
齐墩果酸在小鼠酒精性肝纤维化中的作用机制
2018-10-08 05:53冯世兵宋素贞王洪波
中国老年学杂志 2018年18期
王 宇 冯世兵 宋素贞 王洪波
(北京市和平里医院,北京 100013)
核转录因子相关因子(Nrf)2通过介导Ⅱ相解毒酶和一系列抗氧化酶,如谷胱甘肽-s-转移酶(GST)、血红素氧合酶(HO)-1、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的表达,从而降低各种药物、细菌、毒素及化学致癌物的肝毒性〔1~3〕,是肝脏防御氧化应激的最重要调节因子。齐墩果酸是五环三萜类化合物,广泛分布于自然界近200种植物中,生物活性较多〔4〕,试验证实具有保肝作用〔5〕。研究证明三萜类化合物对Nrf2有激活作用〔6〕。本研究通过检测Nrf2在小鼠肝脏中表达水平,探讨齐墩果酸抗纤维化作用分子生物学机制。
1 材料和方法
1.1材料 雌性昆明小鼠30只,野生型SPF级,体重20~25 g,在山东大学第二医院的动物实验中心饲养。美国Abcam公司抗Nrf2兔抗鼠多克隆抗体(ab53019)、纤维连接蛋白(FN),上海基因有限公司的兔/鼠通用型二抗免疫组化试剂盒(GK500705)、日本Olympus公司全自动生化分析仪等。
1.2动物模型制备和分组方法 实验小鼠均饲养1 w,温度、湿度等条件完全相同。适应环境后随机分为3组:A组(对照组):10只;B组(模型组):10只,每天应用浓度50%乙醇灌胃1次,12 ml/kg(4.8 g/kg);C组(治疗组):10只,乙醇灌胃方法同B组。齐墩果酸提前1天开始灌胃,1次/d,用量20 mg/kg。本实验获我院实验动物伦理委员会批准。
1.3标本采集 于12周末,天平称重,小鼠均12 h禁食(不禁水),给予水合氯醛麻醉(0.3 ml/100 g)腹腔注射,心脏穿刺方法取血,以3 000 r/min离心20 min(离心半径:104 mm)留取血清,-80℃冰箱保存。肝脏标本2/5部分以10%的甲醛溶液固定备用。3/5部分液氮快速冰冻后储存于-80℃冰柜中,用于制备RNA悬液等。
1.4血清生化指标的检测 血清标本送生化检测中心,由专业实验员使用生化分析仪分别检测小鼠血清丙氨酸氨基转移酶(ALT)、门冬氨酸氨基转移酶(AST)和总胆红素(TBIL)水平。
1.5病理学检查 肝组织常规固定、石蜡包埋、切片(厚度约3.5 μm)。小鼠均制作3~5张肝脏标本的组织切片,显微镜下根据HE及MT染色不同,选择不同的放大倍数,每片拍摄5个不互相重叠的视野。慢性肝损害评分标准参照Thompson分级,在低倍镜下(×100)观察肝细胞炎症及坏死情况。具体评分标准,炎症情况:无炎症细胞浸润为0分,炎细胞稀疏分布在坏死区域连接处为1分,炎细胞常见为2分,中性粒细胞占优势为3分,淋巴细胞占优势为4分。肝细胞损害情况:无肝细胞坏死为0分,以局灶性肝细胞坏死为主为1分,肝细胞呈现区域性坏死为2分,连续细条形或呈补丁样坏死为3分,大片样或融合的区域性肝细胞坏死为4分。采用盲法独立阅片由2名熟练病理学教授完成,取平均值。
1.6肝组织细胞外基质蛋白(FN)沉积情况。FN应用SP方法做免疫组织化学染色。胞质内出现黄棕色颗粒判定标准为阳性结果。应用图像分析系统(HPIAS)进行分析(每片随机选择不相重叠5个高倍视野)。
1.7Nrf2的检测 连续切片法将固定好的肝组织包埋后做4 μm组织切片,免疫组化采用EnVision染色法,PBS替代一抗作阴性对照,稀释浓度为1∶300,已知阳性片作阳性对照。肝细胞胞质出现棕色颗粒结果判定标准为阳性,每张组织切片随机选择5个不相重叠高倍镜视野,选 5个高倍视野计算阳性百分比的平均数。病理组织切片染色结果判定及阅片方法同上。
1.8数据处理 采用SPSS16.0软件进行方差齐性检验、ANOVA单因素方差分析、Mann-WhitneyU检验、非参数秩和检验。
2 结 果
2.1各组ALT、AST、TBIL水平比较 B组ALT、AST、TBIL均显著高于A组(P<0.05),C组ALT、AST、TBIL均显著低于B组(P<0.05)。见表1。

表1 ALT、AST、TBIL水平及Nrf2 表达情况
与A组比较:1)P<0.05;与B组比较:2)P<0.05,下表同
2.2各组HE染色结果比较 A组肝细胞形态、小叶结构和汇管区均未见异常。B组:肝脏小叶内炎症细胞浸润以淋巴细胞为主,偶见少量中性粒细胞浸润。肝细胞见散在的多发灶状甚至结节样坏死,变性严重。C组较B组组织学情况明显改善。见表2、表3、图1。

表3 各组肝细胞损伤比较(n=10,n)
2.3肝脏组织MT染色结果 A组肝脏组织纤维化(蓝色)极轻,仅有少量,位于血管周围。C组及B组纤维化范围形态学测量分析差异有统计学意义〔(1.86±1.23)%vs(3.55±1.37)%,P<0.05〕。B组纤维化范围形态学测量分析较A组(0.03±0.02)%显著增加(P<0.05)。B组肝组织见较多增宽的纤维组织分割并且相互连接,位于汇管区及中央静脉周围。C组纤维化分隔纤细,范围较B组明显减小,见图2。
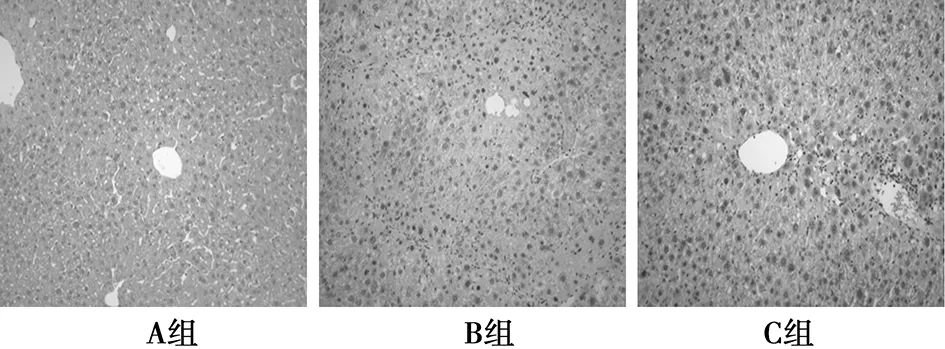
图1 肝组织炎症情况(HE,×200)
2.4FN免疫组化染色 A组极少量FN表达〔(0.05±0.03)%〕。B组FN表达较A组明显增强(P<0.05)。C组与B组FN表达差异显著〔(2.32±1.13)% vs (5.17±2.05)%,P<0.05〕。见图3。
2.5Nrf2表达情况 B组较A组Nrf2表达明显增强(P<0.05),而C组明显强于B组(P<0.05)。见表1及图4。

图2 肝组织纤维化情况(MT,×100)
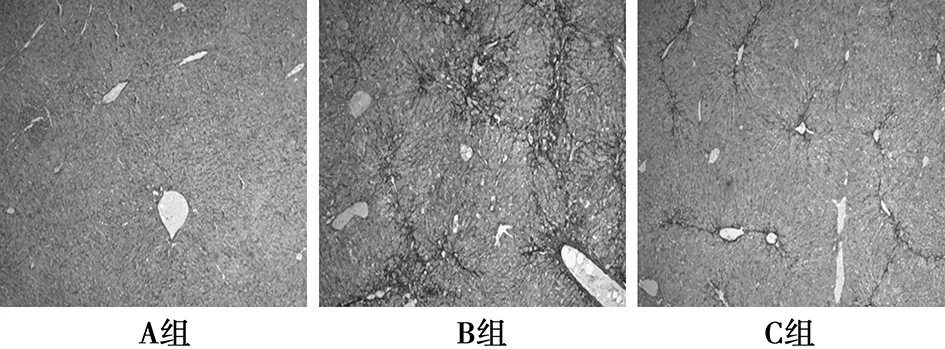
图3 肝组织FN表达情况(DAB,×100)
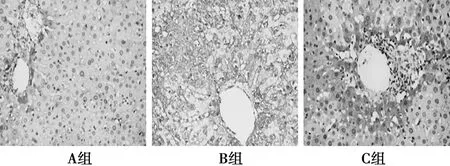
图4 肝组织Nrf2表达(DAB,×400)
3 讨 论
酒精性肝病是临床常见病,传统分为酒精性脂肪肝、肝纤维化、酒精性肝炎、肝硬化。本研究通过酒精灌胃方法模拟饮酒,利用乙醇引发氧化应激原理,12 w后成功构建了昆明小鼠肝纤维化模型。乙醇诱导产生的氧化应激机制是其在代谢过程中干扰能量代谢(线粒体三羧酸循环及脂肪酸代谢),产生大量活性氧自由基(ROS)、细胞色素酶P450(CYP2E1)诱导产生的羟乙基自由基(HER)、耗竭还原性谷胱甘肽继发抗氧化屏障损伤及耗竭Kuffper 细胞共同作用结果〔7〕。氧化应激反应导致线粒体膜的通透性增加,肝细胞对肿瘤坏死因子(TNF)-α的敏感性增加,从而促进肝细胞发生坏死或凋亡。ROS尚可引发脂质过氧化反应,组织蛋白变性和DNA的损伤。肝星形细胞(HSC)的激活引发了促纤维化因子的释放及胶原合成基因的表达,脂质过氧化物及HER继发各种免疫异常,导致肝组织慢性炎症“永久化”,最终发生肝纤维化〔8〕。Nrf2属于b-ZIP家族,是调节抗氧化应激反应的重要转录因子。Nrf2调节肝内诸多细胞抗氧化基因和Ⅱ相解毒酶等基因的基础表达和诱导表达,生理状态下与胞质中的抑制因子Keap1结合处于抑制状态。Keap1在C273等部位存在巯基,氧化应激导致其失去电子,形成二硫键,从而构型发生改变,Nrf2与Keap1解耦联后转移入核与抗氧化反应元件结合,启动下游的抗氧化酶基因及Ⅱ相解毒酶的表达,包括GST、HO-1、SOD、CAT,醌氧化还原酶(Nqo)1等,增加细胞抵抗氧化应激的能力〔9〕。同野生鼠(Nrf2+)相比,Nrf2基因敲除小鼠(Nrf2-)对四氯化碳损害修复延迟,炎症反应及纤维化程度明显增强〔10〕,对乙酰氨基酚引起的肝损伤较敏感,脂质过氧化和DNA损伤增加,肝细胞坏死程度和乙酰氨基酚毒性反应均增加〔11〕;肝脏切除后的肝细胞增生延迟,凋亡增加〔12〕。作为引起肝细胞损伤最常见的氧化应激因素的乙醇能诱导Nrf2表达。Lamle等〔13〕研究表明这是肝细胞对氧化应激一种适应性保护反应。齐墩果酸能显著促进酒精灌胃小鼠肝细胞Nrf2基因表达,推测在与氧化应激密切相关的肝纤维中,齐墩果酸保护肝细胞作用机制是通过增强Nrf2表达实现的。研究表明三萜类化合物可作为Nrf2基因的特别诱导剂,其通过激活Nrf2抑制黄曲霉毒素诱导的肝脏癌变过程〔14,15〕。CDDO-IM是一种人工合成三萜类化合物,能显著降低内毒素诱导的炎性反应水平及死亡率,具体机制与激活Nrf2途径有关。CDDO-IM预处理小鼠能通过诱导Nrf2及下游的抗氧化基因表达减轻乙酰醋氨酚导致的各种肝损害〔6〕。
乙醇代谢过程中产生了大量的ROS及HER,导致肝组织大量炎细胞浸润及促炎因子产生,HSC被激活转化,合成大量FN;细胞外基质在窦周间隙的大量沉积导致纤维化发生。本研究证实,齐墩果酸能显著降低酒精灌胃小鼠的血清ALT、AST、TBIL水平,减轻肝细胞的炎症损伤和肝组织纤维化过程。其抗纤维化分子机制可能是通过增强Nrf2的表达,加强Ⅱ相解毒酶和各种抗氧化基因表达,减弱氧化应激,减少炎细胞浸润,阻止HSC的激活,从而抑制细胞外基质合成并增加降解,减少细胞外基质(例如FN)的产生和蓄积而实现的。Nrf2对肝细胞增殖与凋亡、信号传导通路及活化等作用机制是未来研究的方向。齐墩果酸有效成分(三萜类化合物)提纯,人工合成及改良研究也是未来研究方向。
猜你喜欢
现代实用医学(2021年12期)2022-01-26
皮肤病与性病(2021年3期)2021-07-30
中国当代医药(2018年12期)2018-06-16
天然产物研究与开发(2018年5期)2018-06-13
中国药房(2017年13期)2017-05-16
中国比较医学杂志(2017年3期)2017-01-17
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中国比较医学杂志(2015年9期)2015-05-11
中国当代医药(2015年2期)2015-03-11
中国当代医药(2015年8期)2015-03-01
