
储藏温度对9种内蒙古半干旱草地植物种子萌发的影响
2018-08-09 01:07韩艳婷江林祥钟明星陈安群刘银占
信阳师范学院学报(自然科学版) 2018年1期
韩艳婷,江林祥,钟明星,陈安群,刘银占*
(1.信阳师范学院 生命科学学院,河南 信阳 464000;2.河南大学 生命科学学院,河南 开封 475004)
0 引言
我国草地面积约4 亿hm2[1,2],占国土面积的40%以上, 养育了近一亿人口[3].其中内蒙古草地面积约为7880 万hm2,占我国草地面积的22%,是我国面积最大的草地[4].该地区不但是我国传统的畜牧业生产基地,还是我国重要的天然绿色生态屏障,在涵养水源[5,6],防风固沙[7],保持生物多样性与维护生态系统平衡[8]方面起着重要作用.这些地区受特殊的地理环境和气候特征影响,成了典型的生态脆弱区,同时又受过度放牧以及其他不合理的土地利用方式的影响,造成草地退化严重[9].改变原有的草地生态系统管理模式,努力恢复该地区草地的群落结构与生态系统功能,促进草地生态系统的可持续发展已经迫在眉睫.
种子是植物最重要的繁殖器官,是草地植物世代更替的主要途径[10,11],也是草地恢复的重要材料之一[12-14].探究不同草地植物种子萌发的限制因子,寻找提高种子萌发率及定植成功率的途径,对于加速草地恢复进程具有重要的理论和现实意义.目前关于种子萌发的影响因素已经进行了大量研究,并发现温度是影响种子萌发的主要因素之一[15,16],每一种植物都有其种子萌发的最适温度[17,18],温度过高[19,20]或过低[21,22]都会降低种子的萌发率.文献[23,24]研究了种子大小与种子萌发率、萌发时间的关系.文献[25-28]以高寒草地和荒漠草原物种为研究对象,对比了恒温储藏和变温储藏下种子萌发的差异,最终确定的受试物种的最佳储存方式分别为湿冷[25]、冷干藏[26,27]、冬季室外埋藏[28],即寒冷的储藏方式对种子萌发有利.半干旱草原的物种与高寒草地[25,26]及荒漠草原[27]的物种无论在形态结构、还是在环境适应方面均存在较大的差异.本研究选取内蒙古9种半干旱草地物种作为研究对象,研究不同储藏温度对种子萌发的影响,以及不同萌发指标之间的关系,确定不同物种的萌发特性,为当地植被恢复提供数据支持.
1 材料与方法
1.1 种子采集地概况
受试植物种子采集于中国科学院植物研究所多伦县十三里滩实验基地(42°02′ N, 116°17′E, 1324 m a. s. l.),该地区年均温2.3 ℃,年均降水量380 mm,属温带大陆性气候.主要植被为半干旱草地,优势物种为冷蒿(Artimesiafrigida)和克氏针茅(Stipakrylovii),建群种有阿尔泰狗娃花(Heteropappusaltaicus)、冰草(Agropyroncristatum)猪毛菜(SalsolacollinaPall)、灰绿藜(Chenopodiumglaucum)等.
1.2 种子收集与保存
选取实验地点常见的9种植物作为研究对象.2种多年生杂类草:阿尔泰狗娃花(H.altaicus)和冷蒿(A.frigida);2种多年生禾类草:冰草(A.cristatum)和克氏针茅(S.krylovii);5种一年生植物:刺藜(Chenopodiumaristatum)、狗尾草(Setariaviridis)、灰绿藜(C.glaucum)、烛台虫实(Corispermumcandelabrum)和猪毛菜(S.Pall).在2014年8—10月份收取每个物种成熟的种子,自然风干后放于牛皮纸信封中,分别放于室温(17 ℃)、低温(4 ℃)和超低温(-18℃)条件下储藏至萌发实验开始.
1.3 种子萌发实验
种子萌发实验于2014年12月1日开始进行,每种植物不同储藏方式的种子各选取100粒,置于直径90 mm培养皿内,采用培养皿纸上法在25 ℃进行萌发预实验,每天观察记录每个培养皿内的种子萌发数量.为了精确统计萌发结果,每次观察记录后将已萌发的种子移出培养皿.
整个预实验过程为1个月.通过预实验,得出不同储藏方式下每个物种大致的萌发开始时间(首次观察到种子开始萌发的时间)与峰值时间(当日种子萌发数量最多的时间).预实验结束后开始正式实验,选取每种储藏方式下每种植物种子150粒,按照50粒/份的标准分成3个重复,分别按照预实验的方法进行种子萌发,并在预实验每组种子萌发开始时间的前两天开始观察记录,观察时间内每天8:00、11:00、14:00、17:00、20:00、22:00各观察记录一次种子萌发状况,以精确计算种子萌发的开始时间和峰值时间,峰值时间1天后,改为每天观察记录一次.根据预实验各组种子的萌发规律,规定当某一组种子在特定时间观察到萌发,但是后续的2天均未观察到有萌发现象时,记为该组种子萌发的结束时间.萌发开始时间到结束时间的时间间隔为种子萌发的持续时间.正式实验阶段持续时间为28 d.实验结束后,统计每组种子整个实验阶段的萌发数量,并除以总实验个体数量(50)后转化为百分数作为该组种子的萌发率.
1.4 统计分析
利用双因素方差分析(two way ANOVA)分析物种和处理对种子萌发率及萌发时间参数的影响.利用单因素方差分析(one way ANOVA)和最小显著差数法多重比较(LSD)分析不同储藏温度对每一物种种子萌发率及萌发时间参数的影响.利用线性回归分析萌发率与萌发时间、不同萌发时间参数之间的关系.统计分析采用SPSS 19.0(纽约IBM公司)完成.
2 结果与分析
2.1 不同物种及储藏温度对萌发率的影响
9种植物种子不同储藏温度下的萌发率见图1,灰绿藜和烛台虫实的萌发率显著低于阿尔泰狗娃花、冰草、克氏针茅和冷蒿.超低温储藏的克氏针茅种子萌发率显著高于其他两种储藏方式.储藏温度显著改变了刺藜、狗尾草、烛台虫实和猪毛菜的萌发率.4种植物室温储存下的萌发率显著高于其他两种储存方式下的萌发率.超低温储藏下刺藜、狗尾草和烛台虫实的萌发率比室温储存下分别降低了32.0%,60.7%和38.0%.低温储藏下狗尾草和猪毛菜的萌发率比室温储存下分别降低了46.0%和13.3%.
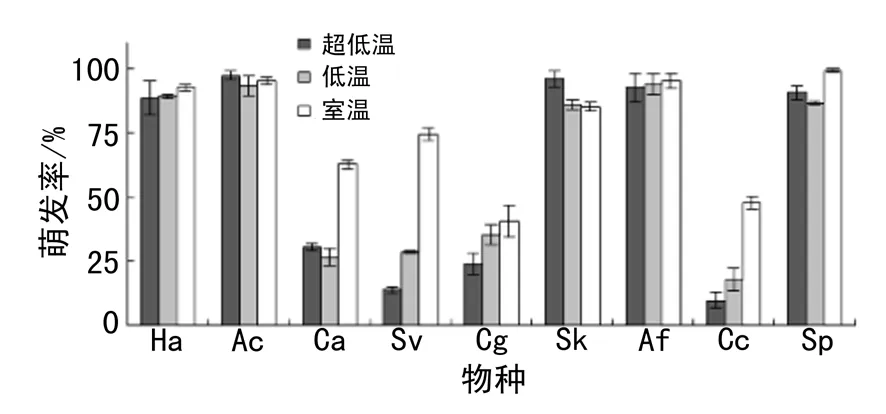
图1 不同物种不同储藏温度下的萌发率(平均值 ± 标准误差)Fig. 1 Germination rate of different species under different storage temperature (Mean ± SE)
注:Ha为阿尔泰狗娃花;AC为冰草;Ca为刺藜;Sv为狗尾草;Cg为灰绿藜;Sk为克氏针茅;Af为冷蒿;Cc为烛台虫实;Sp为猪毛菜;下同
不同物种的萌发率存在显著差异(表1).此外,双因素方差分析结果显示不同储藏温度对种子萌发率有显著影响(表1).综合9个受试物种,超低温和低温储存下的种子萌发率比室温保存的种子萌发率降分别低了16.7%和15.2%.储藏温度对萌发率的影响随物种变化而变化(表1).不同储藏方式对阿尔泰狗娃花、冰草、灰绿藜、冷蒿的种子萌发率无显著影响(表2,所有P> 0.1).储藏温度对克氏针茅萌发率的影响接近显著水平(表2,P< 0.1).
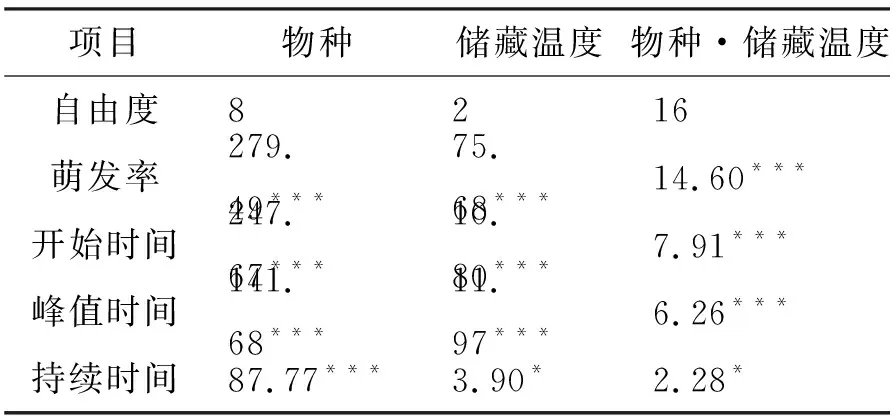
表1 不同物种和储藏温度对种子萌发参数的双因素方差分析结果(F值)Tab. 1 Results (F values) of two-way ANOVAs on the effects of species (Sp) and storage temperature (Tem) on seed germination parameters
注:n=3;*为P< 0.05 ;**为P< 0.01;***为P< 0.001.
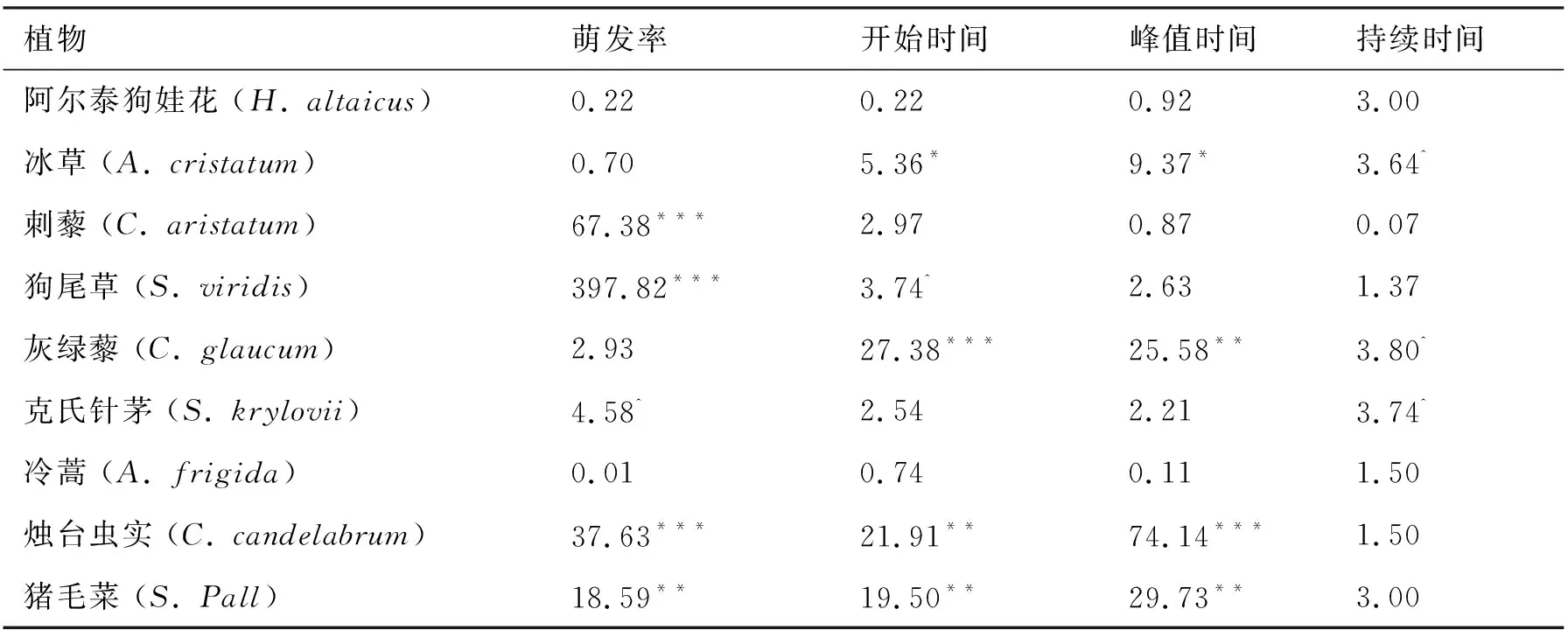
表2 不同储藏温度对每一物种种子萌发参数的单因素方差分析结果(F值)Tab. 2 Results (F values) of one-way ANOVAs on the effect of storage temperature (Tem) on seed germination parameters for each species.
注:n=3; ^表示P< 0.1;*表示P< 0.05;**表示P< 0.01;***表示P< 0.001.
2.2 不同物种及储藏温度对萌发开始时间的影响
不同储藏温度下受试物种的萌发时间见图2.猪毛菜的萌发开始时间最早,克氏针茅的萌发开始时间最晚,多年生禾类草的萌发开始时间较其他植物要晚(图2).低温储藏下冰草的萌发开始时间比超低温晚了1.67 d;超低温储藏下狗尾草的萌发开始时间比室温下晚了0.92 d;超低温和低温储藏下灰绿藜的萌发开始时间比室温储藏的分别晚了0.90、0.80 d;超低温和低温储藏下烛台虫实的萌发开始时间比室温下分别晚了0.79和0.60 d;超低温和低温储藏下猪毛菜的萌发开始时间比室温下分别晚了0.47、0.33 d(图2).方差分析结果显示,萌发开始时间存在显著的种间差异(表1).储藏温度对萌发开始时间同样有显著影响,超低温和低温储藏的种子比室温储藏的种子萌发时间分别晚了0.55、0.45 d.储藏温度对萌发开始时间的影响因物种而异(表1).储藏温度对其余物种的萌发开始时间无显著影响(表2).
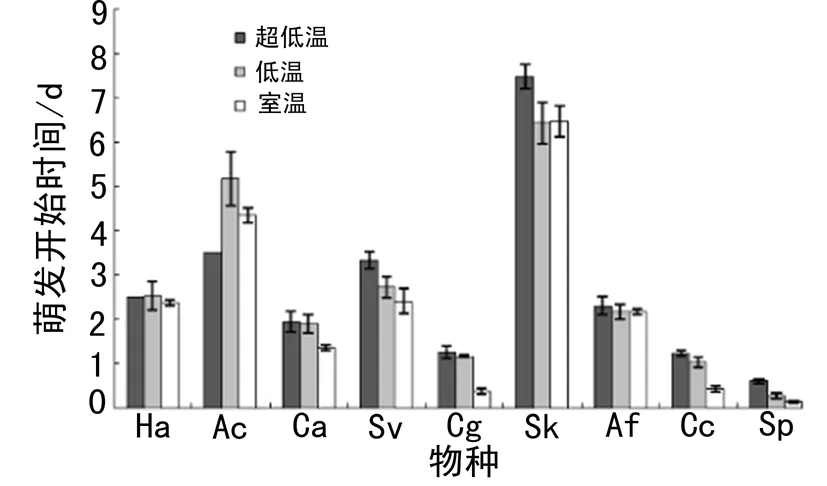
图2 不同物种不同储藏温度下的萌发开始时间(平均值 ± 标准误差)Fig. 2 Germination onset time of different species under different storage temperature (Mean ± SE)
2.3 不同物种及储藏温度对萌发峰值时间的影响
萌发峰值时间随物种和储藏温度的变化趋势与萌发开始时间类似.低温或超低温储存推迟了冰草、灰绿藜烛台虫实和猪毛菜的萌发峰值时间,但对其余物种的萌发峰值时间无显著影响(图3).
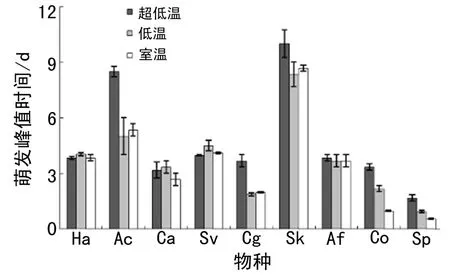
图3 不同物种不同储藏温度下的萌发峰值时间(平均值 ± 标准误差)Fig. 3 Germination peak time of different species under different storage temperature (Mean ± SE)
2.4不同物种及储藏温度对萌发持续时间的影响
不同物种的萌发持续时间不一致(表1).多年生植物尤其是多年生禾类草的萌发持续时间较长,一年生植物的萌发持续时间较短(图4).超低温和低温储藏下种子萌发的持续时间显著延长了0.89、0.93 d(表1,图4).不同物种的萌发持续时间对储藏温度的响应幅度不同(表2).低温储藏的冰草种子萌发时间比室温储藏的延长了0.93 d;超低温储藏的灰绿藜比低温和室温储藏的种子萌发时间分别延长了0.80、0.90 d;超低温储藏的针茅萌发持续时间比低温储藏的延长了1.03 d(图4).温度对其他物种的萌发持续时间无显著影响.
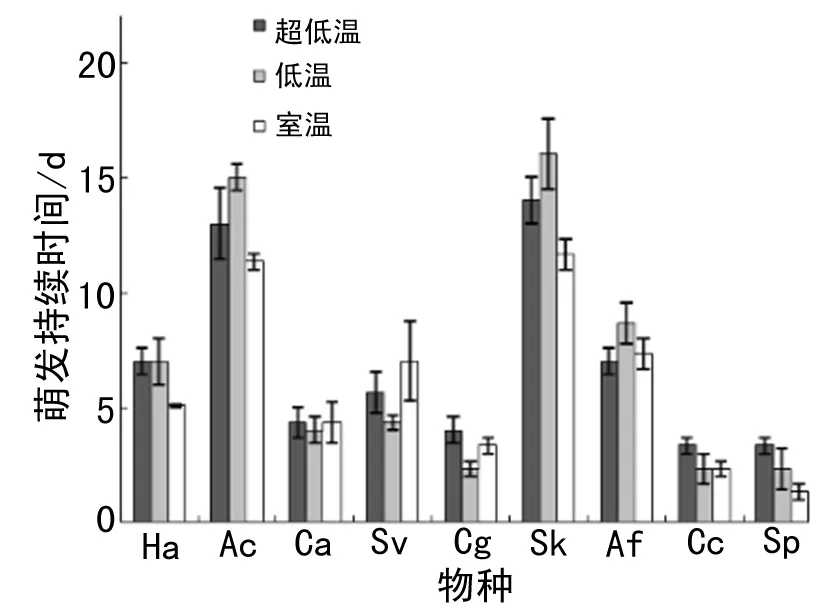
图4 不同物种不同储藏温度下的萌发持续时间(平均值 ± 标准误差)Fig. 4 Germination duration of different species under different storage temperature (Mean ± SE)
2.5 不同萌发参数间的关系
通过利用所有处理和物种的相关参数进行回归分析,种子萌发率与萌发持续时间、开始时间、峰值时间均呈显著的正相关关系(图5).种子萌发持续时间与开始时间、峰值时间之间呈显著的正相关关系,即萌发开始时间或峰值时间越晚,萌发持续时间越长(图6).萌发峰值时间随开始时间的推迟而推迟(图7).
3 结论
不论萌发率还是萌发时间,均存在显著的种间差异.多年生植物的萌发率高于一年生植物,较高萌发率的物种更容易在群落中保持竞争力,因此研究地中针茅和冷蒿的优势度比其他物种大得多.根据萌发特点,参照卜海燕[29]的分类方式,9个受试物种分属于不同的类型.1)爆发型:刺藜、狗尾草、灰绿藜、烛台虫实和猪毛菜均属于此类型,这一类型植物最大的特点是迅速萌芽.回归分析显示,本研究中萌发早的物种具有萌发持续时间短、萌发率低的特点.因为这一类植物在研究地中不占优势,需要迅速利用有利条件完成萌发.保证了其在群落中的存活能力.2)缓萌型:冰草克氏针茅属于此类型,萌发所需时间、持续时间均较长.这类植物的种子较其他物种大很多,根据以前的研究,种子大的物种往往萌发较晚.(3)过渡型萌发型:介于爆发型和缓萌型之间,阿尔泰狗娃花冷蒿属于此类型.萌发类型的多样化有利于植物群落更好地利用资源,对维持群落稳定具有积极的作用.
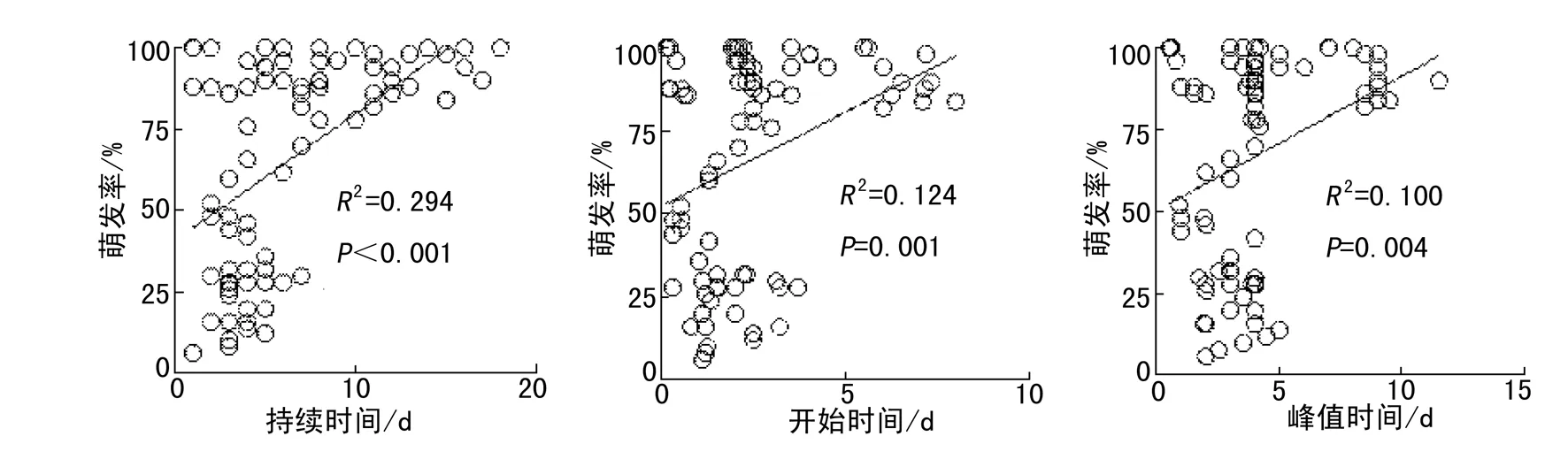
图5 萌发率与持续时间、开始时间及峰值时间的关系Fig. 5 Relationships between germination rate with duration, onset time and peak time
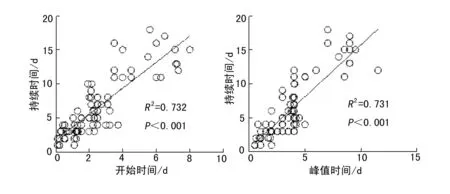
图6 萌发持续时间与开始时间及峰值时间的关系Fig. 6 Relationships between germination duration with onset time peak time with onset time
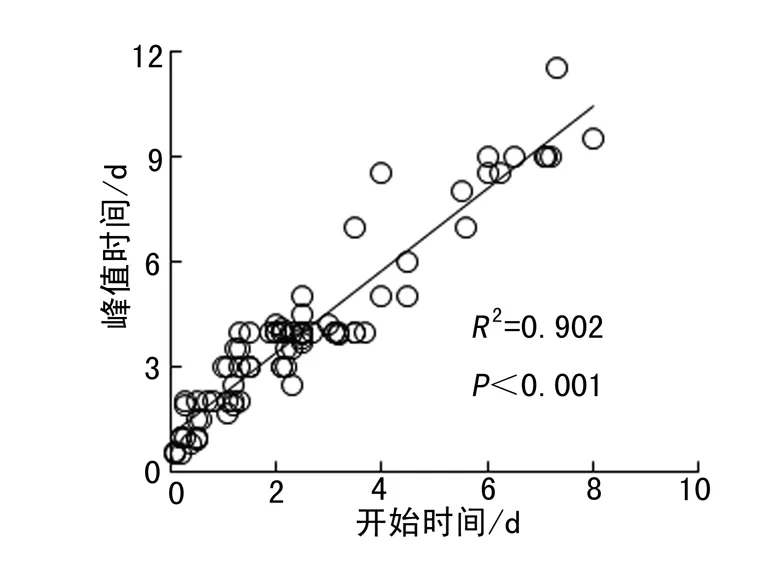
图7 萌发峰值时间与萌发开始时间的关系Fig. 7 Relationships between peak time with onset time
储藏温度会显著影响种子萌发率和萌发时间,室温储藏会显著提高种子萌发率,缩短种子萌发所需的时间,室温储藏对内蒙古草地的种子萌发更为有利.
猜你喜欢
特种结构(2021年6期)2021-12-31
特种结构(2021年6期)2021-12-31
家庭医药(2021年8期)2021-07-28
煤气与热力(2021年2期)2021-03-19
陶瓷学报(2020年2期)2020-10-27
制造技术与机床(2019年4期)2019-04-04
农村农业农民·B版(2017年2期)2017-03-11
科学启蒙(2016年11期)2016-12-09
西藏科技(2015年4期)2015-09-26
橡胶工业(2015年7期)2015-08-29
