
牡丹组织培养技术研究进展
2018-09-17 12:52:14李昱莹郭丽丽廉小芳郭大龙侯小改
浙江农业科学 2018年9期
李昱莹,郭丽丽,廉小芳,郭大龙,侯小改*
(河南科技大学 a农学院,b林学院,河南 洛阳 471000)
牡丹(Paeoniasuffruticosa)属芍药科芍药属,多年生落叶亚灌木,是我国特有的植物[1],主要分布于我国中部、西南部至西北部[2-3]。自古以来,牡丹象征着国家繁荣兴盛,民族幸福吉祥。具有很高的观赏、食用,以及药用价值。国家卫生部监督局于2011年3月指出可将油用牡丹凤丹和紫斑作为新型木本粮油植物资源进行开发和利用[4]。牡丹籽油已逐步发展成我国重要的食用油,与其他油料作物相比,具有高产出(667 m2综合效益达万元)、高含油率(籽含油率≥22%)、高品质(不饱和脂肪酸含量高于90%,α-亚麻酸高达42.8%,脂肪酸比例均衡)、低成本(耐旱耐寒耐贫瘠)等特点。因此,大力推广油用牡丹的种植对促进我国粮油生产、改变食用油消费结构以及保障国家粮油安全均具有重要意义,其发展前景非常广阔。
牡丹的有性繁殖为种子繁殖,但播种后代存在分离变异,不能较好地保存母本优良性状[5],且其种子存在生理后熟和二次休眠现象,导致播种育苗周期较长[6];其无性繁殖主要是分株、扦插、压条、嫁接等[7],但存在繁殖系数低、成苗周期长、出苗量少且质量参差不齐等问题。与传统育苗技术相比,组织培养技术有利于牡丹育种和快速繁殖[8]。因此,通过组织培养技术解决牡丹常规繁殖过程中存在的问题,将是油用牡丹发展走向商品化和产业化的必然趋势[9],也是加速其繁殖和育种的有效措施。
牡丹组织培养可在较短时期内扩繁大量优良品种,并能利用遗传转化技术在短期内培育牡丹新品种[10],可以在一定程度上打破传统育种技术固有的局限性。时至今日,关于牡丹组织培养的研究主要集中在愈伤组织诱导、适宜生长调节剂配比、外植体选择以及培养基完善等方面,但是牡丹快速繁殖(即微繁殖)体系尚未被确定,因此仍然无法满足牡丹新品种培育和优良品种推广栽培的要求。此外,组织培养过程中仍存在污染严重、继代培养中易出现褐化以及玻璃化现象、生根困难、移栽成活率低等问题亟待解决[11]。本文从外植体选择及分化途径,影响增殖和生根的主要因素等几个方面概述了近年来牡丹组织培养的研究进展,同时论述了组织培养过程中污染、褐化以及玻璃化等问题出现的原因及其可能的解决方法,以期为今后开展牡丹组织培养新技术的研究奠定基础。
1 牡丹再生体系研究
1.1 外植体的选择及分化途径
外植体的选择是植物组织培养过程中最重要的一步,近年关于牡丹的组织培养方面,国内外研究学者均有大量报道[12]。可供选择的外植体种类繁多,其中主要有芽、茎尖和茎段、胚和胚珠、叶片和叶柄、花药和花丝等,但不同外植体具有不同的分化能力,同一外植体的采摘时间和处理方式不同,其诱导的结果也不同。另外,不同的发生途径对外植体也有一定要求,因此将外植体选择与发生途径结合起来,才能建立正确有效的组培体系。
1.1.1 外植体的选择
牡丹组织培养的研究开始于1965年。随后Bouza等[13]发现了选择牡丹叶和腋芽组培最有效的细胞分裂素,从而推动了国内外对于全方位了解牡丹组培的高潮。在国内,李玉龙等[14]最早开展了牡丹组织培养技术的研究,其主要研究青龙卧墨池和十八号两个品种不同外植体的诱导情况。现阶段关于建立牡丹无菌体系的研究大都集中在外植体类型、取材时间、品种以及消毒方式4个方面。
外植体类型。自从德国植物生理学家和植物学家Haberlandt提出了植物细胞全能性的概念之后,组织培养作为一种高效的植物快繁技术显示出了巨大的应用价值,即任何外植体都具备在适当条件下诱导生成完整植株的能力。但研究表明,同一植株的不同外植体在器官发生能力上呈现出较大的差异。有研究表明,在牡丹组织培养过程中,土芽和幼芽的诱导率最高,叶片、叶柄次之,心皮和雄蕊却未能诱导成功[15]。
外植体取材时间。关于牡丹芽取样时间的研究表明,2月份获取的外植体进行培养时,污染率低,萌发时间早,植株健壮,培养效果最好;相反,8—10月的外植体在诱导时不仅极易污染,而且萌芽缓慢,后期叶片发黄且生长瘦弱,容易出现褐化死亡等现象[16-17]。这是因为此时的外植体尚未经历休眠,芽体内营养物质少;且对2月份取材而言,冬季的自然低温可将土壤及植株上已附着的病菌、细菌等微生物冻死,从而减少了外源污染;另外,冬季自然低温致使植株及芽内的激素含量产生动态变化,例如IAA、GA3等促进萌发和生长的激素类含量增加,ABA等抑制萌发和生长的激素类含量减少[18],从而改善了芽外植体的培养效果。
关于不同时期的花药诱导愈伤组织的研究结果表明,不同时期的花药诱导率也不尽相同,单核中期花药诱导率最高,其次是单核晚期,而单核早期和双核期均明显低于单核晚期[19]。可能的原因是当花药发育到四分体时期时,还没有发育完全,此时并不适合用来诱导愈伤组织。当达到单核中期时,细胞的生命活力最强,供给的营养也较为充沛,且细胞马上开始有丝分裂,因此对诱导条件最为敏感,此时的花药细胞极易发生染色体轴向的改变[20],从而进入脱分化分裂,导致单核中期花药最易产生愈伤组织。
外植体品种。研究发现,不同观赏品种的牡丹,其诱导结果会存在一定的差异。当以豆绿、黑花魁、大胡红和金星雪浪为材料进行丛生芽诱导时[16],前两者增殖系数较高、诱导分化出的丛生芽数量较多,而后两者的增殖系数却相对较低。各个品种之间诱导差异显著,一些品种极易体外培养成活,一些品种出现褐化、玻璃化现象而使成活率降低,还有一些品种表现出体外发育迟缓的现象,并且这种基因依赖现象还会影响到后期的继代培养表现[21]。由此可见,基因型的差异对牡丹组织培养的影响也较大。
消毒方式。外植体的消毒灭菌是组培技术中最重要的环节之一,不同品种、不同类型的外植体,其最适消毒方法均不同。常用的消毒剂有0.1%~0.2% HgCl2[22]、70%~75%无水乙醇[11]、次氯酸盐[23]等。一般的灭菌程序均为先对新鲜采集的外植体进行表面基本灭菌,然后利用合适的消毒剂消毒,最后用无菌水冲洗残留消毒剂。有的外植体消毒困难,会有二次或多次灭菌[17]。
唐豆豆等[22]研究表明,使用0.1% HgCl2对凤丹根蘖芽与鳞芽消毒8 min时,效果最好;而消毒过程持续超过10 min时,消毒剂对外植体本身的伤害较大,导致后期苗成活率较低。另外有研究表明,最好的消毒方式是直接使用升汞消毒[24],但也有人认为无水乙醇与升汞交替使用效果最佳[25]。对牡丹而言效果最好的消毒方式是75%无水乙醇浸泡40~50 s,无菌水清洗后用0.2%升汞消毒 6~8 min[26-27]。不过也有研究表明,使用生物杀菌剂处理牡丹鳞芽时能达到100%消毒,且对材料无伤害,优于使用HgCl2消毒[28]。此外消毒方式可以间接影响组培苗的褐化现象,研究显示,与使用升汞消毒相比,使用NaClO会导致外植体的褐化程度加深[23]。
1.1.2 腋芽萌发途径
大多数进行组培快繁的植物,其外植体器官发生途径均为腋芽萌发途径,这种途径是不经过脱分化产生愈伤组织后再生,而是直接由其分生中心分化出植物器官,最终成长为完整的植株。腋芽萌发途径能很好地保留母本的优良性状,虽然变异率小,但增殖率通常较低且繁殖系数不高。由腋芽萌发途径建立的组培体系中,外植体部位和外植体的预处理是影响培养成功与否的关键因素。牡丹组培常用的芽有顶芽[29-30]、腋芽[31]、萌蘖芽[32-33]。大部分的研究认为,不同部位的芽的诱导率有明显的差别,其中萌蘖芽[32]诱导能力最强,花芽分化能力最低,可能是萌蘖芽组织分化程度较低,导致其诱导增殖能力最强。另外低温预处理离体芽尖[33]后,其萌发率及发育速率会再次提高,且低温处理之后外植体污染率及褐化率明显降低。
1.1.3 间接器官发生途径
植物的间接器官发生途径是指外植体在一定条件下经过脱分化诱导后产生愈伤组织,而后再由愈伤组织分化出不定芽或不定根,从而形成完整植株。由于脱分化所形成的愈伤组织可在较短时间内重新诱导分化出大量新生植株,因此,通过诱导愈伤组织,可使外植体的繁殖系数大量增加,完成快速扩繁。但愈伤组织细胞团的遗传性质是不稳定的,而且随着继代时间的增长,它们的分化再生能力会逐渐下降,甚至完全消失。近年来关于牡丹组织培养的研究也有以此途径诱导再生植株的案例,在愈伤组织诱导过程中,外植体类型、培养基种类、激素配比和培养条件均对培养结果有不同程度的影响。目前,牡丹组培中愈伤组织诱导常用的外植体有茎尖和茎段[34-38]、叶片和叶柄[39-41]、花药和花丝[42-43]、种胚[44-45]、根韧皮部[46]等。研究表明,牡丹不同外植体诱导出的愈伤组织存在高度异质性[47]。
近年来,以牡丹茎尖为材料进行的组织培养也取得了一些进展。李志军等[34]用大胡红茎尖进行实验时发现,30 d后每小段的簇生芽可达4~6个,说明其茎尖分化能力较强。随后,在不同牡丹品种茎尖和茎段[10,35]诱导实验中进一步证明,茎尖分化能力较强。研究发现,适合愈伤组织分化的培养基为MS+0.5 mg·L-1BA+0.01 mg·L-1NAA。另外,研究学者也进行了大量的不同品种牡丹茎段诱导愈伤的研究实验[36-37],研究结果发现,以茎段为外植体诱导愈伤组织时易出现褐化现象。朱向涛[38]用江南牡丹茎段诱导愈伤组织时发现在愈伤组织分化过程中,加入0.2 mg·L-1KT效果最好,除此之外有机物的增加也有助于促进愈伤组织分化,其中以加入300 mg·L-1水解酪蛋白效果为最佳。
叶片是植物进行光合作用的重要营养器官,而叶柄是茎段培养的重要材料,二者作为外植体可加速良种植物的保存,解决不能用种子繁殖或难以用种子保存的种质资源保存[48]。安佰义[39]用无叶柄幼叶作为外植体诱导愈伤组织,研究MS,B5和WPM 3类培养基的适用性,结果发现WPM培养基更适用于该外植体愈伤组织的诱导。李丽霞等[40]以大胡红和荷花紫的幼叶为外植体,用水解酪蛋白和4种植物生长调节剂(BA、2,4-D、KT、NAA)诱导愈伤组织,结果发现愈伤组织对2,4-D最为敏感。张健欣[41]分别以紫斑牡丹的大田和无菌苗叶片、叶柄为材料,研究NAA在愈伤组织诱导中对其诱导率的影响,结果表明,叶片和叶柄诱导愈伤组织时NAA的最佳浓度为0.3 mg·L-1和0.4 mg·L-1;另外不同外植体愈伤组织诱导率表现出明显的差异,诱导愈伤的能力水平为无菌苗叶柄>大田叶柄>无菌苗叶片>大田叶片。
牡丹具有多核或多细胞花粉,说明其具有良好的诱导潜力,因此花粉诱导是获得单倍体和纯合体的有效途径[49]。当前,对于牡丹花药和花粉培养仍停留在诱导出愈伤组织阶段,并没有诱导出完整植株,说明牡丹花药和花粉培养还处于起步阶段。朱向涛等[42]以凤丹带花丝的花药为材料诱导愈伤组织时发现单核中期为其取材的最佳时期,在4 ℃低温条件下对外植体进行预处理后,其愈伤组织诱导率可达50.8%。刘会超等[43]以乌龙捧盛花药为外植体,研究不同培养基和激素组合对其愈伤组织诱导率的影响中发现,MS培养基为最佳,Miller培养基次之,B5培养基则较差,其诱导愈伤组织的最适培养基为MS+2 mg·L-12,4-D+1 mg·L-16-BA+0.5 g·L-1水解乳蛋白。
胚培养是指无菌条件下将植物的种胚接种在适宜的培养基上使之发育成苗的一种组织培养技术。该技术不仅可以减缓胚败育和发育不良的情况,也可以缩短休眠期,加快种子发育、促进萌发,另外胚的离体培养可将萌发时间缩短28~42 d[50-51]。Demoise等[52]利用牡丹种子成熟胚成功地诱导出愈伤组织,使更多研究人员开始重视对牡丹的胚培养。研究发现,不同发育时期的胚诱导愈伤组织的情况也有差异,幼胚分化程度低,其诱导率比成熟胚高[44]。激素的选择在一定程度上也有很大的影响,在添加2,4-D和TDZ的培养基上,日本品种太阳已成功地由胚诱导出愈伤组织[53]。另外,H2O2等外源物质的使用对牡丹胚的诱导也有促进作用[45]。
胚珠培养是指在无菌条件下培养子房中所取得的胚珠,使胚珠直接或通过诱导愈伤组织、不定芽、珠心及其他器官获得试管苗[50]。胚珠培养具有缩短发芽时间,提高萌发率,保存杂交胚胎和加快育种的能力。2006年何桂梅等[54]首次报道关于牡丹胚珠培养的研究,指出花后48 d的胚珠很难培养,可能是因为没有完成受精或在培养基中没有足够的营养物;花后60 d的胚珠(胚胎完全分化)在MS培养基中,可以发育成苗,但需要及时移除胚乳。
1.1.4 体细胞胚胎发生途径
体细胞胚发生途径最早是在针叶树种离体培养研究中获得的,一些木本植物现已成功建立了通过此途径获得再生植株的体系。目前,能成功诱导出体胚的牡丹品种很少,且诱导率普遍较低[55-65]。对于油用牡丹品种凤丹的研究也仅在殷丽青等[44]和周秀梅[57]的论文中有所提及,但所提出的研究结果均显示体胚诱导率极低,且大多数体胚诱导成功之后因不能继续生长,导致后续实验无法开展。由此可见,牡丹的体细胞胚诱导仍然较为困难。
BMD提供了支持定理1中对函数进行A变换的算术运算[12],如A(xi∧xj)通过算术与实现,A(xi⊕xj)通过算术异或实现,并且可以实现定义6中的“在A变换过程中利用幂等律抑制随机变量的指数成分”.对函数f的逻辑表达式实施相应的算术运算即可得到A(f)对应的BMD,将随机变量Xi的值代入,进行BMD求值[12]即可得到f的信号概率.BMD可以较高效率地表示和操纵类似于概率表达式的代数表达式,并且BMD求值的复杂度为线性复杂度[12],因此本文在底层使用BMD表示概率表达式.
外植体的种类、基因型和前期预处理等均对体细胞胚的诱导有影响。王莹等[58]把不同收获期的紫斑种胚作为外植体进行培养时发现,种子成熟度不同对组培苗的生长有一定影响。大部分研究表明,牡丹体细胞胚发生的最佳时期是花后90~110 d[59]。朱向涛等[60]用合子胚、胚轴和子叶作为外植体进行培养,从而探究不同外植体的诱导情况,结果表明,种胚的诱导率最高。Brukhin等[61]研究发现,以球形胚阶段后的胚状体为外植体时,其繁殖能力最强;而心形胚和鱼雷形胚只有25%能诱导成株;在诱导愈伤组织时,成熟种胚、鱼雷形胚和心形胚的诱导率依次降低。另外,多数学者研究证明激素配比对体细胞胚诱导的影响也极为重要。
1.2 影响增殖的主要因素
1.2.1 培养基
由于牡丹基因型及外植体选择的多样性,其诱导过程中所选培养基种类也不尽相同。主要的基本培养基有B5[62]、LP[63]、N6、LM[64]、DCR[61]、MS[65-66]和WPM[67-68]培养基,牡丹组培中使用最普遍的培养基是MS及各种改良的MS培养基,也有用WPM作为基础培养基。各种培养基内所含无机盐、有机营养物以及外植体生长所需的碳源和氮源有所不同,其种类和成分直接影响到组培苗的生长[69]。
初代培养指外植体接种后最初的几代培养,其目的是为了获得无菌材料和无性繁殖系。初代培养常用诱导培养基,培养基中的外源生长激素主要为细胞分裂素,生长素含量则相对较少。安佰义[39]在研究不同培养基作用于丛生芽诱导的实验中,发现低温处理后的外植体在WPM培养基上丛生芽诱导率最高,MS培养基次之,B5培养基最低。王新等[70]用凤丹鳞芽培养时,也证实了WPM培养基诱导率最高。但也有研究显示,初代培养基为MS时,外植体生长发育快,叶柄长;培养基为WPM时,其生长发育减慢,且叶柄较短[71]。马俊[72]也表示,在牡丹的初代培养中,比较理想的培养基为钙离子浓度加倍的MS培养基。也有人认为关于牡丹最适的初代培养基是半强度MS培养基[73-74],其诱导丛生芽的效果显著优于MS基本培养基。
继代培养是指在初代培养获得无菌材料后再进行的连续数代的扩繁培养过程。其目的是为了得到一定数量的无根苗,然后用获得的无根苗边繁殖边生根,其后代数量按几何级数快速增加。继代培养基通常选用分化培养基,而且不同外植体在继代时所需要的培养基也不同。王燕霞等[75]以洛阳红为试材研究6种不同培养基(MS、1/2MS、C17、WPM、B5、DKW)对继代苗增殖生长的影响时发现,DKW培养基最适合其继代增殖。孟清秀等[76]研究表示,牡丹品种菱花湛露的最佳增殖培养基为改良WPM培养基。张桂花等[30]通过对豆绿、大胡红、黑花魁和金星雪浪的鳞芽进行培养研究,认为MS为最佳继代培养基。
继代培养的周期同样也对牡丹组织培养产生影响,多数研究表明,在第3~4周,牡丹组培苗的质量最佳,生长量大;第4~5周,组培苗生长量增加不大,但不定芽可以充分生长。因此,牡丹组织培养以4或5周为继代周期较好[77-78]。
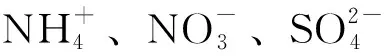
植物组织培养过程中生长调节剂的配比非常重要,直接影响苗和根的形成[79]。不同植物在不同生长阶段其所需要的生长调节剂种类和配比均不相同,另外,不同植物生长调节剂配比不同,其诱导效果也不同,其中生长素和细胞分裂素的使用起着至关重要的作用。牡丹组培常用的生长素有NAA、2,4-D、IAA、IBA;细胞分裂素有6-BA、KT、TDZ、ZT。另外,赤霉素(GA3)、脱落酸(ABA)在牡丹组织培养中也很重要。
不同种类的细胞分裂素在牡丹组织培养过程中的影响不同,大量实验研究表明,6-BA[80-81]是对牡丹叶芽最为重要的细胞分裂素,与其他激素同时使用效果会减弱,在增殖培养中只有6-BA能促进不定芽的发生。随后更多的实验证实,6-BA和GA3同时使用效果较单独使用6-BA更好,但在低钙浓度下会出现茎尖和叶片枯死。因此,不能长时间使用同时含有6-BA和GA3的培养基,应尽可能地间断使用[68]。另外6-BA、KT和TDZ单独使用可促进牡丹腋芽发育[82-83],但增殖率都比较低,低浓度的TDZ、6-BA和KT合用能显著提高腋芽形成和丛生芽形成。另外不同激素组合可以对牡丹增殖系数产生较大差异的影响,研究人员利用极差分析结果表明,6-BA、KT、NAA 3种激素对增殖数的影响逐渐降低[35]。
生长素有利于营养器官伸长生长,可以使细胞进入持续分裂的增殖状态。牡丹鳞芽增殖实验表明,NAA对其影响比较显著[84];对于牡丹组培苗生根的研究中发现,IBA的正向效果极其显著[39]。在牡丹组织培养研究中发现生长素与细胞分裂素有协同作用,搭配使用时其浓度比例不同诱导效果也不同,一般来说细胞分裂素高时有助于芽的分化,细胞分裂素含量较低时,则有助于根的形成。
1.2.3 培养环境条件
除外植体及培养基种类之外,湿度、温度、光照和pH值等因素也严重影响着牡丹组织培养结果的成败。现阶段公认的最适合牡丹组织培养的环境条件如下:温度为(25±1) ℃,光强为1 500~2 000 lx,光照时间是每天10~12 h,培养基pH值为5.8~6.0。
湿度是组培过程中一个较重要的因素,研究表明,将湿度控制在70%上下较为适宜[85],因为过低会使培养基渗透压升高,影响植株健康生长;过高则易造成污染。另外温度的控制也很重要,研究发现24~26 ℃时,试管苗生长快、长势壮、且芽分化多,表现最佳[30]。
光照因子对牡丹组培的影响较多,主要有光强、光质、光照时间等,大部分的研究表明,光强为1 500~2 000 lx,光照周期为12 h的光照条件最佳。对于光质的现有研究则表明,70%红光+30%蓝光的冷光灯处理35 d后,组培苗的叶片数、株高及茎长等均达到理想效果[86]。
1.3 影响生根的主要因素
生根是组培过程中非常重要的阶段,受多种因素影响,包括培养基成分[87-88]和环境因素[89-90]。生根困难是牡丹增殖体系确定过程中最受关注的问题,包括生根质量差以及体外培养存活率低[91]。因此,生根问题的解决将是牡丹组培中一个很重要的突破。
总结所有研究发现,牡丹组培苗诱导生根可分为 3种:(1)速蘸法,将组培苗浸泡在高浓度IBA溶液中,迅速接种到只含活性炭不加任何生长调节物质的培养基上;(2)一步生根法,外植体诱导生根时在含有低浓度IBA的培养基中连续培养[22];(3)两步生根法,先将组培苗在含高浓度IBA的培养基上进行根诱导培养,然后在仅含有活性炭的培养基上生根。大量实验证明,两步法是目前为止最常用且效果最好的方法[92]。此外蔗糖浓度也严重影响组培苗的生根数量,充足的糖类化合物能提高牡丹组培苗的生根率,并同时促进根和茎的生长[93-94]。
牡丹内含有大量的酚酸化合物,对其体外生根有一定的影响[95]。有报道称酚酸在植物生物和非生物胁迫下积累[99]并控制不定根的形成和发展[96-97]。现有研究发现,牡丹组培苗中含有12种酚酸化合物,内源性酚酸含量的变化与其体外生根密切相关,其中二元酚有利于体外生根[98]。
2 制约牡丹组织培养发展的主要因素
2.1 污染
污染现象在组培过程中经常出现,且一旦出现污染就容易造成失败。易引起污染的原因有很多,如接种人员以及外植体带菌、操作环境及接种工具灭菌消毒不彻底、操作不规范、外植体内生菌较多等等。因此,实验过程需严格按照操作要求进行,杜绝接种前污染。组培室以及操作间需定期进行消毒,经常用酒精喷雾降尘消毒,并用紫外线照射消毒灭菌。对于接种后的污染,要根据外植体种类的不同寻找不同的消毒方式进行预防,选择合适的方法和消毒剂,适当情况下可进行多次灭菌。植物内生菌导致的污染,则可在培养基中加入适量的抗生素或适宜的抑菌剂,从而降低污染率。
2.2 褐化现象
褐化是组织培养中最重要的问题,其会引起外植体代谢紊乱,最终将导致死亡。外植体的品种、取材部位、年龄、取材时间以及培养基种类、组培室温度等等均与褐化相关。研究表明,年龄老、高度分化、木质化程度高的外植体材料很容易褐化,而较嫩、分化程度低的材料褐化程度相对较轻[99]。牡丹是多糖多酚植物,继代时外植体基部愈伤被切除,切口处的酚类物质会被氧化形成深褐色的醌类物质,这些物质在多酚氧化酶、酪氨酸酶等作用下,导致其他代谢通路不稳定,最终使外植体衰老死亡。由于各个品种牡丹植株内的总酚含量不同,因此关于影响牡丹褐化的因素,研究人员们做了大量研究。牡丹花型繁多,花型越复杂,越容易发生褐变,这是因为植株自身多酚氧化酶含量是随花型演化程度的增加而增加的。而且温度会影响多酚氧化酶的活性,研究表明,木本植物在温度较低的冬春季取材褐化率较低;一般越小的外植体褐化越严重,故而选取适宜大小的外植体可有效降低褐化率。
另外培养基成分、培养条件及抗褐化剂的添加也会对褐化造成一定程度的影响。不同培养基添加的无机盐离子浓度各不相同。牡丹叶柄培养常用的 3种培养基:WPM、1/2MS和MS培养基,其培养过程中外植体褐化程度依次加重[100]。因此,外植体褐化程度与培养基的无机盐浓度呈正相关。在培养过程中适当的低温[101]和黑暗[27]处理可以缓解褐化的发生,这是由于温度和光照与多酚氧化酶的活性相关。此外在黑暗条件下,6-BA能引起多酚氧化酶活性提高,而2,4-D和IAA可以减少多酚合成[102],所以适当的激素配比也可以减轻褐化发生。由于牡丹组织培养发生褐化是一种酶促反应,选择合适的褐化抑制剂、抗氧化剂和吸附剂可以抑制酶的活性,从而使褐化程度降低。常用的有抗坏血酸(Vc)、AgNO3、Na2S2O3、PVP、活性炭(AC)。Vc可以将Cu2+还原为Cu,而后产生的螯合物可以抑制酶的活性;吸附剂对褐化的抑制作用为 PVP>AC[100],且AC容易聚集在外植体伤口处,阻碍其吸收营养。
2.3 玻璃化现象
玻璃化是植物在组培过程中极易出现的生理病变或生理失调现象,多表现为叶片纵向卷曲或肿胀,表皮角质层蜡质缺少,叶片脆弱易碎;组培苗含水量高,存活率低。已报道80多种植物组培时出现玻璃化[103]。玻璃化的牡丹苗叶柄较粗,叶片肥厚,半透明状,植株呈充水状,与冻害植株形态极其相似。由于玻璃化现象的发生将导致组培苗病变死亡,因此解决玻璃化现象将有利于提升组织培养技术的成功率。

2.4 生根困难和移栽成活率低
目前,牡丹组织培养体系建立过程中最难解决的问题仍在组培苗生根以及驯化移栽过程。因为牡丹组培苗生根率低且生根质量差,且大多数不定根由愈伤产生,不定根与植株之间没有直接连接,另外生根诱导过程中外源IBA的使用会导致植株内源ABA大量积累,继而导致植株分裂能力下降,出现休眠现象,所以移栽成活率低。为了建立合适的驯化移栽体系,研究人员作了大量的工作,包括组培苗生根状态、移栽季节、基质的选择以及后期管理等方面的关键技术。研究证实,待生根后1个月,根长达到3~4 cm时移栽成活率最高[105],主要是因为过早移栽植株适应力差,抵抗力弱;过晚移栽时根系木质化程度大,吸收营养物质能力变差。移栽时应选择昼夜温差大、温度适宜的秋季,以便移栽苗健康生长。
试管苗种植成活过程中另一个重要的因素就是基质的选择。一般而言,基质的质地要疏松透气,同时有良好的保水性,还要容易消毒等。常用的基质有蛭石粉、珍珠岩、草炭土、腐殖质等。通常认为蛭石粉+草炭土(1∶1)更适宜组培苗生长[34]。
2.5 成本高
组织培养技术虽然在一定程度上可以缩短传统育种年限,短时间内获得大量幼苗,达到一定经济效益,但总体来看,其成本仍然相对较高。无论是培养基、外植体、蔗糖等物资,还是人力的消耗,再加上组培苗成活率低、移栽困难等问题导致其处于高成本技术范畴。
3 组培新技术
常规的组织培养中,研究人员一直将重心放在培养基筛选、激素配比及外植体的种类选择等方面。事实上光照、温湿度、CO2浓度等对小植株生长的影响也很大,因此研究人员提出了关于组培的新技术:光自养培养技术/无糖培养技术,植物开放式组织培养以及开放式光自养微繁技术。
光自养培养技术(photoautotrophic micropropagation)又称无糖培养技术(sugar-free micropropagation),是指组织培养过程中改变碳源供给途径,并控制环境因子,从而促进植株光合作用速率,使外植体生长由异养型转变为自养型[106]。光自养培养技术不需要蔗糖为碳源,大量降低了杂菌的繁殖速率;且培养容器向大型化发展,降低了传统培养中,容器较小,环境较密闭,瓶内气体不流通,相对湿度较高等问题;另外,传统培养中多使用琼脂等凝胶物质为支撑材料,组培苗根系脆弱,驯苗困难,而该技术多使用塑料泡沫、蛭石、成型岩棉等多孔性无机材料,价格低廉且大多可重复使用,很大程度上降低了成本。而且这些材料通气性好,可有效促进植株根系生长。目前,国内外学者对于光自养培养技术的研究多集中在培养室环境[107-109]、培养容器大小[110-111]、CO2的输入[112-113]、支撑材料[114]等等。研究表明,与传统组织培养相比,光自养培养获得的组培苗生长快且长势较好,畸形苗少,生根率和成苗率较高。另外,培养过程中污染率大幅降低,使得试管苗驯化期间的成活率大幅上升。
植物开放式组织培养是指无需高压蒸汽灭菌,在超净工作台外的操作环境用普通容器进行接种工作的新型技术,培养基中添加适量抑菌剂,可在自然开放的有菌环境中进行操作[115]。与传统组织培养相比,其从根本上简化了操作环节,成本也极度降低、可大规模生产试管苗。关于开放式培养的研究多集中于抑菌剂的挑选上,这是因为整个操作过程不在无菌的条件下进行,污染便成了关键问题。崔刚等[116]运用中医理论,从多种植物中提取了具有广谱性杀菌、抗菌的物质,并成功地建立了葡萄开放式组织培养体系。目前荸荠[117]、香蕉[118]、魔芋[119]、白菜[120]等植物均已成功建立了开放式组织培养技术体系,关于牡丹的相关技术则尚未见报道。开放式组织培养大力推动了组培苗工厂化发展,为今后研究提供了新思维。
开放式光自养微繁技术,就是将两种新兴技术结合在一起的方法,即在开放有菌的环境下,将外植体接种在含有合适抑菌剂,并且无糖的容器中。这一技术将两个新兴技术完美地融合在一起,既简化了消毒过程,同时使组培室环境得到改善。但开放式光自养培养对植物生长、发育的影响研究还比较有限详细,尚未见成熟技术体系的报道。
4 展望
关于牡丹组织快繁的研究一直是牡丹研究人员关注的重点,这对于保存优良牡丹品种,缩短育种年限以及提高存活率有极大的帮助。虽然还没有呈现出一个完整的技术体系,但已经有了较成熟的基础。比如关于牡丹组织培养过程中常出现的褐化、玻璃化、生根困难等问题均已得到一定程度的解决,研究人员的关注点也不再局限于基础的激素配比上,更多的集中于体细胞培养以及分生结节诱导方面,进而从根本上解决牡丹愈伤分化能力的问题。
随着组织培养技术的快速发展,我国牡丹组织培养技术的研究应在原有的基础上放宽视野,可进一步针对牡丹组织培养进行新技术的研究,从而加快工厂化的推进,或与转基因技术和分子技术相结合,从而致力于与抗性、花期、花色等优良性状相关的牡丹品种的研究。另外,牡丹的药用价值也是关注重点,可通过多学科技术的结合,深层次地掌握其代谢途径。
猜你喜欢
水利水电快报(2024年2期)2024-03-04 03:12:38
果树资源学报(2023年3期)2023-05-11 13:13:12
园艺与种苗(2021年8期)2021-09-23 03:55:30
绿色科技(2021年11期)2021-07-01 13:59:48
高师理科学刊(2020年1期)2020-11-26 05:59:14
现代园艺(2018年3期)2018-02-10 05:18:20
现代园艺(2017年13期)2018-01-19 02:27:58
山东林业科技(2017年5期)2017-04-05 08:33:08
西南农业学报(2016年6期)2016-04-16 05:12:49
新疆农垦科技(2014年6期)2014-02-28 19:20:13
