
长白山北部区域西伯利亚狍局域种群遗传特征的分析
2018-08-21 02:37盛清宇孙铁铎张崇颖罗理杨姜广顺
野生动物学报 2018年3期
盛清宇 孙铁铎 张崇颖 罗理杨 姜广顺
(1.国家林业局猫科动物研究中心,东北林业大学野生动物资源学院,哈尔滨,150040;2.吉林省林业勘察设计研究院,长春,130021;3.吉林省天桥岭林业局,汪清,133204)
狍(Capreolusspp.)是鹿科(Cervidae)中分布最广泛的动物之一,包括2个种,即体型相对较小的欧洲狍(Capreoluscapreolus)以及体型相对较大的西伯利亚狍(Capreoluspygargus)。西伯利亚狍的主要分布区域包括整个亚洲大陆的古北界[1]及东欧的部分地区[2]。在中国,狍广泛分布于东北区大兴安岭亚区、长白山地亚区、松辽平原亚区、华北区黄淮平原亚区、黄土高原亚区、蒙新区天山山地亚区、青藏区青海藏南亚区,同时存在部分边缘分布于西南区南山地亚区及华中区东部丘陵平原亚区[3]。在东北虎(Pantheratigrisaltaica)、东北豹(Pantherapardusorientalis)分布区域内,狍也有广泛分布,并构成了东北豹的主要猎物(占食物构成的66%)以及东北虎的猎物之一(占食物构成的9%)[4]。因此,研究狍的种群遗传多样性,对于探究猎物种群有效恢复,对东北虎、东北豹种群的保护有着重要的意义。
目前,已经有许多关于欧洲狍种群遗传学和系统发生生物地理学的研究[5-7],这些研究利用线粒体DNA和微卫星作为分子标记,揭示了欧洲狍种群的遗传结构和地理分布差异。而关于西伯利亚狍的种群遗传学研究,不管在国外还是国内都相对较少,且一般是使用线粒体DNA作为分子标记,而较少选择微卫星标记来进行研究[8-10]。
微卫星是由2~5个核苷酸串联的重复序列构成,由于其广泛分布于基因组中,在长度上具有明显的多态性[11],并且是进化最快的DNA序列之一[12-14]。因此,是研究种群结构及亲缘关系的有力工具[15]。目前,有不少利用微卫星标记分析欧洲狍种群遗传学的研究,如Randi等通过微卫星STR数据揭示了在伊比利亚北部和意大利南部的狍种群之间的遗传差异[16];Baker和Hoelzel使用线粒体DNA和微卫星标记的研究方法,比较了英国原有的狍种群和引入种群之间的遗传多样性差异以及种群混合和扩散[17]。关于西伯利亚狍种群遗传学的研究目前相对较少,Lee等对亚洲10个地区西伯利亚狍的12个微卫星位点的变异进行了研究,分析了各个种群的遗传多样性水平,基于贝叶斯的聚类分析揭示了西伯利亚狍存在的3个不同地理种群,分别是东南部种群(包括朝鲜半岛、俄罗斯远东地区、跨拜加尔省和蒙古北部地区)、西北部种群(包括西伯利亚西部和俄罗斯的乌拉尔)以及韩国济州岛种群[18]。总的来说,目前国内利用微卫星标记分析西伯利亚狍种群遗传多样性的报道相对空白。鉴于此,本研究利用10个微卫星位点,结合荧光标记PCR技术,对东北虎、东北豹分布区域内的西伯利亚狍种群进行了局域种群遗传多态性的研究,目的在于为狍种群资源的保存利用以及后续研究狍作为猎物对东北虎、东北豹的影响提供支持。
1 材料与方法
1.1 样品采集
在2014年12月~2017年3月期间,在冬季外业过程中采集了共计329份西伯利亚狍的粪便样品,分布的区域包括黄泥河、珲春、汪清、穆棱、东京城、天桥岭、东宁。样品采集区域见图1。所用粪便样品均在冬季采集,并保存于-20℃的条件下,用于后续实验步骤。
1.2 粪便DNA提取、电泳检测
采用QIA amp DNA Stool Mini Kit试剂盒(QIAGEN,Germany)提取DNA,电泳检测后使用提取效果好的进行PCR扩增,第一次提取后质量不高的样品重新提取DNA。329份样品中,共计287份成功提取DNA,用于后续实验步骤。

图1 研究地区地理位置示意图Fig.1 Location of study area
1.3 微卫星位点及引物选择
参考相关文献[19-22],选取了10个微卫星位点并合成引物,进行荧光染料标记。引物序列信息见表1。
1.4 PCR扩增
采用由新海基因检测有限公司合成的10对引物,分别对DNA样品进行3次以上的PCR重复扩增。PCR反应总体系20 μL:10×PCR Buffer 10 μL,dNTPMixture(2.5 mmol/L)4 μL,上/下游引物(10 mmol/L)各0.75 μL,TOYOBO高保真酶0.3 μL,模板DNA 1.0 μL,ddH2O补足体系。PCR扩增程序:94℃预变性2 min;94℃变性20 s,55~58℃退火1 min,68℃延伸1 min,35 个循环;68℃延伸10 min。扩增完毕后,取5 μL PCR产物在含有核酸染料的1.0%琼脂糖凝胶电泳上进行电泳检测以确定PCR扩增是否成功;将扩增成功的样品送至上海生工生物有限公司,将混合液在ABI3730X1 测序仪上进行测序分型。
1.5 统计分析
利用 FSTAT(v.2.9.3)软件、Cervus软件统计遗传参数,包括等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)、多态信息含量(PIC)、脱离种群数量影响的等位基因丰富度(allele richness,AR)以及各种群固定系数(fixation index)。利用FSTAT (v.2.9.3)软件计算两两种群之间的FST统计量[23],用于表征种群遗传分化的水平。
表1 西伯利亚狍种群10个微卫星位点的相关信息及退火温度

Tab.1 Information and annealing temperature of 10 microsatellite loci from Siberian roe deer
2 结果与分析
2.1 分型结果
利用ABI3730X1 测序仪对每个样品不同微卫星位点的荧光标记 PCR 扩增产物进行微卫星样本分型结果的获取,使用GeneMarker软件对这些微卫星样品的分型结果进行判读,得出了10个微卫星位点的扩增产物电泳信号图,选取部分信号图如图2所示。所显示的信号图表明微卫星DNA扩增所得的条带清晰,电泳信号较强,实验结果是可靠的,可用于下一步数据分析。
2.2 微卫星位点的遗传多样性分析
本研究所使用的10个微卫星位点在西伯利亚狍种群中检测到的等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)、多态信息含量(PIC)以及固定系数(fixation index)结果见表2。10个基因座位的等位基因数为4.1~17.1,平均等位基因数为9.6。其中,等位基因数最多的位点是BM757,有17个等位基因,最少的位点是MCM507,有4个等位基因。所检测到的10个座位中的平均多态信息含量为0.73,多态信息含量最高的位点是BM757(0.93),最低的位点是MCM507(0.33)。
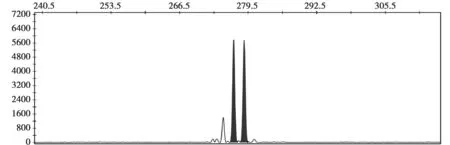
图2 微卫星位点扩増产物电泳信号图Fig.2 Signal figure of microsatellite loci tested by capillary electrophoresis
表2 西伯利亚狍种群10个微卫星位点的遗传多样性参数

Tab.2 Genetic diversity parameters of 10 microsatellite loci from Siberian roe deer population
2.3 种群的遗传多样性分析
本研究所得到的7个西伯利亚狍局域种群的等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)、脱离种群数量影响的等位基因丰富度(allele richness,AR)以及各种群固定系数(fixation index)结果见表3。由表3可知,东北地区西伯利亚狍种群的等位基因数在7.3~11.1之间,平均等位基因数为8.9。等位基因丰富度整体水平较高,为7.5,其中,水平最高的是东宁地区种群的8.4,水平最低的是黄泥河地区种群的6.14。观测杂合度和期望杂合度变化范围在0.7~0.76之间,观测杂合度小于期望杂合度,处于较高水平,并且两者之间未表现出明显的偏离。固定指数的变化范围在-0.01~0.061之间。
表3 东北地区7个西伯利亚狍种群遗传多样性参数
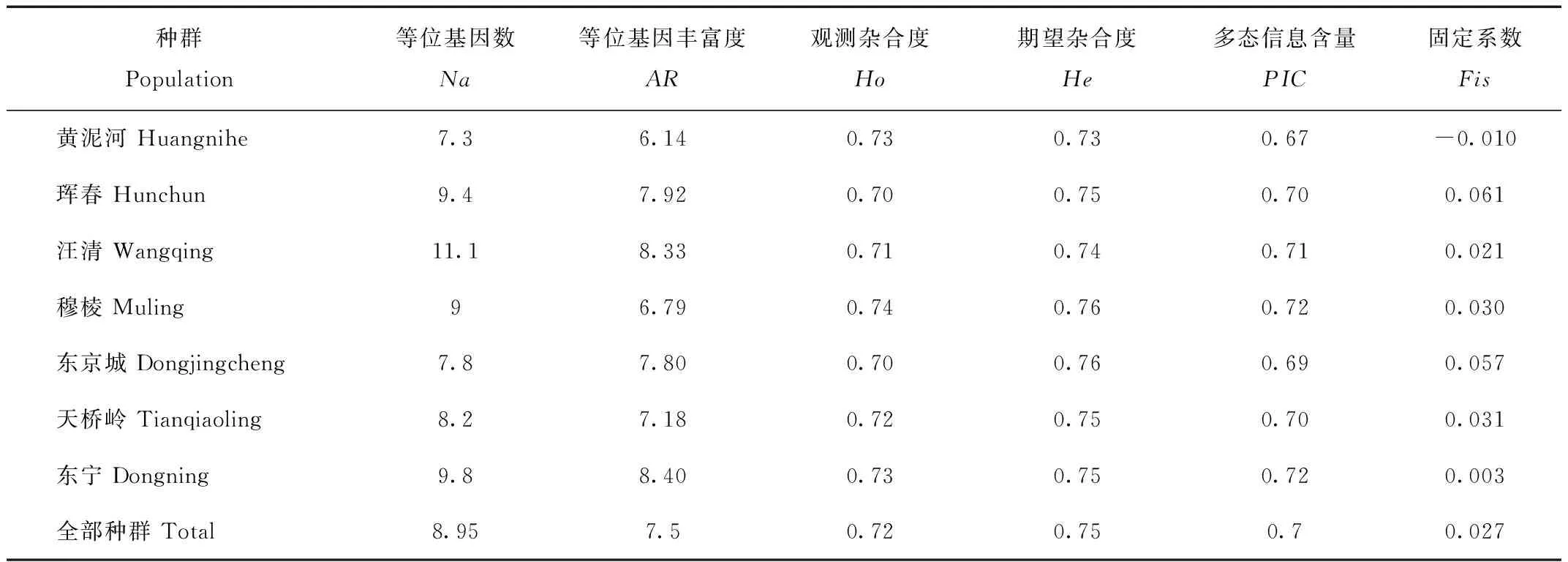
Tab.3 Genetic diversity parameters of 7 Siberian roe deer populations in northeast China
2.4 种群结构
FST评估表明东北地区狍种群之间存在较小至中等程度的遗传分化(表4)。FST值的变化范围在0.002~0.074之间,其中,黄泥河地区的狍种群与其他6个地区的狍种群均存在中等程度的遗传分化,而剩余6个地区的种群之间遗传分化程度相对较低。
表4 种群间遗传分化系数 FST
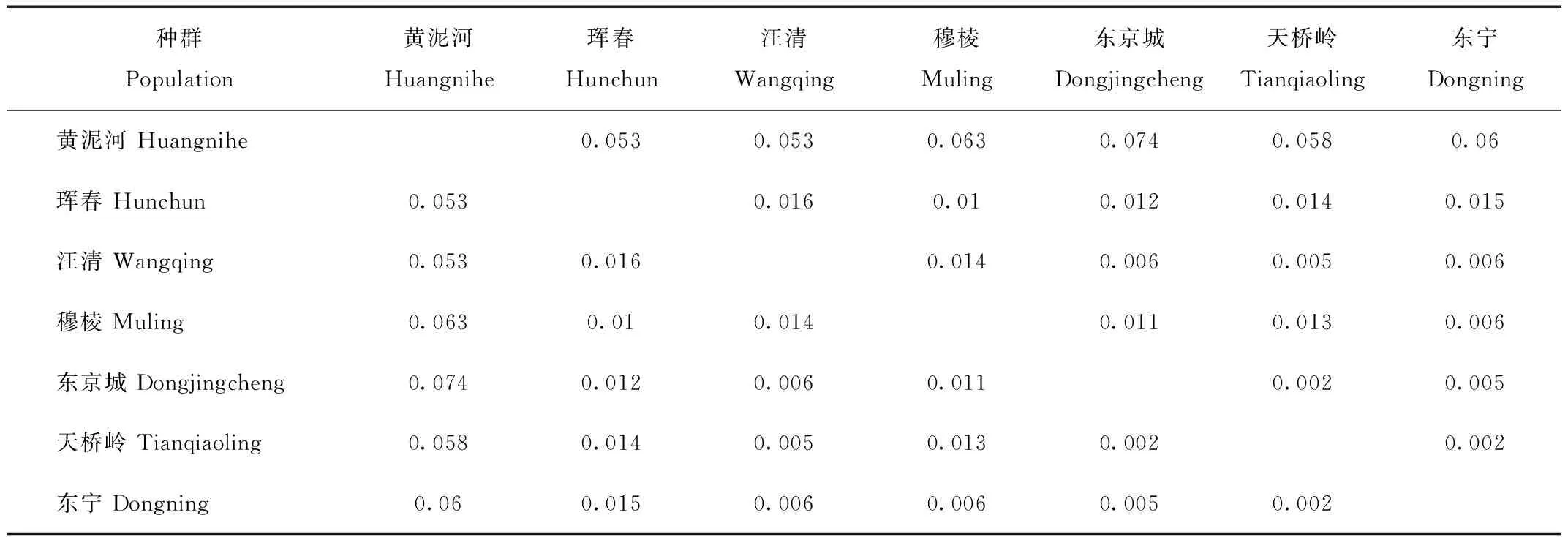
Tab.4 Population comparisons with FST values
3 讨论
多态信息含量(PIC)是用以描述微卫星位点多态性大小的尺度,Botstein等[24]的研究表明,当PIC>0.5时,认为该基因座位为高度多态位点,当0.25 杂合度(H)是衡量种群内遗传变异大小的一项可靠指标,其中,期望杂合度值能够直接反映种群内个体的均匀度,杂合度值越高,则种群内的遗传变异程度越高,杂合度值越低,则种群内的遗传变异程度越低[25]。一般情况下观测杂合度与期望杂合度越接近,表明该种群受外界因素或近交等的影响越小。本研究所得出的东北地区西伯利亚狍种群的平均观测杂合度(0.72)比期望杂合度(0.75)低,7个种群中,除黄泥河地区的狍种群观测杂合度与期望杂合度相同之外,其余6个种群的观测杂合度均低于期望杂合度,且各地理种群的期望杂合度和观测杂合度之间不存在显著差异,直接排出了各种群存在近交的可能性。 从等位基因丰富度来分析,亚洲其他地区(韩国、俄罗斯、蒙古)西伯利亚狍种群的平均等位基因丰富度为3.68[18],欧洲狍种群的平均等位基因丰富度为4.1[26],而本研究所得到的东北地区西伯利亚狍7个种群的平均等位基因丰富度为7.5,明显高于欧洲狍种群以及亚洲其他地区的西伯利亚狍种群。从杂合度水平来分析,亚洲其他地区(韩国、俄罗斯、蒙古)西伯利亚狍种群的观测杂合度在0.329~0.628之间[18],欧洲狍种群的观测杂合度在0.391~0.686之间[26],其中英国地区种群的观测杂合度在0.49~0.74之间[17]。本研究得到的7个种群的观测杂合度的变化范围在0.70~0.74之间,高于亚洲其他地区西伯利亚狍种群以及欧洲狍种群。研究结果表明,东北地区西伯利亚狍种群遗传一致性较低,种群遗传多样性相对丰富、变异程度较高。 固定指数(Fis)也被称为杂合子缺乏系数,一般被用于评估种群内个体间的近交情况,为-1,则表明种群内全部为杂合子,Fis值为1,则种群内全部为纯合子。研究结果显示,东北地区西伯利亚狍7个种群中,仅黄泥河种群的Fis值为负值(-0.01),其余6个种群的Fis值均>0,在0.021~0.061之间。英国地区欧洲狍种群的Fis值的变化范围为-0.044~0.091[17],亚洲其他地区西伯利亚狍种群的Fis值均大于0,在0.031~0.247之间[18]。可以看出,东北地区西伯利亚狍种群的Fis值变化范围与其他地区狍种群的Fis值比较一致,可能存在一定程度的近交。 本研究还发现调查区域内7个西伯利亚狍种群之间的遗传分化程度较低(FST=0.002~0.074),低于亚洲其他地区西伯利亚狍种群之间的遗传分化程度(0.002~0.412)[18]。需要指出的是,本研究未发现7个西伯利亚狍种群之间存在显著的遗传分化,在后续研究中,需要进一步分析各种群遗传差异的变异来源以及种群间的基因流情况,以探究各种群之间的交流水平。同时拓展研究区域,在更大范围内探究其他地区西伯利亚狍种群与本研究7个种群之间的遗传多样性差异、遗传分化以及基因流情况。 本研究初步建立了东北地区西伯利亚狍种群10个微卫星位点的遗传基础,研究结果表明东北地区西伯利亚狍种群具有较高的遗传多样性,遗传信息丰富:杂合度水平和等位基因丰富度均高于欧洲地区欧洲狍种群以及亚洲其他地区的西伯利亚狍种群。7个种群之间,遗传多样性比较接近,未出现遗传多样性较低的情况。选取的10个微卫星位点的多态性分析也表明7个种群的多态信息含量水平均较高,多态性丰富。这些数据表明,研究区域内的西伯利亚狍种群具有良好的选育和遗传潜力,是可以进行持续利用的种质资源。因此,本研究为以后进行西伯利亚狍种群的合理利用、种群保护提供了一定的理论依据,同时也为下一步分析西伯利亚狍作为猎物种群,对东北虎、东北豹种群的保护和恢复提供了科学依据。 致谢:感谢国家林业局野生动物与自然保护区管理司“虎、东北豹资源调查技术研究”和“东北虎、东北豹种群及栖息地调查评估标准制定及信息汇总”项目的资助。感谢实验室师兄、师姐、师弟和师妹给予的帮助。本研究还得到了浙江大学方盛国教授及其科研团队的指导。实验在野生动物资源学院综合实验室完成,得到了罗理杨老师的指导。野外采样工作得到了项目区各林业局和保护区工作人员的支持和帮助,在这里一同表示感谢。4 结论
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
水产科学(2022年2期)2022-03-20
南京师范大学学报(工程技术版)(2021年2期)2021-10-20
种子(2021年3期)2021-04-12
小读者(2019年20期)2020-01-04
红领巾·成长(2019年3期)2019-04-16
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
中学生理科应试(2016年7期)2016-05-14
