
桥山林区不同坡向麻栎林种群生态位特征分析
2018-10-12 06:33徐凤玲刘小军马建全张文辉
西北林学院学报 2018年5期
徐凤玲,刘小军,马建全,张文辉*
生态位可以反映出物种与环境之间的相互关系[1],是生态学重要的理论基础。生态位理论被广泛用于分析物种对环境的耐受能力和对资源的利用能力,以及种群所处群落的稳定性[2-3],成为研究种间关系、群落演替和生物多样性的重要手段[4]。生态位分析通过计算林地群落物种间重要值、生态位宽度、生态位重叠阐明种间关系,为森林经营抚育间伐提供依据。近年来,有关桥山林区麻栎林研究,主要集中在麻栎林的抚育间伐、营养成分、种群空间格局以及碳储量等方面[5-8],有关麻栎林生态位分析鲜有报道。杨澄[9]研究桥山天然栎林的生态位,并发现种间存在竞争,但具体哪些物种间生态位重叠,重叠有多大,对建群种麻栎和群落稳定性有什么影响并未阐明,而这是实现群落内种间关系和谐与群落持续正向演替的关键。
桥山林区地处黄土高原子午岭南部,植物种类丰富,森林类型多样。麻栎林占当地森林组成50%以上,属地带性森林植被,对黄土高原水源涵养具有重要作用[10]。然而随着人类对资源及土地利用程度的深化,桥山林区的麻栎林受到了严重的破坏,导致林分结构不合理,生产力低下,面积日益萎缩,亟待研究与改善。
本研究通过典型样地调查的方法,分析桥山林区不同生境下麻栎林群落内所有物种的生态位宽度和不同物种间生态位重叠状况,阐明不同生境对麻栎种群生态位影响、麻栎种群对环境资源的利用能力以及麻栎群落的正向演替驱动力,为桥山林区天然麻栎林的保护和可持续经营提供依据。
1 研究区概况
研究区位于黄土高原陕西省延安市桥山林区,地理位置 108°31'-109°12'E,35°29'-35°50'N,海拔945~1 165 m,处于暖温带半湿润与半干旱的气候过渡地带,年均温9.2℃,最高气温36℃,最低气温-23℃,具有大陆性季风气候特征,年均降水量612.3 mm(集中在7-9月),无霜期185 d,土壤类型为森林褐色土,地带性植被为落叶阔叶林。主要乔木种类:麻栎(Quercus acutissima)、辽东栎(Q.wutaishanica)、杜梨(Pyrus betulifolia)、茶条槭(Acer ginnala)、油松(Pinus tabuliformis);灌木种类:狼牙刺 (Sophora davidii)、卫矛(Euonymus alatus)、南蛇藤(Celastrus orbiculatus)、胡枝子(Lespedeza bicolor)等;草本种类:苔草(Carex lanceolata)、披碱草(Elymus dahuricus)、银兰(Cephalanthera erecta)。

表1 样地自然概况Table 1 Natural condition of the sample plots
2 研究方法
2.1 样地设置与调查
黄土高原区由于水分条件的限制,阴坡的林木通直高大,郁闭度高,耐阴性植物较多,而阳坡的林木质量相对较差,郁闭度较低,耐阴性植物种类较少。本研究对延安市桥山林区天然麻栎林充分踏查后,选择生境条件基本一致、受外界干扰较小的阴坡和阳坡,各设置12块面积为20 m×20 m的标准样地(表1),采用对角线法在每个标准样地内设置5个面积为5 m×5 m的灌木样方和5个面积为1 m×1 m的草本样方。对样地内乔木进行每木检尺,调查种类、坐标、高度、胸径、冠幅、频度;灌木和草本样方调查植物种类、数量、高度、基径、冠幅、频度和盖度。
2.2 数据处理方式
2.2.1 重要值计算公式

其中,相对胸高断面积为样地内某一乔木的胸高断面积之和占所有乔木的胸高断面积之和的百分比[11-12]。

2.2.2 生态位宽度计算公式
Levins生态位宽度(LB)
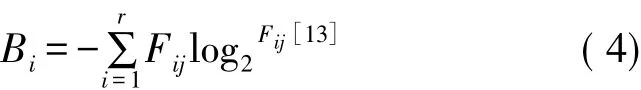
式中,Bi是种i的生态位宽度;Fij是种i对第j个样地的利用占其对全部资源利用的频度。即:

式中,nij为种i在样地j的优势度(本研究为物种重要值);Ni为种i所利用全部资源总优势度(即种i在所有样地的重要值之和);r为资源等级数(样地数);生态位宽度值域[0,logr]。
Hurlbert生态位宽度(HB);
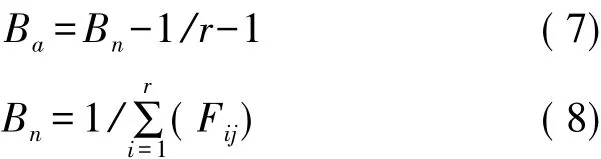
式中,Ba为生态位宽度,该方程的值域为[0,1][1]。
2.2.3 生态位重叠

式中,NO为生态位重叠值;nij和nkj为种i和k在样地j上的优势度[14],本研究为物种重要值。
3 结果与分析
3.1 不同坡向对桥山重要植物重要值与生态位宽度的影响
根据麻栎林主要植物的重要值之和计算可知(表2、表3),乔木层阳坡和阴坡麻栎种群重要值之和分别占所有物种重要值之和的67.57%、76.45%;阳坡灌木层狼牙刺、卫矛、胡枝子分别占所有物种重要值之和的 24.68%、23.90%、21.86%,阴坡狼牙刺、南蛇藤分别占26.62%、19.03%;草本层阳坡和阴坡苔草种群重要值之和分别占所有物种重要值之和的64.43%和53.80%;而其他的种群在各自资源位上占较小比例且相差不大。
应用LB指数和HB指数计算桥山林区天然麻栎林群落主要物种生态位宽度的趋势大致相同。阳坡麻栎林乔木层主要物种的生态位宽度排列顺序为麻栎、杜梨、茶条槭、辽东栎、山楂、槲栎、红麸杨、毛梾、山杏,阴坡的生态位宽度排列顺序为麻栎、茶条槭、辽东栎、槲栎、杜梨、红麸杨、梾木、白桦。麻栎在阴坡和阳坡的资源位都出现,在林内分布范围广,对环境资源的利用较充分,为该林分的建群种,在群落中的地位较高,其位于阴坡和阳坡的生态位宽度为最大值。而茶条槭、辽东栎为乔木层的主要伴生种,它们对环境的适应能力、数量与分布频度仅次于麻栎,所以它们的生态位低于麻栎。阳坡的山杏和阴坡的白桦虽在资源位中出现但频率低,对环境要求较苛刻,对环境资源的利用较弱,导致其生态位宽度较低。
狼牙刺、南蛇藤、胡枝子都属于灌木层的优势种。狼牙刺在阴坡和阳坡都有分布,生态适应范围大,在群落中所处地位高,生态位宽度最大。生长在阳坡的卫矛和南蛇藤生态位宽度仅次于狼牙刺,其对环境资源的利用能力和出现频率相对小,悬钩子则为阳坡生态位宽度最小的物种;在阴坡,南蛇藤和胡枝子的生态位宽度仅次于狼牙刺,生态位宽度最小的是菝葜。苔草在阳坡和阳坡所处草本群落中都占绝对优势,数量大且生态位宽度最大,阳坡中银兰生态位宽度最小,阴坡中则是白茅草。
在阴坡和阳坡,乔木层的麻栎、灌木层的狼牙刺、草本层的苔草生态位宽度都最大,坡向并未影响它们的生态位宽度,它们对桥山林区的环境适应、资源利用能力较强,分布广,生态幅大;而乔木层的杜梨、灌木层的卫矛、南蛇藤、胡枝子 、鼠李则受到坡向对其生态位宽度的影响。
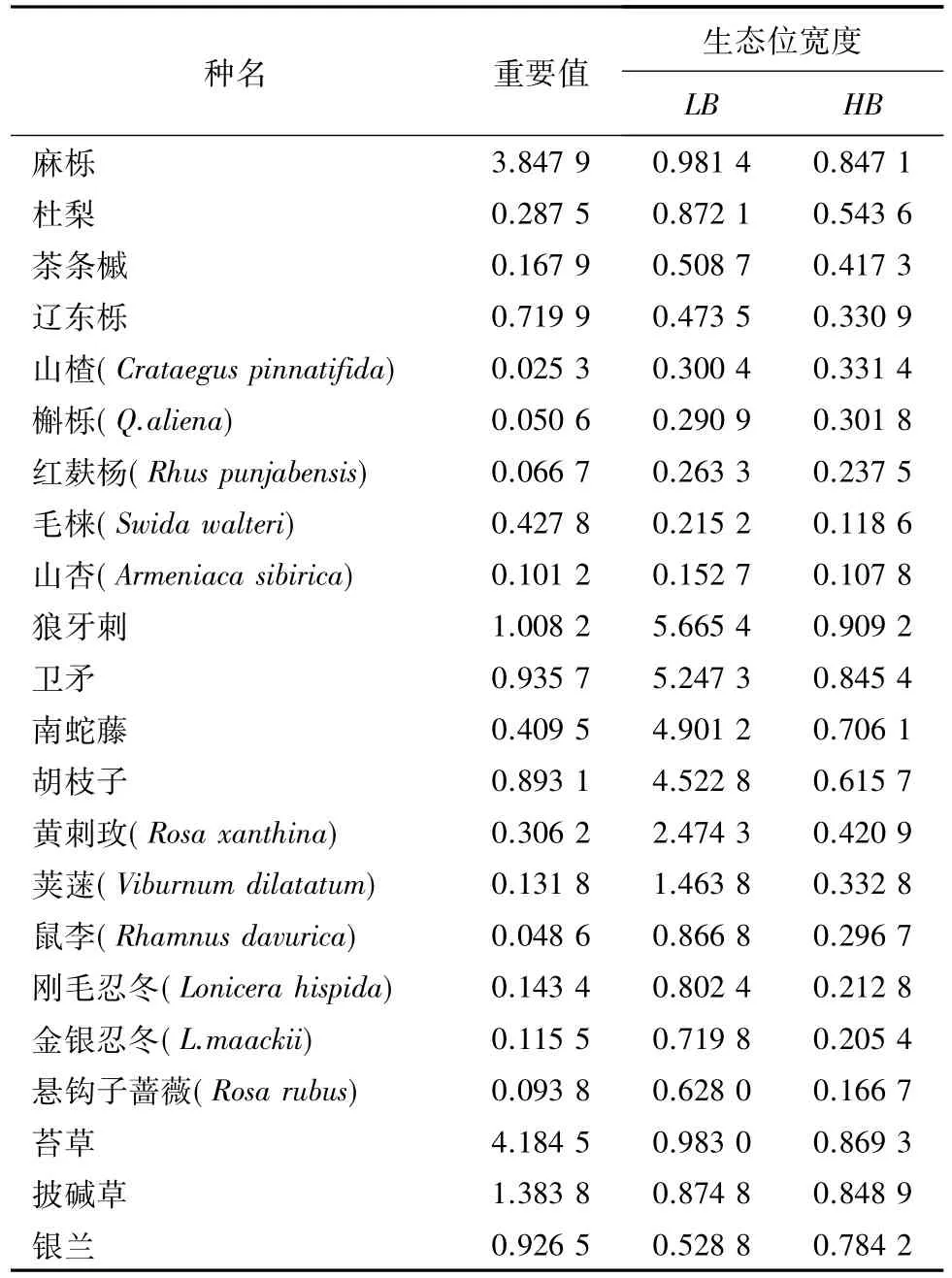
表2 麻栎林主要植物在阳坡的重要值及种群生态位宽度Table 2 Importance value and niche breadth of main plants species in Q.acutissima forest in sunny slopes
3.2 不同坡向对桥山重要植物生态位重叠的影响
阳坡乔木层生态位重叠占全部种对的94.44%,生态位重叠值在 0~0.973 2,而阴坡占 89.29%,重叠值在0~0.836 0(表4、表5)。阳坡和阴坡主要乔木层生态位重叠值>0.6的分别有18、9对,各占全部种对的 63.8%、32.14%;重叠值在 0~0.6(不等于 0)的都有16 对,各占 44.3%、57.14%,重叠值为 0 的各占全部种对的 5.6%、10.71%。
阳坡和阴坡主要灌木层的生态位重叠值处于0.6~1.0 的分别有 22、16 对,各占全部种对的48.89%、15.38%,在 0.4~0.6 的有 8、20 对,占全部种对的 17.78%、19.23%,0~0.6 分别有 21、74 对,各占全部种对的46.67%、71.15%,重叠值=0 的分别有2、14 对,各占全部种对的 4.44%、13.46%;
阳坡草本的生态位重叠值都集中在0.6~1.0,阴坡草本生态位重叠值在0.6~0.8有1对,占全部种对的16.17%,在0~0.6的有 5对,占全部种对的82.84%。
阳坡的茶条槭和红麸杨、胡枝子和金银忍冬、苔草和披碱草的生态位重叠值最大,分别为0.973 2、0.999 6、0861 2,阴坡的茶条槭和杜梨、卫矛和山楂、苔草和披碱草的生态位重叠值最大,分别为0.836 0、0.943 6、0.712 1,表明以上种对间对桥山林区生存环境的要求极为相似。在阳坡,生态位重叠值=0的种对有4对,而阴坡则有17对,这些种对无论是生态学特性还是对生境因子的要求都不相同。生态位重叠值低或者没有重叠的原因可能由于物种间生物生态学特性不同引起,也可能是生态学特性相同,为了共存而长期的种间竞争使得对环境资源的要求分化造成的。比如阳坡乔木层红麸杨和山楂为不同科属,生态学特性决定了物种间生存环境的差异性,在同一个资源位共同出现的频度较低甚至为0;山楂与山杏都为蔷薇科物种,生态学特性相似,为了共存而长时间的种间竞争使得对桥山林区资源环境的要求分化,导致种间没有重叠。

表3 麻栎林主要植物在阴坡的重要值及种群生态位宽度Table 3 Importance value and niche breadth of main plants species in Q.acutissima forest in shade slopes
4 结论与讨论
根据阴阳坡麻栎林群落内乔灌草的重要值和生态位宽度大小可知,一般重要值大的物种生态位宽度大,重要值小的物种生态位宽度小,而有的物种重要值大小与生态位宽度排序并不是一一对应,比如阴阳坡乔木层的辽东栎,重要值较大而生态位宽度却相对偏低。汪建华[15]等认为生态位宽度大小既与重要值之和的大小有关,还与其频度有重要联系,物种的生态位宽度与分布频度呈正相关[15]。
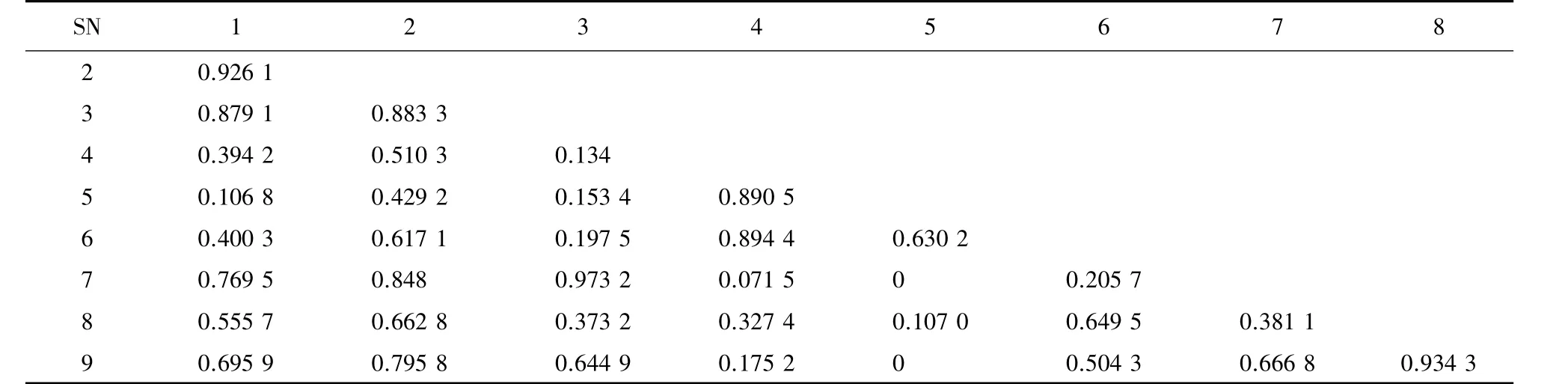
表4 麻栎林主要乔木在阳坡生态位重叠Table 4 Niche overlaps among the main trees in Q.acutissima forest in sunny slopes

表5 麻栎林主要乔木在阴坡生态位重叠Table 5 Niche overlapsamong main treein Q.acutissima forest in shade slopes
应用Levins和Hurlbert计算桥山林区天然麻栎林群落主要物种生态位宽度的趋势大致相同。麻栎林群落中麻栎和茶条槭为乔木层的优势种,狼牙刺、胡枝子和南蛇藤为灌木层的优势种,草本层的优势种为苔草,它们对桥山林区的环境资源充分利用,生态幅宽,分布范围广,生态位宽度大[16-17]。坡向影响了乔木层和灌木层有些物种的生态位宽度,比如乔木层的杜梨,杜梨为喜阳树种,对光照的要求比较苛刻,导致阳坡的生态位宽度高于阴坡。由于草本层生长在林下,对光照等生境条件的要求不高,所以草本的生态位宽度并未受到坡向影响。
有关研究表明,生态位宽度大的物种与其他种群的生态位重叠较高,生态位宽度小的物种与其他种群的生态位重叠较低[16-18]。也有研究表明,生态位宽度大的物种和其他种对间的生态位重叠不一定高,这是由于物种本身的生态学特性不同,它们对环境资源的利用不一致或者环境空间的异质性造成的[19-22],本研究也得到相同结果。除此之外,本研究发现生态位宽度较小的种对间可产生较大的生态位重叠,例如阳坡乔木层山杏和毛梾生态位宽度都比较小,生态位重叠反而较大,为0.934 3,虽然山杏和毛梾在样地内出现的频率低,生态幅窄,但是由于二者都为喜光树种,在适宜生长空间共存,或者是因为生存环境斑块较多导致它们之间的生态位重叠较高。所以物种的生态位宽度与生态位重叠没有绝对的相互关系。
阳坡麻栎林主要物种生态位重叠值>0.6的占全部种对的 51.20%,阴坡则占 18.80%,无论是乔木、灌木还是草本,阳坡生态位重叠值大多集中在0.5~1.0,而阴坡的则多集中在 0~0.5。生态位重叠值越大,表明种对间利用环境资源的能力越相似[23],在资源匮乏的情况下就会引起物种间竞争[24]。阳坡的麻栎林群落在桥山林区生境条件尤其是水分条件方面存在竞争,使得阳坡的有些物种在长期演替的过程中被淘汰,造成物种丰富度降低。通过勘测发现阴坡的麻栎林长势整体较阳坡好,从而阴坡为麻栎林生存的最佳适宜环境。麻栎对温度要求较高,随着辽东栎、茶条槭等耐阴树种的生长,林冠郁闭度增大,麻栎的生长优势会逐渐被取代,阴坡麻栎林可对乔木层辽东栎、茶条槭等伴生树种进行适当间伐,同时对狼牙刺、南蛇藤、胡枝子等生态位宽度较大的灌木进行适当的砍伐,通过抚育间伐来增大阴坡麻栎林林窗面积与光照强度,不断调整林分中物种关系,使林木能处于一种良好生存环境,实现麻栎林的可持续经营。
猜你喜欢
广东蚕业(2022年4期)2022-11-23
种子科技(2021年17期)2021-10-29
思维与智慧·下半月(2021年7期)2021-08-13
生态学报(2021年6期)2021-04-25
湖北农机化(2020年16期)2020-10-21
江西农业(2020年13期)2020-09-08
现代农村科技(2018年11期)2018-11-15
课程教育研究(2017年11期)2017-04-17
