
硫化氢在缓解拟南芥镉胁迫中的作用
2018-10-17 06:08贾红磊刘珂娜
陕西科技大学学报 2018年5期
贾红磊,刘珂娜,刘 洋,杨 珺
(陕西科技大学 环境科学与工程学院,陕西 西安 710021)
0 引言
镉是植物生长的非必需因素,是目前广泛存在的重金属污染源之一.近年来,随着资源的开发与利用,造成土壤中重金属含量升高,其中镉成为我国农田土壤重金属污染的主要元素之一[1].由于镉存在着强毒性与可迁移性,当在环境中积累到一定水平时容易被植物的根系吸收,并可被运输到地上部分,通过食物链进入人体,对身体健康造成潜在的威胁.过量的镉会通过损伤植物细胞的结构、抑制细胞分裂、破坏酶活等方式直接或间接抑制植物的生理过程,最终抑制植物生长甚至导致植物死亡[2].作物受重金属胁迫时主要表现为株高和根都会比正常植株矮、细黄、结实率下降.近年来,如何解除或缓解重金属胁迫成为国内外研究的热点科学问题.
硫化氢(H2S)是一种无色有臭鸡蛋气味的气体,并且也是一种重要的气体信号分子.目前,气体信号分子有NO、CO和H2S三种,在动物体内H2S的产生机制和生理作用已经被充分证明,目前关于植物体内H2S 的合成及功能研究也取得了较大进展[3].H2S参与了许多植物的生长发育过程,如调节气孔运动、促进根的发育和增强光合作用、缓解干旱、渗透和金属离子等多种非生物胁迫造成的伤害等[4].H2S通过调控基因表达、离子通道活性等参与植物生长发育和逆境胁迫响应等过程,其与活性氧(ROS)、NO、CO、Ca2+,以及脱落酸(ABA)、茉莉酸(Ja)、赤霉素(Ga)、生长素、乙烯等植物激素信号路径存在相互作用.最近研究表明,外源生理浓度 H2S可通过上调病程相关基因的表达和一些次级代谢产物的含量,增强植物抵抗生物胁迫能力[5].
综上所述,H2S信号可从不同水平增强植物应答各种非生物胁迫和生物胁迫的能力,并参与调节植物的整个生长、发育、成熟和衰老过程.Jin等[6]证明硫化氢通过与脱落酸(ABA)的互作影响气孔的关闭,进而提高拟南芥的抗旱性.另外,NaSH处理也诱导了干旱相关基因的表达,表明H2S还参与了拟南芥对干旱胁迫的响应.目前关于H2S对拟南芥抗重金属胁迫的研究相对较少.
本研究以模式植物拟南芥为材料,用NaSH作为供体提供H2S,采用培养基培养拟南芥分析镉胁迫的条件下外源H2S信号对植物生理生化指标的影响,探究H2S在提高拟南芥对镉抗性中的作用,为镉污染的防治提供理论依据.
1 实验部分
1.1 材料及培养
拟南芥种子野生型购自于ABRC公司,将春化后的拟南芥种子经10%的次氯酸钠溶液消毒10 min后,用无菌水冲洗5~6次播种于含1%蔗糖和0.8%琼脂的1/2 MS培养基(pH 5.8)上,播种好的培养皿垂直放置于人工气候箱中,使种子呈水平排列,进行垂直培养.培养条件为恒温22 ℃,相对湿度(H):75%,16 h光照/8 h黑暗周期[7].
1.2 实验方法
1.2.1 种子萌发测定
将拟南芥种子分别播种于空白培养基、含有50μmol/L CdCl2、50μmol/L NaSH(用以提供H2S)、50μmol/L CdCl2+50μmol/L NaSH的培养基上进行培养,培养条件如1.1.播种后按式(1)计算不同处理培养基中48 h、72 h、96 h种子的萌发率.
萌发率=发芽种子数/受试种子数×100%
(1)
1.2.2 根长测定
拟南芥种子在基础1/2 MS培养基上生长5 d后,将幼苗转移至含有不同处理条件的培养基中继续垂直培养3 d,对幼苗进行成像,并用Image-J测量根长度[8].
1.2.3 离子渗透率的测定
将7日龄拟南芥幼苗在含有不同化学物质的1/2 MS琼脂培养基上处理5 d.处理后,收集拟南芥根并用去离子水中洗涤3次以上,去除表面附着的电解质.然后,将待测样品放置于含有10 mL去离子水的试管中,25 ℃条件温育3 h.温育后,测定浴液中的电导率C1,并测定去离子水的电导率C0.之后将样品在沸水中加热1 h,然后在测量浴液中总电导率C2,按式(2)计算离子渗透率.
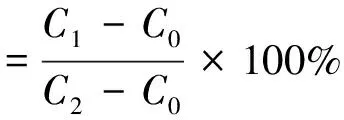
(2)
1.2.4 叶绿素含量测定
叶绿素用体积分数为80%的丙酮提取,之后用分光光度法测定,计算叶绿素a、叶绿素b及总叶绿素的含量.
1.2.5 MDA、SOD含量测定
MDA含量的测定采用硫代巴比妥酸法[9],SOD活性的测定采用氯化硝基四氮唑蓝法(NTB)[9].
1.3 数据分析
本实验数据统计分析使用Orign7.0、Image J及SPSS7.0软件进行统计分析和显著性分析.所有数据均为三次试验平均值.
2 结果与讨论
2.1 镉胁迫下H2S对拟南芥生长的影响
2.1.1 对拟南芥萌发率的影响
种子萌发是系统认识重金属伤害机理的较好途径,有助于了解镉胁迫下H2S拟南芥的影响.最近的研究发现,适宜浓度的外源H2S(NaSH)预处理可提高大豆种子的发芽率[10]. 本实验条件下种子的发芽率如图1所示.结果表明,在50μmol/L的CdCl2浓度下及加入外源H2S后,拟南芥种子的发芽率与对照组相比没有显著性差异,说明50μmol/L的CdCl2并不影响拟南芥种子萌发.
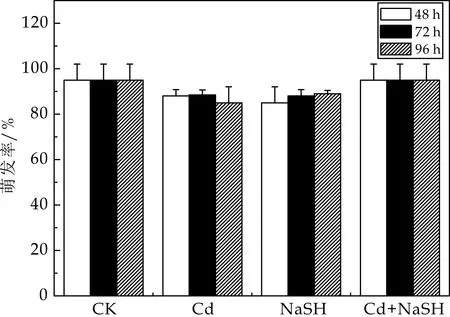
图1 不同处理对拟南芥萌发率的影响
2.1.2 对根长的影响
重金属影响植物的作用机理主要包括抑制植物根尖细胞的分裂或根的伸长、影响植物激素的分布或细胞死亡等[11].高浓度的镉严重影响植物幼苗根的生长[2].H2S作为一种气体信号分子还可能参与根发育的调控,一些研究结果显示H2S可能通过IAA和NO来诱导植物不定根的发生[12].如图2所示,用50μmol/L的Cd2+处理后,其根长明显较对照组的幼苗短,与对照组相比主根长度降低54.37 %,说明50μmol/L浓度的Cd2+能够显著抑制拟南芥根伸长,对拟南芥的根部生长造成了损伤.同时,与镉胁迫处理相比,镉胁迫下外源H2S(NaSH+Cd2+)处理下根长明增加,上调比例为48.94 %,说明外源添加H2S后,能够在一定程度上缓解Cd对拟南芥根生长的抑制作用,缓解Cd的胁迫作用,从而有助于提高拟南芥的抗逆性.
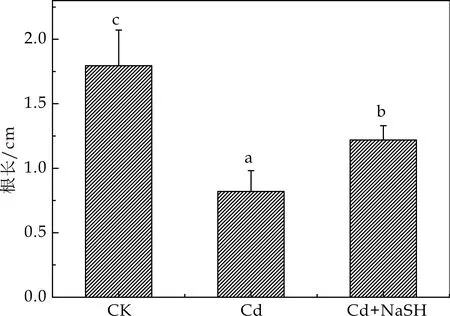
图2 不同处理对拟南芥根长的影响
2.2 镉胁迫下H2S对拟南芥生理指标的影响
2.2.1 离子渗透率的影响
当植物处于逆境环境时,膜结构被破坏导致细胞膜通透性增加,使细胞电解质外渗.如图3所示,与对照组相比50μmol/L CdCl2处理条件下,拟南芥的离子渗透率上调48.79 %,表明50μmol/L的CdCl2对拟南芥膜通透性造成损伤.此外再加入外源添加NaSH后,离子渗透率下降26.77 %,说明外源H2S在一定程度上能缓解Cd对植物细胞膜造成的损害.

图3 不同处理对拟南芥离子渗透率的影响
2.2.2 对叶绿素含量的影响
重金属能通过破坏光合过程中电子传递和叶绿素完整性影响植物光合作用,许多植物在重金属胁迫下都出现了叶片失绿现象[13].有研究表明拟南芥在镉处理下,叶绿素a、叶绿素b、胡萝卜素含量减少,说明镉对拟南芥的叶绿体造成了损伤[14].图4显示,与对照组相比在50μmol/L的Cd2+胁迫下,拟南芥叶绿素含量下降33.45 %.同时,在添加外源NaSH后与镉胁迫处理相比叶绿素含量上升45.83 %.说明在外源H2S供体NaSH的作用下,Cd对拟南芥的叶绿素的破坏作用得到了减缓,由此可见外源H2S是能够缓解镉胁迫对拟南芥叶绿素含量造成的影响.
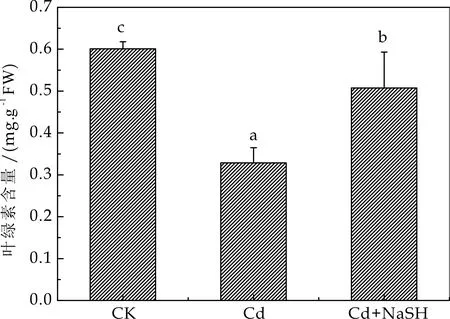
图4 不同处理对拟南芥叶绿素含量的影响
2.2.3 MDA含量
MDA是镉胁迫下植物体内活性氧类物质大量积累所造成的膜脂过氧化的产物之一.正常条件下,植物体内活性氧的产生和清除处于动态平衡.在镉胁迫下平衡被破坏导致活性氧大量积累,从而激活植物体内的抗氧化系统,诱导抗氧化酶活性发生变化[15].MDA的含量可反应膜脂过氧化的程度,间接表明膜系统受损程度及植物的抗逆性.图5表明,与对照组相比50μmol/L的CdCl2处理的拟南芥幼苗MDA含量显著上升,升高154.16%,说明50μmol/L浓度的CdCl2使拟南芥体内积累的活性氧自由基导致了膜脂过氧化.同时,在加入外源H2S后,与镉胁迫下相比MDA含量明显下降63.93 %.可见,外源H2S可缓解镉胁迫下拟南芥的膜脂过氧化程度.
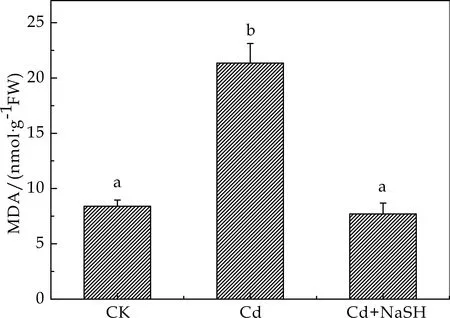
图5 不同处理对拟南芥MDA含量的影响
2.2.4 SOD含量的测定
SOD是一种广泛存在于动植物中的金属酶,可以催化氧自由基的歧化反应,生成过氧化氢,过氧化氢又可以进一步被过氧化氢酶转化为水和分子氧,该酶和植物的抗逆性及衰老有关,是清除植物体内活性氧的第一道防线.在镉浓度相对较低的情况下,植物通过提高抗氧化酶的活性来降低体内的过氧化自由基;但是在镉浓度过高的情况下则会与抗氧化酶的巯基结合,导致酶的催化中心或酶结构受损,造成其活性的降低[16].图6显示,在50μmol/L的镉胁迫下,与对照组相比,拟南芥的SOD活性下降16.70%.外源H2S处理提高了镉胁迫下SOD活性,与受50μmol/L CdCl2处理相比,SOD活性升高13.90 %.此结果表明外源H2S能够提高SOD的活力,降低拟南芥在镉胁迫下·O2-的含量,提高了拟南芥的抗逆性.
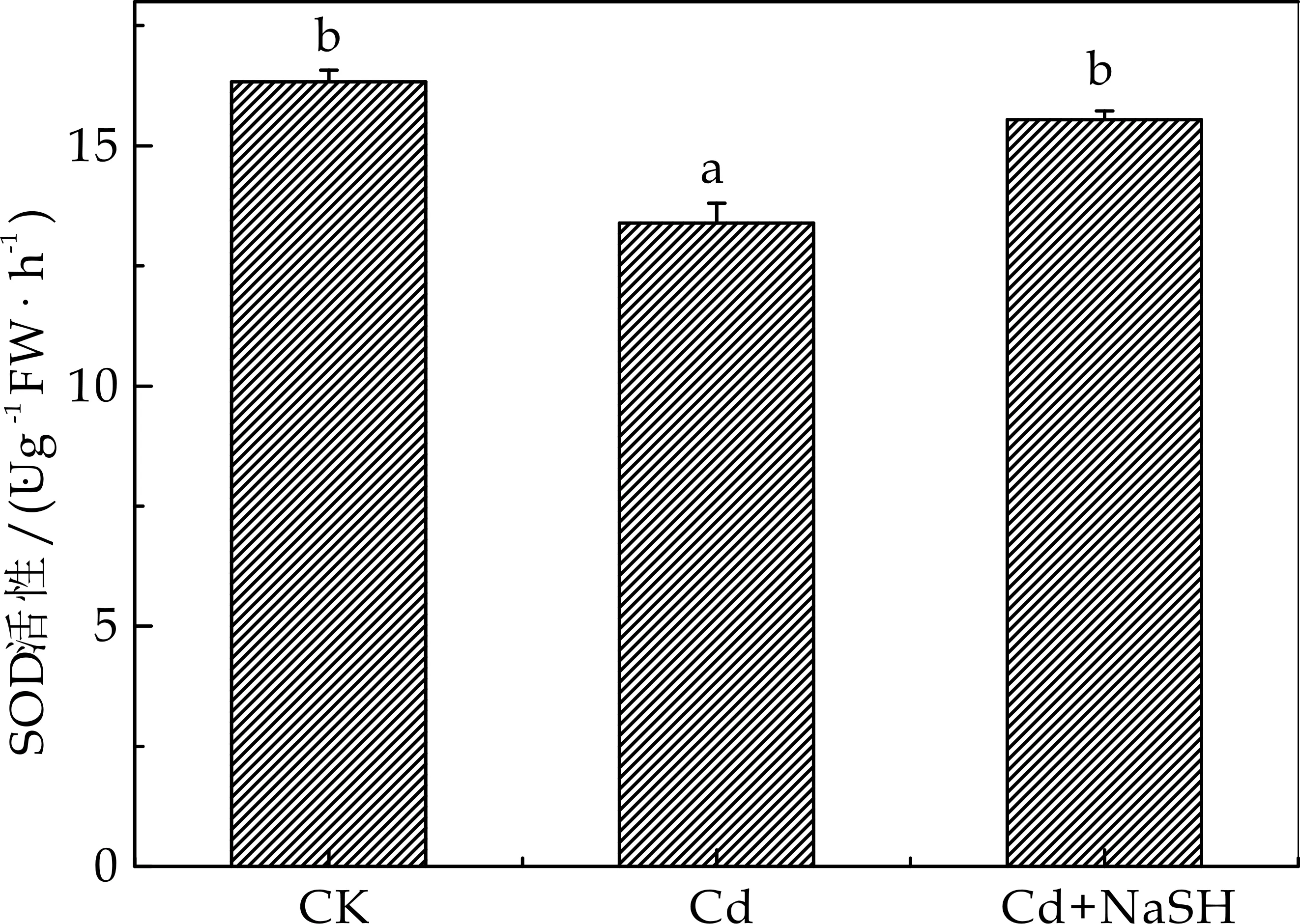
图6 不同处理对拟南芥SOD值的影响
3 结论
通过本研究发现,50μmol/L镉胁迫会抑制拟南芥根生长,降低叶绿素含量,增加MDA导致细胞内的氧化损伤,引起细胞结构与功能的破坏,降低SOD含量,提高种子中·O2-的含量,进而影响植物的正常生长;H2S供体NaSH处理能够通过诱导镉胁迫下提升SOD的活力,降低·O2-含量,说明H2S可以提高Cd2+胁迫下拟南芥的抗逆性,促进拟南芥幼苗的生长;H2S供体NaSH处理一定程度上延缓MDA含量的上升,减缓膜脂过氧化,保护膜结构的稳定性,为细胞的正常代谢提供了良好环境.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
海洋通报(2021年5期)2021-12-21
山西师范大学学报(自然科学版)(2021年3期)2021-11-02
农业资源与环境学报(2021年5期)2021-10-06
园艺与种苗(2020年12期)2021-01-08
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
现代园艺(2018年3期)2018-02-10
红领巾·探索(2015年9期)2015-09-10
