
17种植物提取物对大豆疫霉的抑制作用
2018-08-03 01:45马林昊刘凡巧马冠华
现代食品 2018年11期
◎ 许 敏,马林昊,刘凡巧,高 剑,马冠华
(西南大学植物保护学院,重庆 400715)
大豆疫霉根腐病于1989年首次在中国东北地区发现并于1991年被公开报道[1],该病害对大豆生产具有毁灭性损害[2]。目前,该病已经上升为大豆主产区黑龙江省大豆生产中对产量为害最大的病害。大豆疫霉菌(Phytophthora sojae)可侵染大豆生长发育的各阶段,尤其是种子和幼苗阶段,会导致种子腐烂,幼苗根、茎基部腐烂或出现水浸状或立枯并变红褐色,最终引起植株萎蔫、死亡[3-4]。
植物源农药是来源于植物体的农药,其有效成分通常不是单一化合物,而是植物有机体中的多种甚至大部分有机物质[5],具有低毒、低残留、环保友好、不破坏生态环境、靶标有害生物对其不易产生抗药性、对非靶标生物安全、活性成分的作用方式特异等特点,在有机农业领域得到广泛应用[6-7],目前,已经有多种植物源活性成分的农药取得登记并成功应用,如2.5%鱼藤酮乳油、0.5%楝素乳油等[6]。
甲霜灵是有效防治大豆疫霉根腐病的杀菌剂之一,但由于甲霜灵作用位点单一,大豆疫霉菌易对其产生抗药性[8]。笔者利用菌丝生长速率法测定了包括青蒿琥酯、百里酚、肉桂醛、8-甲氧基补骨酯素在内的17种植物提取物对大豆疫霉的作用效果,来寻找可代替甲霜灵等化学药剂的植物提取物。
1 材料与方法
1.1 供试菌株
大豆疫霉野生型菌株P7076,抗甲霜灵突变体Me3-7-3(2 739.81倍 )、Me9-7-2(4 527.93倍 )、Me7-3(9 764.4倍),抗丁吡吗啉突变体Py4-7-2(115.77倍)、Py4-7-3(500.24倍),由西南大学植物生态病理研究所提供。
1.2 供试植物提取物、试剂及材料
青蒿琥酯(99%)、百里酚(≥99%)、肉桂醛(99%)、8-甲氧基补骨酯素(99%),购自上海泰坦科技股份有限公司。5,5-二烯丙基-2,2’二苯基二醇(97%)、β-细辛醚(≥95%)、乙酸丁香酚酯(≥98%)、黄独素B(≥95%)、蛇床子素(≥98%)、香芹酚(99%)、异丁香酚(97%)、4-异丙基苯甲醛(98%)、1,2,3-三苯酚(99%)、杨梅素(97%),购自北京伊诺凯科技有限公司。黄藤素(97%)、α-细辛醚(99%),购自上海麦克林生化科技有限公司。黄酮(99%),购自百灵威科技有限公司。V8蔬菜汁,市售,由美国Campbell公司生产。
1.3 母液的制备
取0.1 g或100 µL植物提取物溶于适量的二甲基亚砜中,配制成104µg/mL母液,于4 ℃冰箱中保存备用。
1.4 培养基的制备
10%V8培养基:100 mL V8蔬菜汁、900 mL ddH2O、10 g CaCO3、20 g琼脂。115 ℃高压灭菌21 min。
1.5 植物提取物对病原菌菌丝生长抑制活性的测定
采用菌丝生长速率法[9]。用无菌水将104µg/mL母液稀释成适宜的浓度,在培养基中加入一定量的药剂,配成相应浓度的含药培养基,充分摇匀,倒入直径为6 cm的培养皿中,每皿10 mL,以等量无菌水配成无药培养基平板作为空白对照,每个浓度3次重复。将供试菌株于25 ℃下黑暗培养4 d,用直径为5 mm的打孔器在靠近菌落边缘的同一圆周上打取菌饼,然后将菌饼菌丝面朝下接种到各平板中央,并在25 ℃下黑暗培养4 d后,用十字交叉法测定各个处理的菌落生长直径。按照下面公式求出各个药剂浓度对菌丝生长的抑制率。

最后将药剂的浓度转化为对数(x),抑制率转化为机率值(y),根据x与y的线性关系求出毒力回归曲线方程y=a+bx、相关系数R、有效抑制中浓度EC50。
2 结果与分析
2.1 17种植物提取物对大豆疫霉野生型菌株的抑制作用
17种植物提取物对大豆疫霉野生型菌株P7076的菌丝生长均有一定的抑制作用,厚朴酚的抑制作用最佳,其EC50值为17.032 5 μg/mL。其中,EC50值在50 μg/mL以下的有厚朴酚、香芹酚、黄藤素、百里酚和1,2,3-三苯酚5种;EC50值在50~100 μg/mL的有异丁香酚、肉桂醛、黄酮、青蒿琥酯、黄独素B和乙酸丁香酚酯6种,EC50值在100~200 μg/mL的有杨梅素、α-细辛醚、蛇床子素、8-甲氧基补骨酯素和β-细辛醚5种;17种植物提取物中,效果最差的是枯茗醛,其EC50值为201.803 0 μg/mL,见表1。

表1 17种植物提取物对大豆疫霉野生型菌株P7076菌丝生长的抑制作用表
2.2 17种植物提取物对大豆疫霉抗甲霜灵突变体的抑制作用
厚朴酚对抗甲霜灵突变体菌丝生长的抑制作用最佳,3株抗性菌株的EC50值分别为14.879 2、17.294 0和14.427 7 μg/mL,其中厚朴酚、香芹酚、青蒿琥酯、黄酮、黄藤素、百里酚、1,2,3-三苯酚、异丁香酚、8-甲氧基补骨酯素、肉桂醛的EC50值在10~65 μg/mL,乙酸丁香酚酯、杨梅素、黄独素B、α-细辛醚、蛇床子素、β-细辛醚的EC50值在95~160 μg/mL,抑制效果较好;效果最差的是枯茗醛,3株抗性菌株的EC50值分别为505.779 7、402.447 6、326.589 9 μg/mL。经统计分析表明,17种植物提取物对3株抗甲霜灵突变体的抑制作用无显著性差异,即抑制作用与抗性倍数无相关性,见表2。
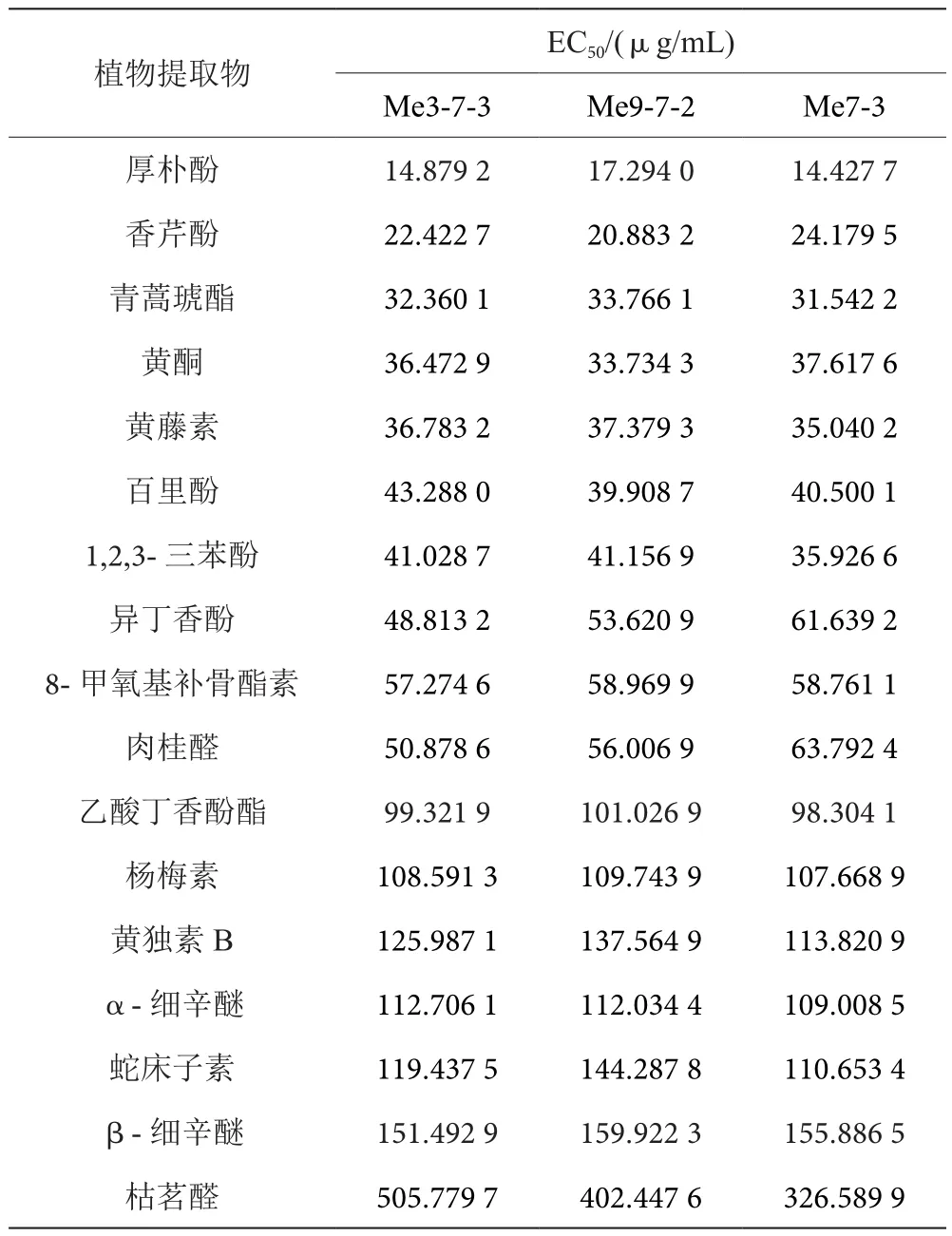
表2 17种植物提取物对大豆疫霉抗甲霜灵突变体的抑制作用表
2.3 17种植物提取物对大豆疫霉抗丁吡吗啉突变体的抑制作用
厚朴酚对抗丁吡吗啉突变体菌丝生长的抑制作用最佳,2株抗性菌株的EC50值分别为13.941 9、14.785 4 μg/mL,其中厚朴酚、香芹酚、青蒿琥酯、黄酮、黄藤素、百里酚、1,2,3-三苯酚、异丁香酚、8-甲氧基补骨酯素、肉桂醛的EC50值在10~50 μg/mL,乙酸丁香酚酯、杨梅素、黄独素B、α-细辛醚、蛇床子素、β-细辛醚、枯茗醛的EC50值在90~190 μg/mL,抑制效果都较好。经统计分析表明,17种植物提取物对2株抗丁吡吗啉突变体的抑制作用无显著性差异,即抑制作用与抗性倍数无相关性,见表3。

表3 17种植物提取物对大豆疫霉抗丁吡吗啉突变体的抑制作用表
2.4 野生型菌株与抗性菌株之间抑制作用的比较
通过比较17种植物提取物对3种类型的大豆疫霉菌株之间的抑制作用发现,青蒿琥酯、黄酮和8-甲氧基补骨酯素3种提取物对野生型菌株P7076的EC50值显著高于抗性菌株的EC50值,即这3种提取物对抗性菌株的抑制作用更好(表4),其余14种植物提取物对野生型菌株和两种药剂的抗性菌株抑制作用无明显差异。
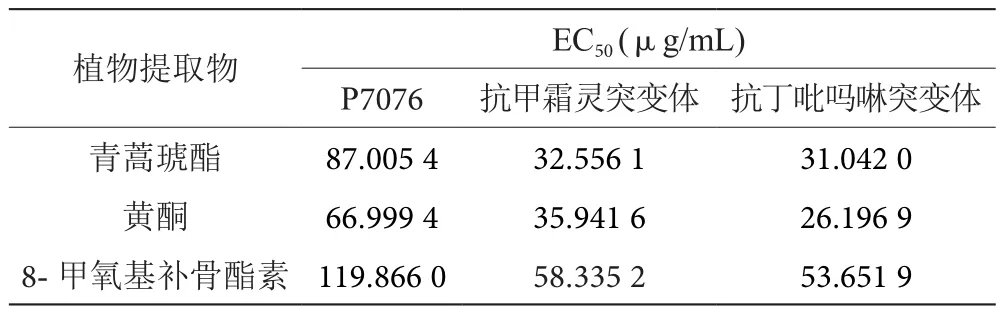
表4 野生型菌株与抗性菌株之间抑制作用的比较表
3 讨论
植物是生物活性化合物的巨大天然宝库,植物中产生的次生代谢产物超过40万种,其中许多次生化学物质具有杀虫或抑菌的生物活性,在农业上有着重要的研究价值[10]。目前,国内外对植物提取物对病原菌抑制作用的研究非常活跃[11-12]。有的植物提取物甚至可以直接用于一些农作物病害的防治[13]。研究发现,青蒿琥酯、蛇床子素和厚朴酚对荔枝霜疫霉菌均有用于开发防治荔枝霜疫霉的植物源农药的潜力,对荔枝霜疫霉菌菌丝生长抑制作用中,厚补酚的EC50值约为31 μg/mL,青蒿琥酯的EC50值约为133 μg/mL,蛇床子素的EC50值约为138 μg/mL[14];本试验对大豆疫菌丝生长的抑制测定发现,厚补酚的EC50值约为15 μg/mL,青蒿琥酯的EC50值约为41 μg/mL,蛇床子素的EC50值约为120 μg/mL,说明厚朴酚、青蒿琥酯和蛇床子素相较于荔枝霜疫霉对大豆疫霉的抑制效果更好。百里酚与香芹酚是同分异构体,研究发现其对褐腐菌的菌丝扩展和孢子萌发均具有较好的抑制能力[15],本试验中百里酚对大豆疫菌丝生长的抑制作用的EC50值约为40 μg/mL,说明百里酚对大豆疫霉也有较好的防治效果。
本试验发现,除枯茗醛外的16种植物提取物对大豆疫霉野生型菌株和两种药剂的抗性菌株均有较好的抑制作用,其EC50值均低于200 μg/mL,对于3种类型的菌株,抑制效果最好的是厚朴酚,EC50值约为15 μg/mL,其次是香芹酚,EC50值约为23 μg/mL,均低于常用药剂霜霉威对大豆疫霉的菌丝生长抑制作用的EC50值[16],表明厚朴酚和香芹酚具有开发为植物源农药的潜力。另外,青蒿琥酯、黄酮和8-甲氧基补骨酯素3种植物提取物对野生型菌株的EC50值显著高于抗性菌株的EC50值,即这3种提取物对抗性菌株的抑制作用更好,是否由于抗药突变体的突变位点与该植物提取物的作用位点一致还需要进一步研究。
本试验测定了17种植物提取物对大豆疫霉的菌丝生长的抑制效果,要全面评估这些植物提取物的开发价值,还需要测定其对游动孢子囊产量、卵孢子产量等方面的影响,更全面地说明这些植物提取物的抑制作用。同时,目前虽然不少关于植物提取物的抑制作用研究报道,但大多数仍然停留在实验室水平,真正应用于农作物病害防治的比较少[17-20]。因此,还需要对这些植物提取物进行田间应用效果测试,以为植物源农药的开发应用提供依据。
猜你喜欢
保健医苑(2021年7期)2021-08-13
绵阳师范学院学报(2021年5期)2021-05-28
新世纪智能(高一语文)(2020年5期)2020-07-24
河北大学学报(自然科学版)(2020年1期)2020-01-15
广东农业科学(2017年5期)2017-08-29
中成药(2017年3期)2017-05-17
中国民族民间医药(2016年24期)2017-01-07
天然产物研究与开发(2016年6期)2016-06-05
散文选刊·下半月(2016年9期)2016-05-14
中国烟草学报(2012年4期)2012-04-09
