具有脱胶功能的细菌资源,通过形态特征、生理生化试验和16S rDNA测序,对菌种进行初步的分类鉴定和多样性分析,并从中筛选出代表性的菌株进行产酶等功能分析,旨在为揭示不同脱胶菌的差异和关联提供参考,同时为脱胶微生物遗传改良、脱胶复合菌群的构建等生物脱胶技术提供菌种资源。
1 材料与方法
1.1 试验材料
苎麻原麻:产自湖南长沙,手工剥制的去壳干皮。
分离基质:在湖南长沙和海南三亚2个地区的麻类作物田地土壤、麻类植株腐殖质、喂养苎麻饲料的牛羊粪便、椰壳麻堆肥4类环境的不同部位取20个样品,作为细菌分离基质。
1.2 主要试剂及培养基
细菌DNA提取、PCR扩增等试剂产自TaKaRa公司。果胶(柑橘,BR)和木聚糖(桦木,GR)产自Sigma公司。其它常规试剂为国产(BR或AR)。
富集培养基:生苎麻30.0 g,牛肉膏1.5 g,NaCl 1.5 g,蛋白胨3.0 g,自然pH,蒸馏水定容至500 mL,121℃灭菌30 min。
初筛麻瓶、复筛麻瓶:225 mL灭菌水,接种后加入15.0 g灭菌原麻。
分离平板培养基:果胶 5.0 g/L,蛋白胨1.5 g/L,NaCl1.5 g/L,琼脂15.0 g/L,自然 pH,121℃灭菌30 min。
木聚糖酶活性检验培养基:NaCl 6.0 g/L,MgSO40.1 g/L,KH2PO40.5 g/L,CaCl20.1 g/L,(NH4)2SO42.0 g/L,K2HPO42.0 g/L,木聚糖 5.0 g/L,酵母粉 1.0 g/L,琼脂 15.0 g/L,pH 7.0。
刚果红染色液:用蒸馏水溶解刚果红,终浓度为0.1%(w/v)。脱色液:终浓度为1.0 mol/L的NaCl溶液。
1.3 试验方法
1.3.1 菌株的初筛、分离和纯化
富集培养:称取菌样基质约30 g,加入300 mL灭菌水中,120 r/min震荡培养10 min,双层纱布过滤,取滤液30 mL,加入富集培养基中,于33~35℃、80 r/min震荡发酵2 d。
初筛:取以上样品富集培养液10 mL分别加入初筛麻瓶中,33~35℃、80 r/min震荡发酵2 d,观察原料变化,能使原麻明显变软或纤维分散的发酵液中可认为有脱胶菌群。空白对照的麻瓶中,加入纯净水代替富集培养液。
分离与纯化:取富集培养液用果胶琼脂平板进行稀释涂皿,平板于34~36℃培养36 h,分离出单菌落;对分离的菌株进行纯化、菌落和菌体形态观察以及革兰氏染色试验。
1.3.2 16S rDNA分子鉴定和系统发育树构建
提取高质量的细菌基因组DNA,PCR扩增16S rDNA,由广州基迪奥公司采用通用引物27F、1492R进行16S rDNA测序;序列在GenBank进行BLAST比对分析,选择相似性最高的种属做为菌种鉴定初步结果;利用Clustal X软件进行多序列比对,用MEGA 7软件构建系统进化树,进行聚类分析。
1.3.3 多样性分析
对筛选的细菌进行种群分类、分离基质分类,并进行形态多样性以及功能多样性研究。采用多样性统计方法,对不同样品基质群落类型中的脱胶菌种进行物种多样性研究,包括多样性指数、优势度、均匀性以及相似性指数。
Shannon多样性指数H′=-∑(PilnPi)=-∑(Ni/N)ln(Ni/N),Ni是第i个物种的个体数,N是全部物种的总个体数;Simpson优势度指数C=∑(Pi)2=∑(Ni/N)2;Pielou均匀度指数E=H′/lnS,H′是多样性指数,S是物种的数目;群落相似性Jaccard指数Cs=c/(a+b-c),a为A群落物种数,b为B群落物种数,c为A、B两群落共有的物种数[10,11]。
1.3.4 脱胶功能复筛
对纯培养单菌进行脱胶能力的筛选:采用苎麻脱胶实效法,将纯培养菌液加入复筛麻瓶,33~35℃、120 r/min震荡发酵,并设空白对照,发酵36 h,以是否明显发生原麻变软或纤维分散判断是否脱胶。
1.3.5 木聚糖酶检验
采用透明圈法,测量计算水解透明圈直径与菌落直径的比值,对复筛产生的菌种进行产木聚糖酶的验证。以本团队选育的脱胶菌E.carotovora CXJZ95-198为对照(CK)。
2 结果与分析
2.1 菌株分离与筛选结果
经富集和初筛,20个样品全部具有一定的脱胶能力,形成20个脱胶复合菌群。因果胶降解能力是脱胶菌的重要特性,所以菌群通过在果胶琼脂平板(含少量氮源和无机盐)上生长分离,然后进一步筛选,根据基本形态去掉源自同一个样品的重复菌株,共获得78个苎麻脱胶菌种。
2.2 16S rDNA分子鉴定
通过16S rDNA分子鉴定,将筛选的78个菌种初步分为17个属,分别为无色杆菌属(Achromobacter)、不动杆菌属(Acinetobacter)、气单胞菌属(Aeromonas)、芽孢杆菌属(Bacillus)、短芽孢杆菌属(Brevibacillus)、金黄杆菌属(Chryseobacterium)、柠檬酸杆菌属(Citrobacter)、肠杆菌属(Enterobacter)、微小杆菌属(Exiguobacterium)、赖氨酸芽孢杆菌(Lysinibacillus)、泛菌属(Pantoea)、假单胞菌属(Pseudomonas)、嗜冷芽孢杆菌属(Psychrobacillus)、鞘氨醇杆菌属(Sphingobacterium)、葡萄球菌属(Staphylococcus)、土地芽孢杆菌属(Terribacillus)、魏斯氏菌属(Weissella),至少 27种(表 1)。其中,芽孢杆菌类占70.51%,这与其他生物脱胶类报道中芽孢杆菌比较多见的情况一致[5-9]。4类基质中,麻类腐质中筛选出的菌种数最多,占43.59%。
将序列与NCBI数据库比对后发现,除2个(赖氨酸芽胞杆菌和鞘氨醇杆菌)同源性为95%和97%以外,其它均在99%以上,种属鉴定的可信度较高。对应参考菌株序列构建系统发育树,从图1可以看出,筛选的脱胶可培养细菌主要聚为8大类,其中7大类基本为芽孢杆菌类细菌,属革兰氏阳性菌,另外聚成一大类的基本为革兰氏阴性菌。
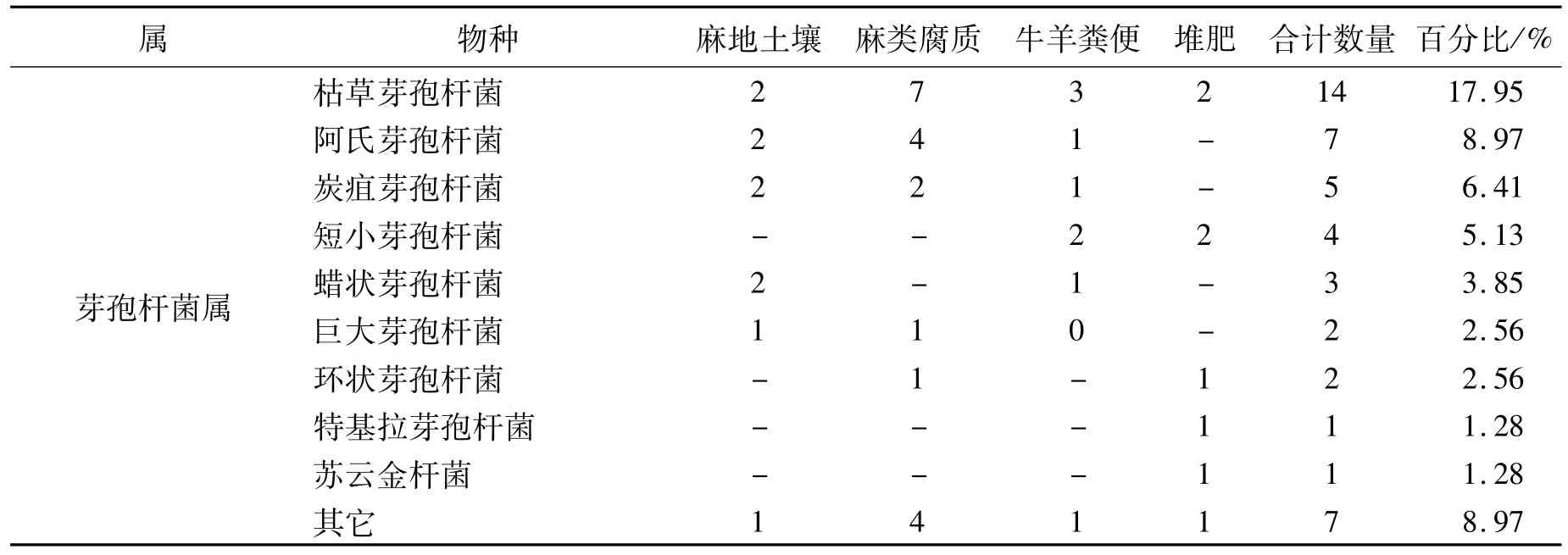
表1 脱胶菌种与其对应的分离基质Table 1 The degumming bacteria and their habitats
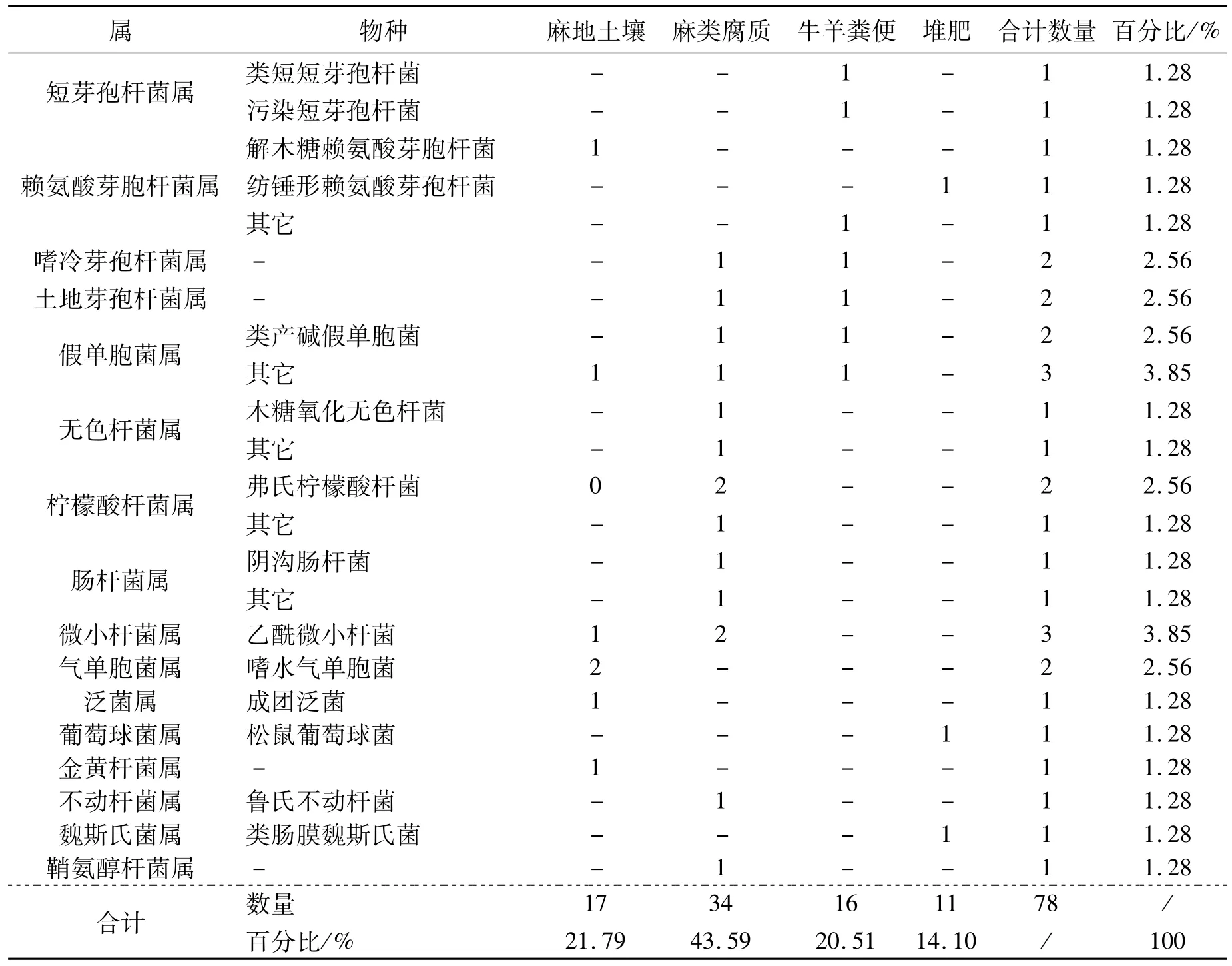
续表1
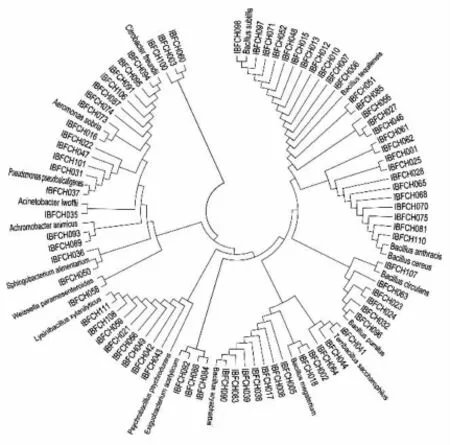
图1 脱胶细菌系统发育示意图Fig.1 Cluster analysis for degumming bacteria based on 16S rDNA sequencing
2.3 多样性分析
对于来源于不同基质的菌种,其多样性指数如表2所示。多样性指数按麻类腐质、牛羊粪便、麻园土、堆肥依次下降。麻类腐殖质中的脱胶菌含量丰富,是分离专用菌种的较好基质。堆肥中的脱胶细菌丰富程度相对较低,但是堆肥优势度相对明显,这可能是因为堆肥中有大量椰壳发酵物,成分和发酵条件较为特殊,其菌株已经经历了发酵过程的自然筛选。均匀性都较好,这与初筛、分离时去掉同一样品形态重复菌株有关。

表2 不同分离基质的多样性指数Table 2 Diversity indexes of different habitats
表3所示来源不同基质菌种的相似性指数,所有样品来自于2个地理环境差异很大的地区,而且同一个地区的每个取样生境差异大,所以4类样品间整体相似度不高。堆肥和其它样品都极不相似,说明其提供给微生物的营养和温度等条件专一性比较强。这与麻类脱胶微生物资源在厩肥中的多样性研究结果相似[11]。

表3 不同分离基质的相似性指数Table 3 Similarity indexes of different habitats
2.4 脱胶功能复筛
通过对78个菌种进行苎麻脱胶功能复筛,有10个菌种分别在24 h内能够使苎麻明显脱胶。编号为:H006、H021、H028、H032、H038、H046、H050、H056、H060、H096。

图2 有明显脱胶效果的苎麻试样Fig.2 Ramie samples obviously degummed
2.5 木聚糖酶活力验证
对以上10株有显著脱胶功能的菌进行水解透明圈试验,结果表明,大部分菌株具有分泌木聚糖酶的能力,其中H021和H056与对照菌株相比,分解木聚糖的能力显著。10株菌株的木聚糖酶水解圈结果如图3和表4所示。

图3 分泌木聚糖酶形成的水解圈比较Fig.3 Comparison of hydrolyzed circle caused by bacteria.
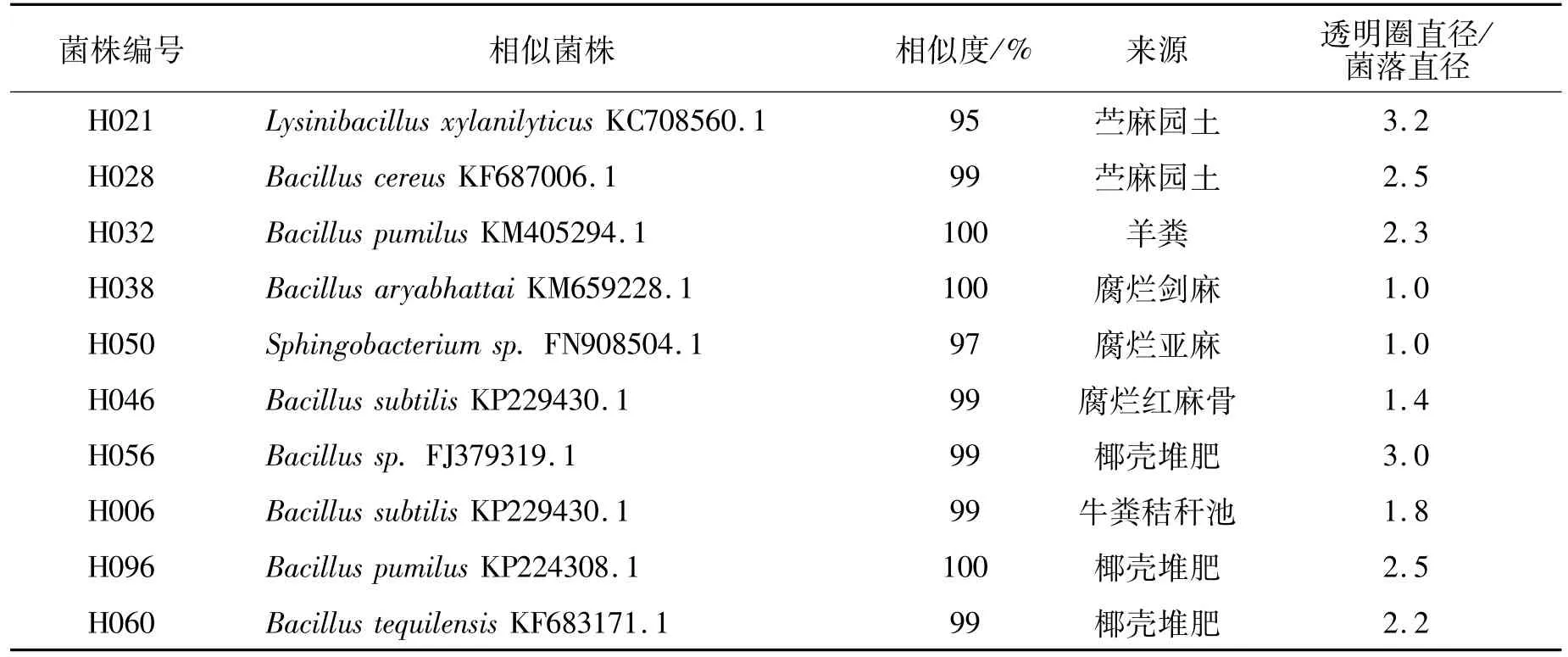
表4 复筛出的10个菌种的信息Table 4 Information of the 10 strains further studied
3 讨论
本研究筛选、分离出数量较多的具有一定脱胶功能的细菌。以苎麻韧皮原料为培养基具有良好的富集和筛选效果,而采用果胶琼脂平板分离培养可进一步提高筛选的准确性。本研究筛选出的菌种具有降解苎麻和果胶的能力,在此基础上,采用简易的脱胶实效法和木聚糖透明圈法对细菌的脱胶功能进行了初步鉴定。但微生物脱胶酶系复杂,有脱胶能力的微生物究竟是哪些酶在起作用仍不完全清楚。本研究中对脱胶效果的判断并不全面,所筛选的脱胶菌株可能产生纤维素酶,进而对纤维产品质量带来不利影响。因此,这些细菌的产酶特性还有待进一步研究。
物种多样性分析表明,通过麻类作物种植土壤、麻类腐殖质、牛羊粪便和堆肥等麻类植株所处的环境和植物降解环境里的基质能分离出大量功能菌种。麻类腐殖质所含功能菌种丰富,厩肥等只能适合于特定的类群生长,种类较少。本研究只采用了单一培养基进行分离,培养条件也限定在比较狭窄的范围,对微生物多样性反映有一定局限。微生物资源多样性研究需要足够大的样本量,如果取样和分离的菌种数量少,或去掉同一样品中的重复菌株,则影响多样性分析结果的准确性。另外,富集后的菌群表现出脱胶能力,这其中可能有不可培养微生物的作用。
本研究筛选出的赖氨酸芽胞杆菌(Lysinibacillus sp.)和鞘氨醇杆菌(Sphingobacterium sp.),此前报道中极少见到应用于麻类脱胶,且比对的相似性较低,具备潜在新种的可能,值得深入研究。
虽然16S rDNA分子鉴定效率高,但少数菌种未能鉴定到种。且聚类分析中发现,有些分子鉴定结果表明应该聚为一组的菌却单独相聚(如H003、H060、H100);少数菌种比对的相似性不高。因此,部分菌种的分子鉴定和系统发育树的构建需要完善和改进,要更精确地对筛选出的菌种进行鉴定,还需要结合外观形态、生理生化特征进行鉴定。
猜你喜欢
中国麻业科学(2021年5期)2021-12-02 02:08:44
粮食与食品工业(2021年1期)2021-02-06 12:18:20
中国麻业科学(2018年6期)2018-04-09 11:22:38
质量技术监督研究(2018年1期)2018-03-26 08:04:36
中国麻业科学(2018年1期)2018-03-15 01:03:48
中国麻业科学(2017年6期)2017-12-21 08:17:37
动物营养学报(2017年2期)2017-02-28 05:04:26
中国麻业科学(2015年5期)2015-12-28 06:22:11
中国麻业科学(2015年5期)2015-12-28 06:22:08
中国麻业科学(2015年5期)2015-12-28 06:22:07
