
发酵豆粕对生长猪生长性能、血清生化指标和抗氧化性能的影响
2018-08-24 09:10:00蔡爱萍
中国饲料 2018年14期
蔡爱萍
(河北省保定市阜平县农业局,河北保定 073200)
豆粕是动物生产中重要的蛋白来源,但豆粕中含有的抗营养因子限制了动物机体对养分的吸收利用(袁林,2012)。近年来,利用微生物发酵技术降解豆粕中的抗营养因子成为主流,发酵豆粕可使豆粕中的抗营养因子降低,大分子蛋白转化为易消化吸收小肽,伴随发酵过程还会产生维生素、消化酶和未知生长因子(宋文心和邵庆均,2009)。随着我国对猪肉安全问题的重视,无抗养殖成为趋势,微生态产品也将逐渐替代抗生素。在猪日粮中添加发酵豆粕可改善猪的肠道健康,增强抵抗力,提高养殖效率。目前,发酵豆粕的研究多集中在仔猪阶段,对生长速度快、发病率高的生长猪阶段研究较少。本试验以生长猪为试验动物,研究发酵豆粕对生长猪生长性能、血清生化指标和抗氧化性能的影响,意在为生长猪日粮中发酵豆粕添加量提供参考依据。
1 材料与方法
1.1 试验饲料 试验用发酵豆粕由日照东维饲料有限公司提供,粗蛋白质≥48%、酸溶蛋白质≥10%、乳酸≥3.5%。试验日粮为玉米-豆粕型生长猪配合饲料,营养水平参照猪NRC(2012)营养需要量标准(见表1)。
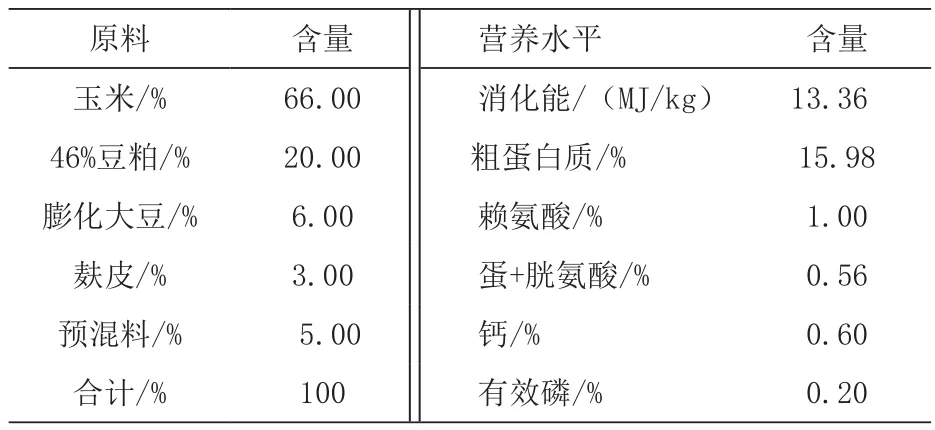
表1 基础饲料组成及营养水平
1.2 试验动物及设计 试验用生长猪由新希望六和集团提供,在山东新希望集团岭泉猪场进行养殖试验。将300头杜长大外三元生长猪,按照健康状况一致,公母比例相同,体重相近的原则随机分为5组,每组6个重复,每个重复10头猪,采用圈养方式,每圈为一个重复。Ⅰ组为空白对照组,饲喂不添加发酵豆粕的基础日粮,Ⅱ~Ⅴ组分别在基础日粮中添加2%、4%、6%、8%的发酵豆粕替代46%豆粕,试验期为35 d。
1.3 饲养管理 试验开始前对猪舍、设备彻底清洗消毒,空栏一周。正式试验前各试验组先以生长猪前期料与试验用基础日粮按比例混合过渡预饲7 d,其他基础条件相同。正式开始后,生长猪的平均重量为(30.55±0.52)g,试验猪全期自由采食、饮水,少喂勤添,定期清理粪便。每天记录温度、病死数、采食量等数据,注意观察生长猪的健康状态。
1.4 测定指标及方法 试验开始日、试验结束日早上8:00分别以重复为单位进行空腹称重,称重前不停水,停料6 h。对生长指标进行统计记录,得出试验周期猪只初始重、期末重、平均日增重(ADG)、平均日采食量(ADFI)、料重比(Feed/Gain)、腹泻率等数据。
试验期最后一天对试验猪采血。每重复随机选择2头健康状况良好的猪只(一公一母),早上8:00空腹颈静脉采血10 mL,采集血样放于10 mL离心管中,于4℃冰箱静置3 h,3000 r/min离心后取上层血清,每圈作为一个样品,置于-20℃冰箱保存备用。根据试验设计测定指标,将样品分为两部分,分别用于血清生化指标和抗氧化指标。
黄嘌呤氧化酶法测定血清总超氧化物歧化酶(T-SOD)活力,比色法测定谷胱甘肽过氧化物酶(GSH-Px)活力,紫外法测定过氧化氢酶(CAT)活力;分别采用TBA法、FRAP法测定血清丙二醛(MDA)含量和总抗氧化能力(T-AOC),模拟机体黄嘌呤氧化酶与黄嘌呤反应系统的方法测定血清中抗超氧阴离子活力(ASA)。葡萄糖氧化酶-过氧化物酶法测定血清葡萄糖(GLU)含量;考马斯亮蓝法测定总蛋白(TP)含量;脲酶法测定尿素氮(UN)含量;酶偶联比色法测定总胆固醇(TCH)和甘油三酯(TG)含量;AMP缓冲液法测定碱性磷酸酶(ALP)活性。试验用试剂盒均为南京建成生物工程研究所提供。
1.5 数据处理与统计分析 以重复为单位对试验所得数据进行处理,应用SPSS 19.0软件中单因素方差分析(One-way ANOVA)中的LSD法进行,用Duncan’s法进行多重比较。以“平均值±标准差”形式表示试验数据结果。P<0.05和P<0.01分别表示为差异显著和差异极显著。
2 结果与分析
2.1 发酵豆粕对生长猪生长性能的影响 由表2可知,生长猪期末重,与对照组相比,Ⅳ、Ⅴ组显著增加(P<0.05),Ⅳ、Ⅴ组间差异不显著(P>0.05);平均日增重,Ⅳ、Ⅴ组极显著高于对照组(P<0.01),Ⅱ、Ⅲ组显著高于对照组(P< 0.05),Ⅳ、Ⅴ组间差异不显著(P> 0.05),Ⅱ、Ⅲ组间差异不显著(P>0.05)。
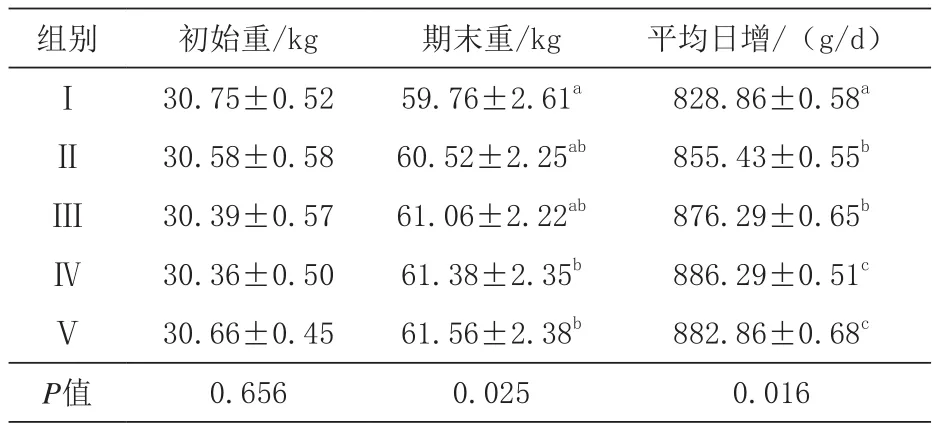
表2 初始重、期末重、平均日增重
由表3可知,Ⅳ、Ⅴ组平均日采食量显著高于对照组(P<0.05),Ⅳ、Ⅴ组间差异不显著(P>0.05);Ⅲ、Ⅳ、Ⅴ组料重比显著低于对照组(P<0.05),且各添加发酵豆粕组间差异不显著(P>0.05);腹泻率各组间差异不显著(P>0.05),但随发酵豆粕添加比例增大而呈下降趋势。
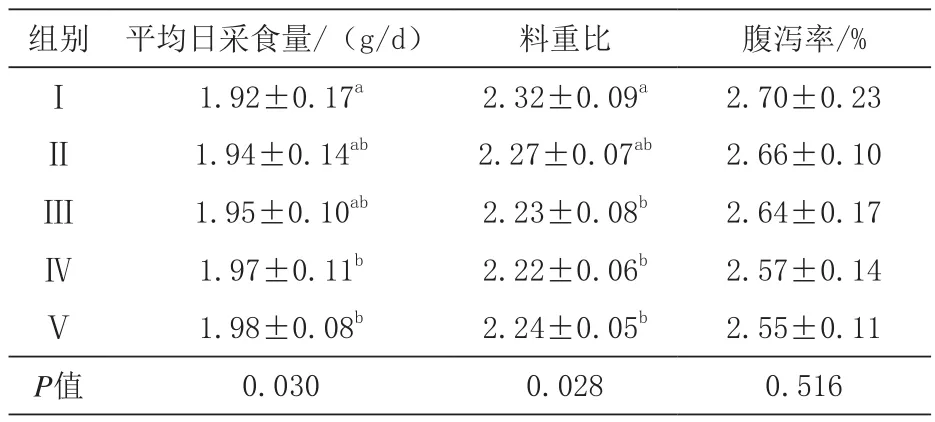
表3 平均日采食量、料重比、腹泻率
2.2 发酵豆粕对生长猪血清生化指标的影响由表4可知,与对照组相比,血清葡萄糖含量Ⅳ组显著降低(P<0.05),Ⅴ组极显著降低(P<0.01),Ⅲ、Ⅳ、Ⅴ组间差异不显著(P>0.05);Ⅳ组血清总蛋白含量极显著高于对照组组(P<0.01),Ⅳ、Ⅴ组间差异不显著(P>0.05);与对照组相比,Ⅳ、Ⅴ组血清尿素氮含量显著降低(P<0.05),且各添加发酵豆粕组间差异不显著(P > 0.05)。
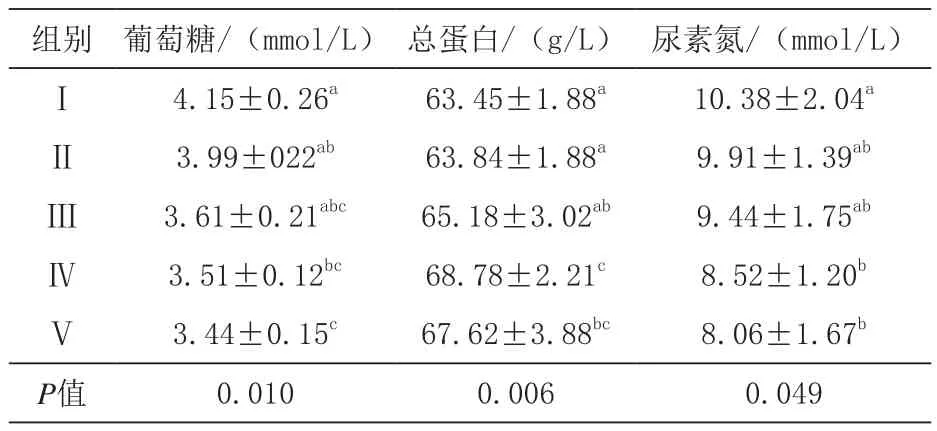
表4 葡萄糖、总蛋白、尿素氮含量
由表5可知,血清总胆固醇含量,Ⅳ、Ⅴ组显著低于对照组(P<0.05),各添加组间差异不显著(P>0.05);与对照组相比,Ⅴ组血清甘油三酯含量显著降低(P<0.05),各组间差异不显著(P>0.05);与对照组相比,Ⅳ、Ⅴ组血清碱性磷酸酶活性显著提高(P<0.05),且各添加组间差异不显著(P>0.05)。
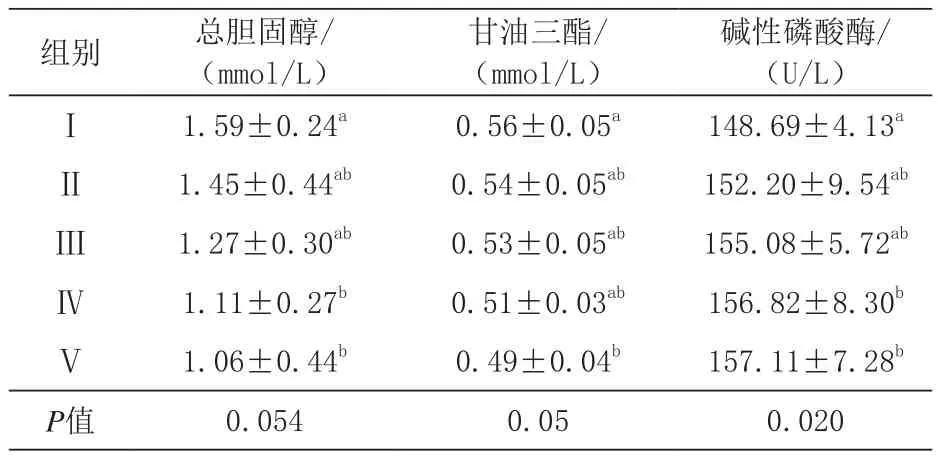
表5 总胆固醇、甘油三酯、碱性磷酸酶活性
2.3 发酵豆粕对生长猪抗氧化性能的影响 由表6可知,各组间血清T-SOD活性差异不显著(P>0.05),但随发酵豆粕添加比例增大而呈增加趋势;Ⅳ、Ⅴ组血清GSH-Px活性显著高于对照组(P<0.05),Ⅳ、Ⅴ组间差异不显著(P>0.05);,各组间血清CAT活性差异不显著(P>0.05),且与发酵豆粕添加比例无相关性。
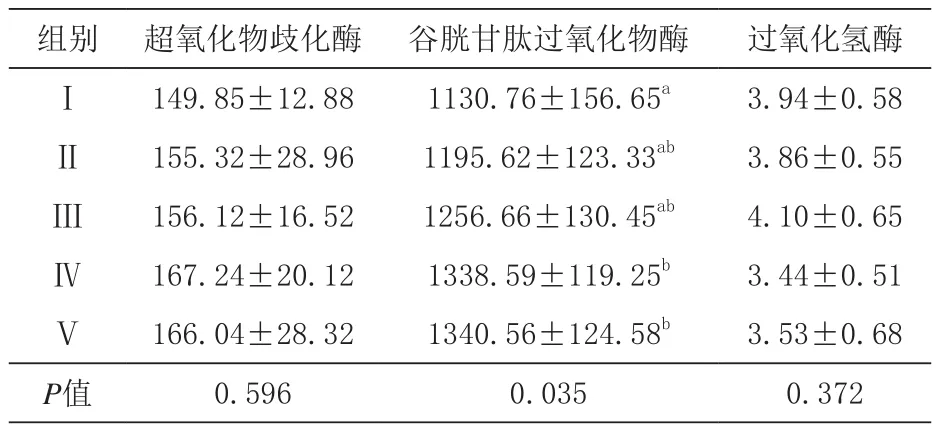
表6 超氧化物歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶活性 U/mL
由表7可知,与对照组相比,Ⅳ、Ⅴ组血清抗超氧阴离子活力显著提高(P<0.05),Ⅳ、Ⅴ组间差异不显著(P>0.05);,与对照组相比,Ⅳ、Ⅴ组血清丙二醛含量显著降低(P<0.05),Ⅳ、Ⅴ组间差异不显著(P>0.05);Ⅳ、Ⅴ组血清总抗氧化能力极显著高于对照组(P<0.01),Ⅱ、Ⅲ组显著高于对照组(P<0.05),且Ⅳ、Ⅴ组间差异不显著,Ⅱ、Ⅲ组间差异不显著(P>0.05)。
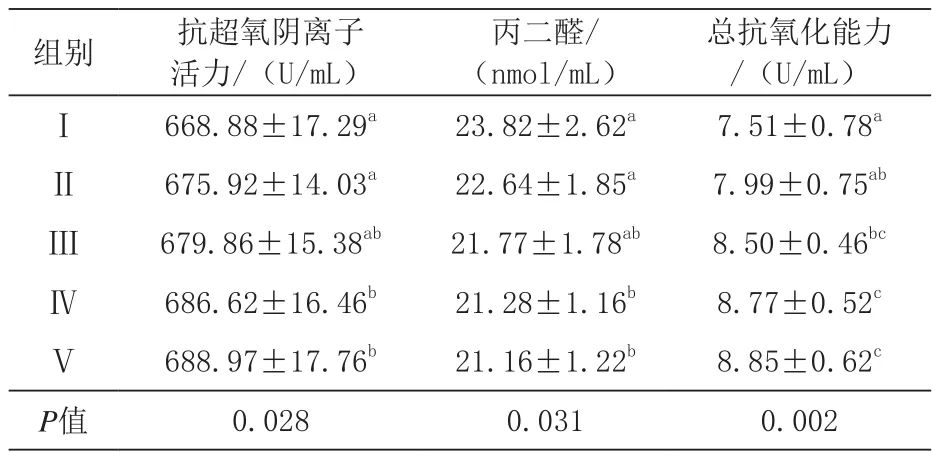
表7 抗超氧阴离子活力、丙二醛含量、总抗氧化能力
3 讨论
3.1 发酵豆粕对生长猪生长性能的影响 蛋白质作为动物机体重要组成部分,可以修补、更新细胞组织,蛋白质利用率的高低直接影响畜禽生产性能(马静等,2007)。经微生物发酵处理后的豆粕富含小分子肽、游离氨基酸、未知生长因子、消化酶和维生素,可提高机体对营养物质的消化利用率(刘海燕等,2010)。通过发酵工艺益生菌将豆粕中的不良气味去除,使发酵豆粕具有特殊酸香风味,提高日粮适口性,增加采食量(Mital等,2010)。豆粕发酵过程中产生的乳酸能改善肠道环境,适宜益生菌生长繁殖,有机酸还能提高消化酶活性,增强肠道蠕动,从而提高肠道消化吸收能力(李少宁等,2016)。徐述亮等(2013)在仔猪日粮中添加4.3%的发酵豆粕,与对照组相比,平均日增重提高12.32%(P<0.01),平均日采食量提高8.17%(P<0.05),试验期末重提高3.81%(P< 0.05),料重比降低 3.79%(P< 0.01),腹泻率降低33.33%(P<0.05)。章世元等(2009)在断奶仔猪日粮中添加35%和17.5%的发酵豆粕,与对照组比,料重比分别降低9.47%和4.21%。郑云峰等(2006)在40日龄仔猪日粮中添加8%的发酵豆粕代替膨化大豆和鱼粉,与对照组相比,试验组腹泻率显著降低(P<0.05),经济效益得到提高。本研究中,与对照组相比,6%添加组可极显著提高生长猪结算的平均日增重(P<0.01),显著增加生长猪的平均日采食量(P<0.05),显著降低生长猪的料重比(P<0.05),腹泻率各组间差异不显著(P>0.05),但随日粮发酵豆粕添加水平的增加有降低趋势。因此,生长猪日粮中添加发酵豆粕6%可去除豆粕中的有害因子,促进猪肠道的消化吸收,提高经济效益,提升生长性能。
3.2 发酵豆粕对生长猪血清生化指标的影响动物血清中的葡萄糖分为肠道吸收、糖原分解和肝脏糖的异生3种主要来源,为机体提供能源(孙淑洁等,2012)。发酵豆粕中小分子活性肽能显著提高动物血液中胰岛素水平,促进血糖分解吸收,提高畜禽对糖类的利用率(张琳琰等,2012)。总蛋白由白蛋白(ALB)和球蛋白(GLB)组成,是机体细胞的重要组成部分,在一定限度内,总蛋白浓度与机体合成利用蛋白质能力正相关(孟苓凤等,2013)。蛋白质进入机体经过一系列代谢后形成血清尿素氮,在正常生理状态下,体内氮沉积量与血清尿素氮含量呈负相关,氮沉积水平的高低直接影响机体蛋白质合成量(徐涛等,2013)。李少宁等(2015)在生长猪日粮中添加3%的发酵豆粕,与对照组相比,总氮、粪氮、尿氮分别降低了19.6%、1.07%、22.24%,氮沉积率提高53.83%,氮生物学价值提高12.26%,表明发酵豆粕可提高生长猪蛋白利用率,降低氮排泄量,减少环境污染。章世元等(2008)在猪日粮中添加17.5%和35%的发酵豆粕,与对照组相比,显著提高猪对粗脂肪的表观消化率,从而促进脂肪代谢,这与发酵豆粕促进小肠绒毛发育和增加胃黏膜厚度相关。碱性磷酸酶与磷代谢密切相关,重要的解毒因子有非特异性免疫作用(郭航等,2008)。本研究中,与对照组相比,添加发酵豆粕组均能显著改善生长猪的糖、蛋白质、脂肪代谢功能,6%添加组生长猪血清葡萄糖含量显著降低(P<0.05),血清尿素氮含量显著降低性(P<0.05),血清总蛋白含量显著增加(P<0.05),血清总胆固醇含量显著降低(P<0.05),血清碱性磷酸酶活性显著增加(P<0.05)。因此,生长猪日粮中添加6%的发酵豆粕可提高机体对糖、脂肪、蛋白质、磷等的利用率,显著改善其血清生化指标。
3.3 发酵豆粕对生长猪抗氧化性能的影响 机体内部和外界化学物质代谢过程中产生了活性氧和自由基,引起氧化损伤导致各种疾病,机体抗氧化性能反应了其健康状况与抗外界环境损伤的能力(Giannenas,2015)。发酵豆粕中富含益生菌,使氨基酸容易被畜禽消化吸收,动物机体对氨基酸的获得跟组成吻合,保证氨基酸的平衡,从而提高机体抗氧化能力(梁明振等,2002)。发酵过程中产生的有机酸能清除机体自由基,螯合重金属,增强还原能力(Gibbs等,2004)。经过益生菌发酵后,豆粕异黄酮结构和分子量均发生显著变化,异黄酮糖苷转化为异黄酮戒元,抗氧化功能均得到显著提高,但抗氧化功能主要是由抗氧化肽作用产生的,大豆中的生物活性物质经发酵后释放出来形成抗氧化肽,天然抗氧化剂稳定性强,能防止脂质过氧化发生,保护细胞和器官的正常结构和功能(Junko等,2010)。机体中抗氧化酶主要为T-SOD、GSH-Px、CAT等,能单独或协同清除体内氧自由基,MDA直接反应机体脂质过氧化程度,间接反映细胞损伤程度,超氧阴离子为机体生成氧自由基的源头(吴先华等,2013)。刘海燕(2012)分别在试验猪日粮中14.27%和28.54%的发酵豆粕,与对照组相比,试验组仔猪血液中MDA含量分别降低了24.89%和27.97%,SOD和T-AOC含量随添加比例的提高呈升高趋势,该研究结果与本试验结果相似。本研究中,与对照组相比,6%添加组生长猪血清GSH-Px活性显著提高(P<0.05),血清T-AOC极显著增高(P< 0.01),血清 ASA 显著提高(P< 0.05),血清MDA含量显著降低(P<0.05)。各添加组间血清SOD活性差异不显著(P>0.05),但随添加水平的提高有增加趋势。因此,生长猪日粮中发酵豆粕添加量为6%,可提高机体抗氧化酶活性,去除脂质过氧化物跟自由基,显著改善其抗氧化性能。
4 结论
在本试验条件下,生长猪日粮中添加6%和8%的发酵豆粕均能显著改善生长猪的生长性能、血清生化指标和抗氧化性能,但从经济角度出发,最适添加量为6%发酵豆粕替代普通豆粕。
猜你喜欢
中老年保健(2021年7期)2021-08-22 07:40:46
当代水产(2019年11期)2019-12-23 09:03:54
当代水产(2019年7期)2019-09-03 01:02:18
当代水产(2019年6期)2019-07-25 07:52:10
中学生数理化·七年级数学人教版(2019年6期)2019-06-25 01:01:32
柴油机设计与制造(2018年3期)2018-10-13 01:45:08
中华皮肤科杂志(2018年4期)2018-01-22 05:09:25
China Geology(2018年3期)2018-01-13 03:07:16
河南畜牧兽医(2017年8期)2017-11-24 03:21:13
中国铸造装备与技术(2017年3期)2017-06-21 11:33:37
