
卵巢癌中MPC2的表达及其生物学功能
2018-09-11 08:51
中国肿瘤临床 2018年15期
卵巢癌是女性生殖系统中常见的恶性肿瘤之一,发病率仅次于宫颈癌和宫体癌而位居第三位,但卵巢癌死亡率占各类妇科肿瘤的首位,对女性生命造成严重威胁[1]。目前积极的手术治疗和放化疗等基础治疗使得卵巢癌患者的生存状况较前有所好转,但5年生存率较低[2],所以寻找潜在并有效的治疗靶点迫在眉睫。
丙酮酸是糖类、脂肪和氨基酸代谢联系的枢纽,机体细胞整体代谢状态往往取决于丙酮酸的代谢去向[3]。对于正常细胞,在有氧条件下丙酮酸进入线粒体被代谢,在无氧环境中,丙酮酸在胞质中经糖酵解形成乳酸。但是,肿瘤细胞即使在有氧状态下,葡萄糖生成的丙酮酸依然通过糖酵解途径代谢,这就是著名的Warburg效应[4]。虽然Warburg效应被提出了很多年,但是其中的具体机制尚不清楚。线粒体丙酮酸载体(mitochondirial pyruvate carrier,MPC)是介导丙酮酸穿越线粒体膜的重要蛋白,由MPC1和MPC2组成[5−6]。作为介导丙酮酸进入线粒体的重要蛋白,MPC在肿瘤发生及发展过程中是否存在重要的病理生理作用知之甚少。
本研究首先利用http://kmplot.com/analysis/数据库分析卵巢癌组织中MPC1和MPC2的mRNA表达水平与患者生存预后的关系,结果显示MPC2表达水平与患者的预后显著相关。由于MPC2与卵巢癌的显著相关性,因此着重研究MPC2在卵巢癌中的作用,通过体外细胞实验以及检测临床病例组织标本中MPC2的表达情况,分析MPC2与各临床病理参数以及患者生存预后的关系,并探讨MPC2在卵巢癌细胞中的生物学作用,从而为进一步深入研究MPC2在卵巢癌中的作用提供临床和实验依据。
1 材料与方法
1.1 临床资料
选取天津医科大学肿瘤医院2009年1月至2011年12月137例卵巢上皮源性恶性肿瘤患者手术切除的组织标本临床病理资料。所有病例术前均未接受化疗及放疗,且术后组织标本均经病理确诊为卵巢癌。患者年龄19~83岁,中位年龄59岁;组织学分级为高、中分化腺癌59例,低分化腺癌78例;临床分期早期患者49例,晚期88例;其中出现腹水患者90例,远处转移78例。
1.2 方法
1.2.1 免疫组织化学染色及评分 多克隆抗体MPC2(abcam,ab10391)购自美国Abcam公司,免疫组织化学专用一抗稀释液、辣根过氧化物酶标记通用二抗(PV−6000)及DAB显色试剂盒均购自北京中杉金桥生物技术有限公司。免疫组织化学染色采用SP二步法,抗体稀释液(1:100)稀释一抗MPC2,4℃孵育过夜,室温下通用型二抗孵育1 h,DAB显色2 min。免疫组织化学判断原则:综合考虑阳性细胞数和阳性强度,采用半定量方法,依据阳性细胞比例评分为0分(0)、1分(1%~25%)、2分(26%~50%)、3分(51%~100%),并依据着色强度为分0分(无着色)、1分(淡黄色)、2分(棕黄色)及3分(棕褐色),每张切片于热点区域随机选取5个高倍视野评分,最后依据染色结果并按评分结果将两项分值相乘后所有病例划分为两组,0~3分为低表达组,4~9分为高表达组。
1.2.2 细胞培养及过表达MPC2细胞系的构建 卵巢癌细胞系OVCAR3和SKOV3均用含有10%胎牛血清的培养液DMEM培养,于37℃、5%CO2饱和湿度的恒温培养箱中。过表达MPC2的OVCAR3和SKOV3稳定细胞所需系质粒购自深圳华大基因科技服务有限公司。
1.2.3 Western blot检测MPC2过表达水平 取对数生长期的细胞,RIPA裂解液提取细胞蛋白,用BCA法测量蛋白浓度并统一上样量,SDS−PAGE凝胶电泳90 min,Bio−Rad装置湿转蛋白于PVDF膜上,17 mA转膜过夜,切取含目的条带的PVDF膜用5%脱脂奶粉室温封闭1 h。孵育MPC2抗体(1:2 000)与GAP⁃DH内参抗体(美国Santa Cruz公司,1:5 000)4℃过夜,TBST洗膜10 min×3次,室温孵育二抗(1:10 000)1 h,再用TBST洗膜10 min×3次,ECL显影。采用Im⁃age J软件分析细胞内MPC2的表达量。
1.2.4 细胞增殖实验 取对数生长期细胞,首先用0.05%的胰蛋白酶消化,制备成单细胞悬液,细胞计数,以每孔0.2 mL(细胞浓度1×104/mL)细胞接种于96孔板,分别在24、48、72及96 h时间点取出96孔板,每孔加入20 µL的5 mg/mL MTT(3−4,5−dimethyl⁃thiazol− 2− yl)− 2,5− diphenyl− 2H− tetrazolium bro⁃mide),孵育4 h后弃液,再向每孔中加入150µL DMSO,震荡10 min,用酶标仪于490 nm处检测各孔的OD值,绘制生长曲线。
1.2.5 细胞克隆形成实验 制备单细胞悬液,接种于6孔板中,取细胞浓度为1×104/mL 0.1 mL,加入细胞培养液4.9 mL。细胞静置培养10~14 d,当6孔板中出现肉眼可见的克隆时,终止培养。弃去上清液,用PBS清洗3次,加入甲醇原液于−20℃冰箱中固定克隆6 min,0.1%结晶紫溶液室温染色30 min。在光学显微镜(×40倍)下计数>50个细胞的克隆数,并统计克隆形成数量。
1.2.6 Transwell实验 取对数生长期细胞,制备单细胞悬液,离心后用不含血清的培养液重悬细胞计数,在小室中加入取1 mL细胞浓度为1×104/mL细胞液,在24孔板中加入500µL含10%胎牛血清的培养液,将小室放在24孔板中;培养24 h后,取出小室用棉签擦拭小室内部,结晶紫染色后在显微镜下采集图像,计数细胞,进行侵袭迁移细胞数量统计分析。
1.3 统计学分析
采用SPSS 16.0软件进行统计分析,MPC2表达水平与患者临床病理指标之间的相关性分析用χ2检验;生存分析采用Kaplan−Meier法,单因素分析采用Log−rank检验,多因素分析采用Cox比例风险模型。P<0.05为差异具有统计学意义。
2 结果
2.1 MPC1和MPC2 mRNA对肿瘤患者生存预后的影响
首先利用http://kmplot.com/analysis/网站中的数据,分析MPC1和MPC2的基因表达水平对卵巢癌患者总生存期和无进展生存预后的影响。结果发现,MPC1 mRNA的水平表达对卵巢癌患者的无进展生存期(progression−free survival,PFS)(图1A)和总生存率(overall survival,OS)(图1B)的影响差异均无统计学意义;但是,MPC2 mRNA低表达的患者不论是PFS(图1C)还是OS(图1D)均较差。
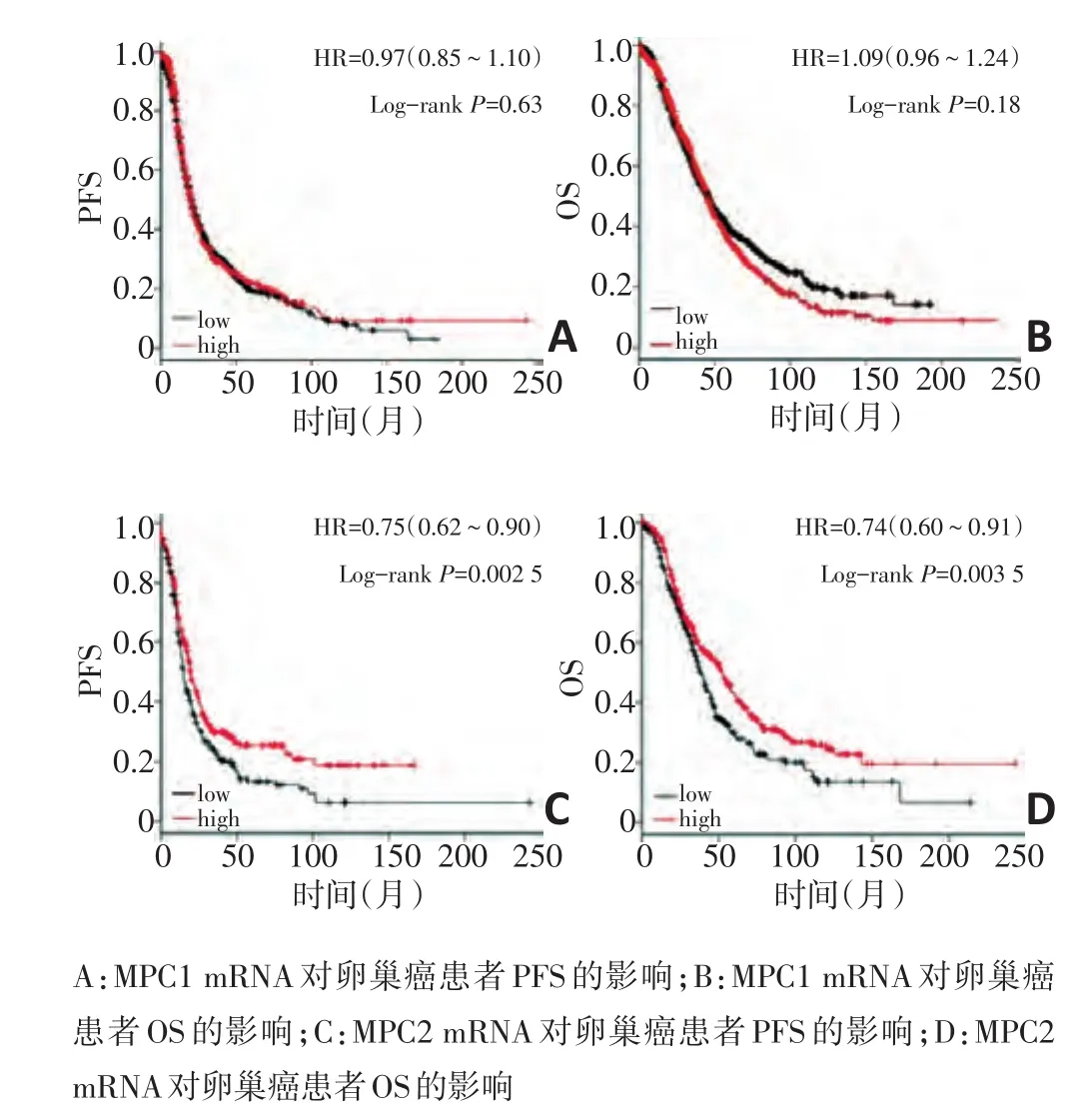
图1 MPC1和MPC2 mRNA对患者生存预后的影响
2.2 MPC2在卵巢癌组织中的表达及其与患者临床病理指标之间的关系
基于前期数据库分析的结果,MPC1对卵巢癌患者的生存影响不显著,本研究主要关注MPC2在卵巢癌中的作用。通过免疫组织化学染色方法检测MPC2在137例卵巢癌组织中的表达水平,结果显示MPC2主要表达于肿瘤细胞的细胞质内,而癌旁正常的卵巢间质细胞及表面上皮内则少有表达(图2A),137例卵巢癌组织中MPC2高表达45例(32.8%)、低表达92例(67.2%)。分析患者年龄、分化程度、临床分期、远处转移及腹水等临床病理指标与MPC2表达水平的关系,发现MPC2的表达水平与腹水的产生(χ2=4.54,P=0.037)、远处转移(χ2=8.31,P=0.004)及临床分期(χ2=17.13,P<0.001)有关,MPC2低表达的患者临床分期较高(图2B)、但MPC2的表达水平与患者的病理分级无关(图2C),MPC2低表达的患者易伴有腹水(图2D)和远处转移(图2E)。且MPC2的表达水平与患者年龄无显著相关性(表1)。
2.3 MPC2表达水平与卵巢癌患者生存预后的关系
采用Kaplan−Meier法对137例卵巢癌病例行统计学分析,单因素生存分析显示,临床分期为早期的患者总生存率显著优于晚期患者(P=0.04,图3)。单因素分析MPC2表达水平与患者生存预后的关系结果显示,MPC2低表达的患者总体预后较差(P=0.03)。Cox比例风险模型多因素分析结果显示,MPC2低表达是卵巢癌患者不良预后的独立风险因素(HR=2.40,P=0.01,表2)。
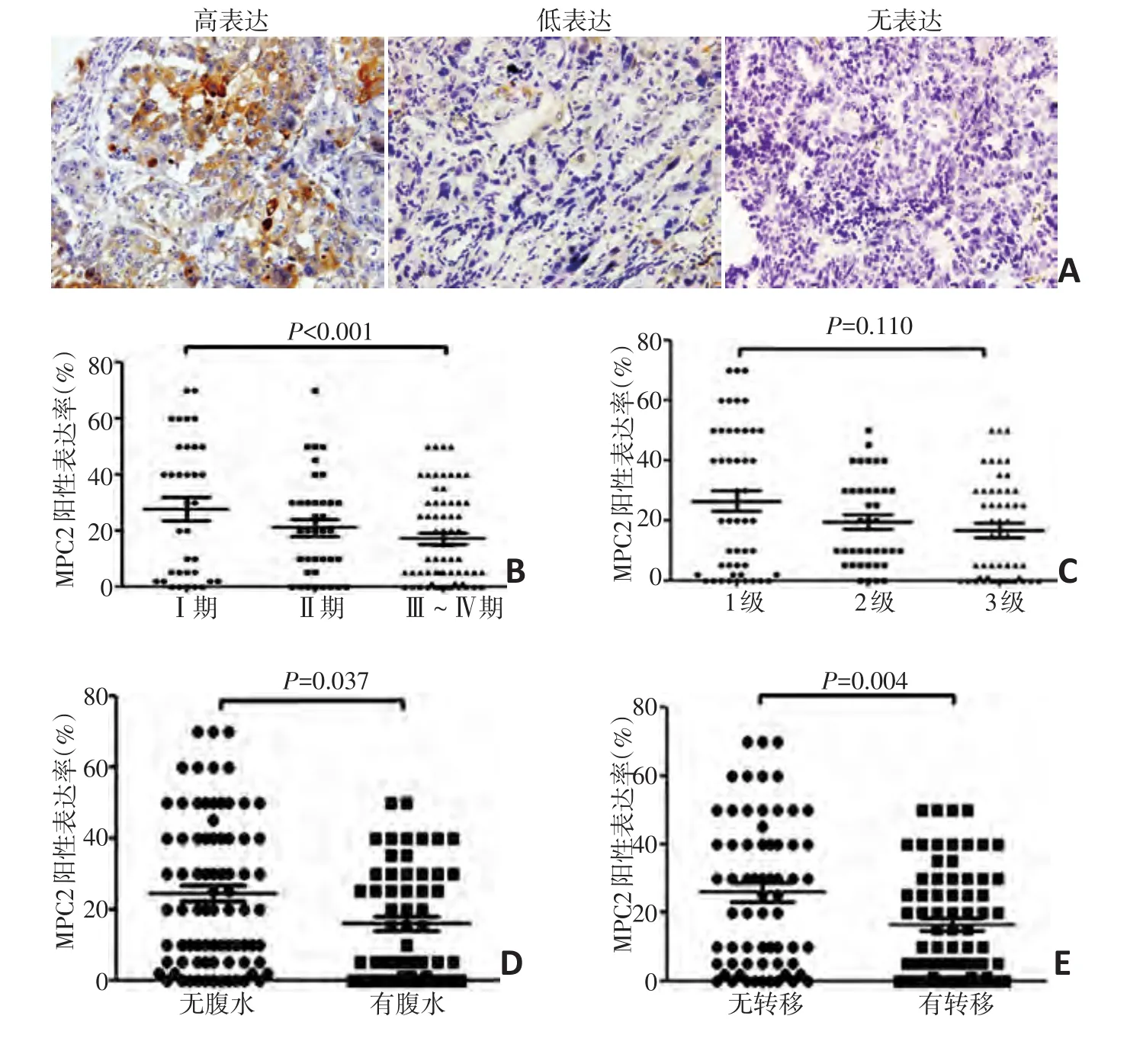
图2 MPC2在卵巢癌组织中的表达及其与临床病理指标之间的关系
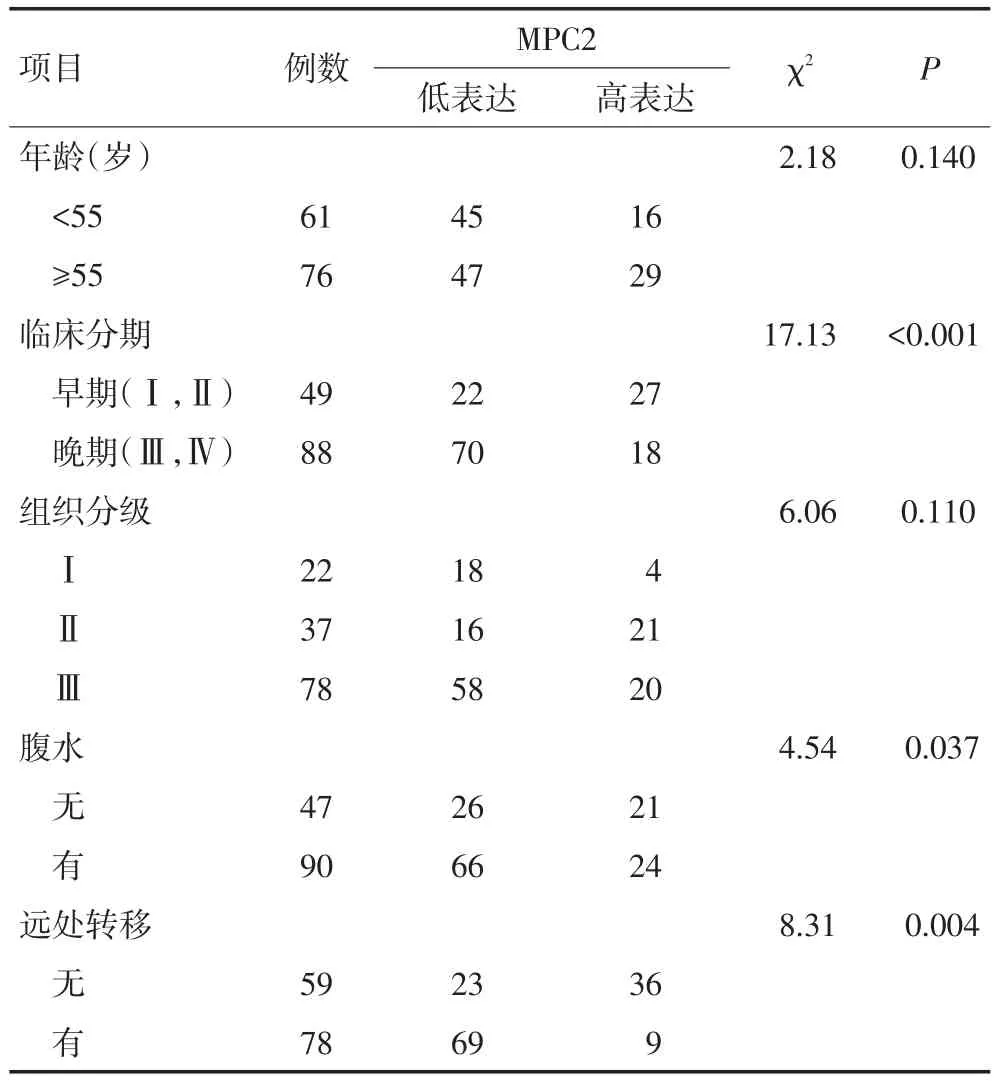
表1 MPC2的表达水平与患者临床病理参数之间的关系
2.4 过表达MPC2对卵巢癌细胞增殖和迁移的影响
在卵巢癌细胞系OVCAR3和SKOV3中转染过表达质粒MPC2,并筛选过表达MPC2的稳定细胞系(图4A)。应用MTT法分别检测MPC2过表达细胞和对照组细胞的生长情况。结果显示过表达MPC2的两种卵巢癌细胞在第3天和第4天的OD值均低于对照组细胞(P<0.05,图4B)。通过二维平板克隆形成实验及Transwell侵袭迁移实验分别检测MPC2的表达对肿瘤细胞克隆形成及迁移能力的影响,结果显示MPC2过表达可以显著抑制细胞的克隆形成,且MPC2过表达可以显著抑制细胞的侵袭迁移能力(图4C)。组间差异具有统计学意义(P<0.05,图4D)。
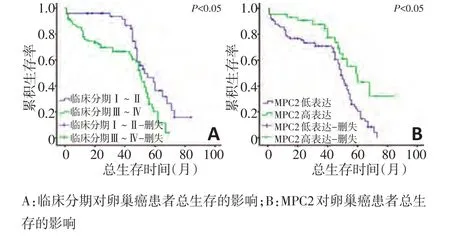
图3 临床分期及MPC2对患者生存预后的影响
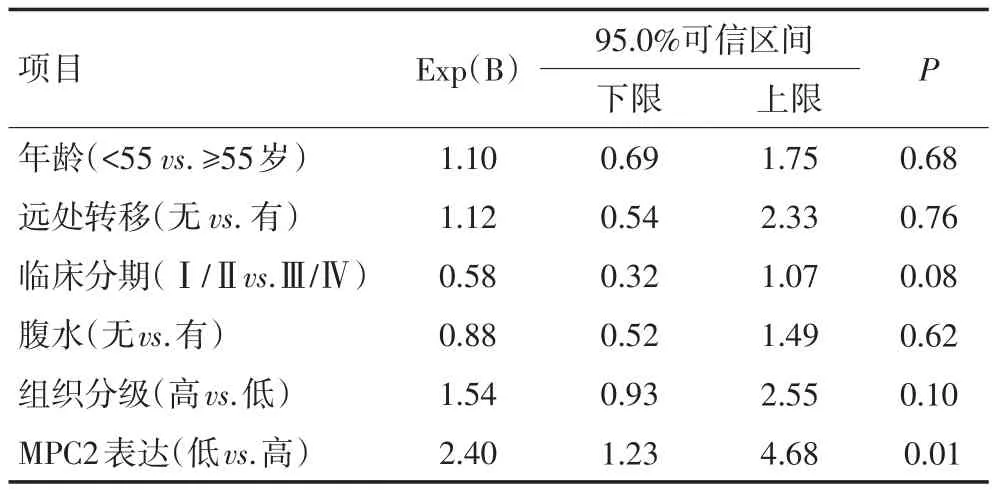
表2 多因素分析各指标对137例卵巢癌患者预后的影响
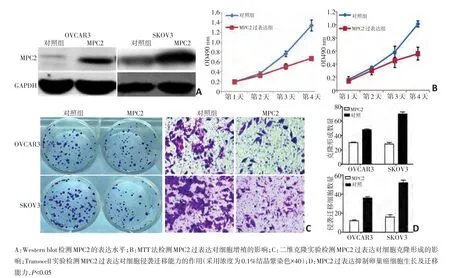
图4 过表达MPC2对卵巢癌细胞生物学行为的影响
3 讨论
上皮性卵巢癌是女性生殖系统常见的恶性肿瘤,世界范围内卵巢癌每年新发病人数与死亡人数均较高[7−8]。多数患者虽然经外科手术有效切除肿瘤,但因出现化疗药物耐药以及肿瘤的复发和转移,致使卵巢癌复发率高和预后不良[9−10]。因此寻找有效的治疗卵巢癌的潜在靶点以及监测预后的标记物迫在眉睫。
失控性增殖是肿瘤的基本特征,肿瘤细胞为了满足自身增殖的需要,经常会出现代谢的重组[11]。肿瘤细胞即使在氧气充足时,依然优先通过糖酵解途径产能,这种供能方式即“Warburg效应”[12]。这种特有的能量获取方式已在多种肿瘤细胞中得到验证,成为肿瘤的重要特征之一[13]。即便经过多年的研究,Warburg效应的发生机制一直未有合理的解释。为什么肿瘤细胞在有氧状态下依然将丙酮酸转化为乳酸,而不是转运到线粒体进行有氧代谢?丙酮酸进入线粒体需要线粒体丙酮酸转运载体蛋白MPC的参与[14],可见MPC在肿瘤细胞代谢重组过程中可能发挥着重要作用。MPC是由MPC1和MPC2组成共同组成的蛋白复合体,目前关于MPC在肿瘤发生及发展过程中的作用研究较少。MPC蛋白在恶性肿瘤中的作用不一致,一项中枢神经系统胶质瘤的研究中发现伴有IDH1突变的胶质瘤病例中MPC1高表达的患者预后明显好于MPC1低表达者,相反MPC2蛋白高表达者生存率低于MPC2低表达患者[15]。由此可见MPC1和MPC2在同一类型肿瘤中所起作用并不一致。本研究首先利用http://kmplot.com/analysis/数据库分析卵巢癌组织中MPC1和MPC2 mRNA表达水平与患者生存预后的关系,结果显示MPC1表达水平对患者生存预后的影响差异无统计学意义,但MPC2表达水平与患者的预后显著相关,MPC2高表达的患者其生存率明显高于低表达。由此可见MPC家族成员在卵巢癌与胶质瘤中的研究结论并不一致,提示MPC1和MPC2在不同组织起源的肿瘤中所起作用不同。本研究重点探讨MPC2在上皮源性卵巢恶性肿瘤中的作用。通过免疫组织化学染色方法检测发现MPC2在卵巢癌组织中的表达水平与肿瘤的恶性进展密切相关,MPC2低表达的患者临床分期较高、易产生腹水和远处转移;并且MPC2低表达是卵巢癌患者不良预后的独立预测因子。通过对137例卵巢癌患者的预后分析,单因素生存分析发现MPC2低表达的患者预后较差,并且多因素生存分析结果也表明MPC2是卵巢癌患者独立的预后因素。MPC2在卵巢癌中的作用与MPC1在低级别神经胶质肿瘤中的作用类似,MPC2表达比率高提示患者可能生存率高,这就提示MPC2有望成为未来监测卵巢癌患者预后的影响因子之一。但MPC如何调控肿瘤细胞的代谢重塑过程,且不同活性的MPC蛋白在不同细胞种类中如何发挥其调控作用仍需进一步研究证实[16]。
已有报道[14]MPC1基因在结肠癌组织中的表达低于相对应的正常组织,MPC2在结肠癌组织中低表达,且与不良预后相关。本研究经免疫组织化学染色方法检测发现MPC2蛋白在卵巢癌组织中的表达均比正常卵巢间质细胞及表面上皮高,提示MPC2在肿瘤发生发展中可能存在细胞代谢的重组调控,致使MPC2表达较正常组织有所升高。肿瘤细胞中能量代谢重塑是使肿瘤进展及细胞转化的关键因素,已有研究报道[6]证实重新过表达MPC可以导致癌细胞的干性特征减弱,这就进一步证明了MPC活性降低或缺失是癌细胞发挥其致癌作用所必须的关键步骤。有研究结论已证实结肠癌组织中MPC蛋白表达下降,且MPC低表达和不良预后有关[17]。另外一项前列腺癌的研究报道也提出MPC1和MPC2阳性表达是良好预后的标记之一[18]。这些研究结论与本研究中发现MPC2的低表达与卵巢癌的恶性进展密切相关结论相同。正常机体细胞主要以氧化磷酸化的方式供能,源于这种产能方式所提供的能量比糖酵解途径要多,为何肿瘤细胞通过糖酵解的方式代谢是当前研究细胞能量代谢的重点领域,MPC将丙酮酸运载至线粒体起核心作用,为研究MPC对卵巢癌细胞能量代谢及生物学功能的影响作用,本实验利用体外细胞实验检测卵巢癌细胞系中增加MPC2的表达后对细胞生长及运动能力的影响,结果显示MPC2可以显著抑制细胞的增殖和迁移能力。MPC2低表达在卵巢癌中是如何促进肿瘤恶性进展及其对肿瘤细胞能量代谢转化并重塑的具体分子机制,尚需进一步深入研究证实。本研究结论提示MPC2在卵巢癌恶性进展过程中可能起着重要作用,而且MPC2低表达可以作为卵巢癌患者不良预后的独立预后指标,MPC2可抑制卵巢癌细胞的生长及侵袭迁移能力,MPC2在包括卵巢癌在内的多种恶性肿瘤的发生发展中起着重要作用,且MPC2低表达具有一定的普遍性,逆转MPC2表达可影响肿瘤细胞能量代谢转化,MPC2有可能成为未来肿瘤生物治疗中的潜在靶点。
猜你喜欢
山西医科大学学报(2022年6期)2022-08-04
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
牡丹江医学院学报(2021年5期)2021-12-05
天津医科大学学报(2021年3期)2021-07-21
河南化工(2020年2期)2020-04-20
食品安全导刊(2018年30期)2019-01-28
中成药(2017年8期)2017-11-22
CHINESE JOURNAL OF AERONAUTICS(2016年3期)2016-11-23
医学研究杂志(2015年7期)2015-06-22
