
蒙古高原种子植物属的区系地理分析
2018-07-28 08:18,,,*
草地学报 2018年2期
, , , *
(1. 内蒙古自治区林业监测规划院, 内蒙古 呼和浩特 010020; 2. 内蒙古农业大学草原与资源环境学院, 内蒙古 呼和浩特 010019)
蒙古高原地处亚洲大陆腹地干旱与半干旱地区(87°40′~122°15′ E,37°46′~53°08′ N)。南以阴山、贺兰山、祁连山为界,北至唐努山、萨彦岭、肯特山以及雅布洛诺夫山脉一带,东起大兴安岭,西部为萨彦岭和阿尔泰山脉所环绕[1]。蒙古高原大致包括蒙古国全部地区,俄罗斯联邦西伯利亚部分地区,以及中国内蒙古自治区、甘肃和新疆维吾尔自治区的部分地区,广义的蒙古高原还包括阴山以南被黄河包围的鄂尔多斯高原。
蒙古高原宏观地貌主要为山地和高平原,平均海拔1580m,整体地势西高东低[1]。蒙古高原周边为中、高山地所环绕,远离海洋,阻挡了来自远海和远洋吹来的湿润气流,成为一个强烈的大陆性气候区[2]。气候特点为冬季漫长寒冷,夏季短暂炎热,降水稀少,且分布极不均匀,除高原东部、东南部以及北部等少数地区外,绝大部分地区年降水均少于400mm[1]。在山地半湿润气候带形成了森林、草甸和草原的植被组合,在广大的半干旱气候带,发育形成了草原,在干旱地带发育着荒漠草原植被,在极干旱气候带是荒漠植被[3]。
1 材料与方法
2008-2010年间,野外调查以蒙古高原的重要地区为原点进行多方位呈辐射状逐段调查,根据野外考察线路设置植物样地,对不同生境不同植被类型的区域进行植物种类调查,并采集制作标本[4]。室内数据处理结合内蒙古、甘肃和新疆地区各大学的植物标本馆标本,并参考相关资料[5-17]准确鉴定出物种名称,系统整理出研究区种子植物名录,同时运用植物区系地理学基本原理和方法,通过统计、排序以及比较分析的方法,对研究区种子植物区系进行了系统的研究和详细的论述。
2 结果与分析
2.1 植物种类组成
本区种子植物共有4 280种(含种下分类单位,不包括栽培种),隶属于761属115科。其中裸子植物3科、8属、32种;被子植物112科、753属、4 248种(双子叶植物92科、610属、3 333种;单子叶植物20科、143属、915种),占区域种子植物总种数的99.3%,在数量上占绝对优势。从各类群所含的科、属、种数占世界区系相应类群的比例(表1)可见,蒙古高原种子植物区系相对贫乏,所含种数仅占世界种子植物种数的1.9%(远低于中国所占比例12.1%),且以草原植被和荒漠植被为主,说明蒙古高原自然环境较为严酷,这与蒙古高原所处的半干旱和干旱地区的地理位置相适宜。
2.2 属的组成分析
对属的大小统计表明(表2),本区域单型属312属,含312种,占总属数的41.0%,总种数的7.3%,其中,单种属80属,占区域总属数的10.5%;少型属(2~9种)共338属,含1 317种,占总属数的44.6%,占总种数的30.8%;中型属(10~19种)共71属,含911种,占总属数的9.3%,占总种数的21.3%;较大属(20~29种)共22属,含519种,占总属数的2.9%,占总种数的12.1%;大型属(≥30种)只有18属,共计1 221种,仅占本区域总属数的2.4%,却占总种数的28.5%。这说明本区域属的分化较大,中型属、小型属非常丰富,大型属较少,却包含较多的种,比较发达,在植被构成中占据着十分重要的地位。从种属比例看,大型属、较大属和中型属的种属比例分别是67.8、23.6和12.8,明显高于区域植物种属比的平均值5.6,说明大型属、较大属和中型属在种水平上丰富度较高,具有一定的优势。

表1 蒙古高原植物大类群统计Table 1 Statistic of communities in Mongolian Plateau
注:*裸子植物的世界资料来源于J.C.Willis(1973)[18].**被子植物世界资料来源于R.Good(1974)[19]
Note:*indicates the Gymnosperms data of the World from J.C.Willis(1973)[18].**indicates the Angiosperms data of the World from R.Good(1974)[19]
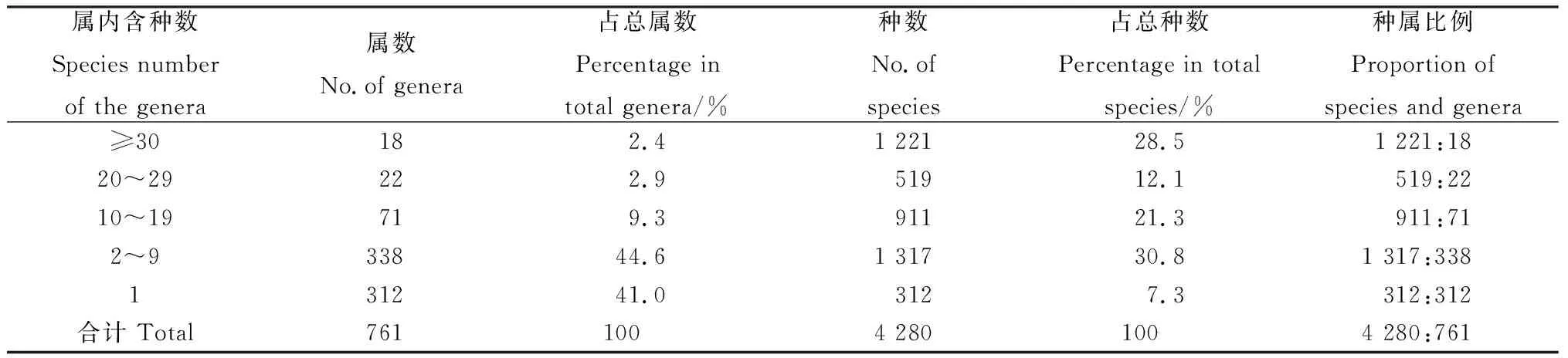
表2 蒙古高原种子植物属的统计Table 2 Genera of spermatophyte in Mongolian Plateau
2.3 属的地理成分分析
参考吴征镒先生对中国种子植物属的分布区类型的划分,将蒙古高原种子植物761属划分成15个分布区类型和22个变型(表3)[20-21]。
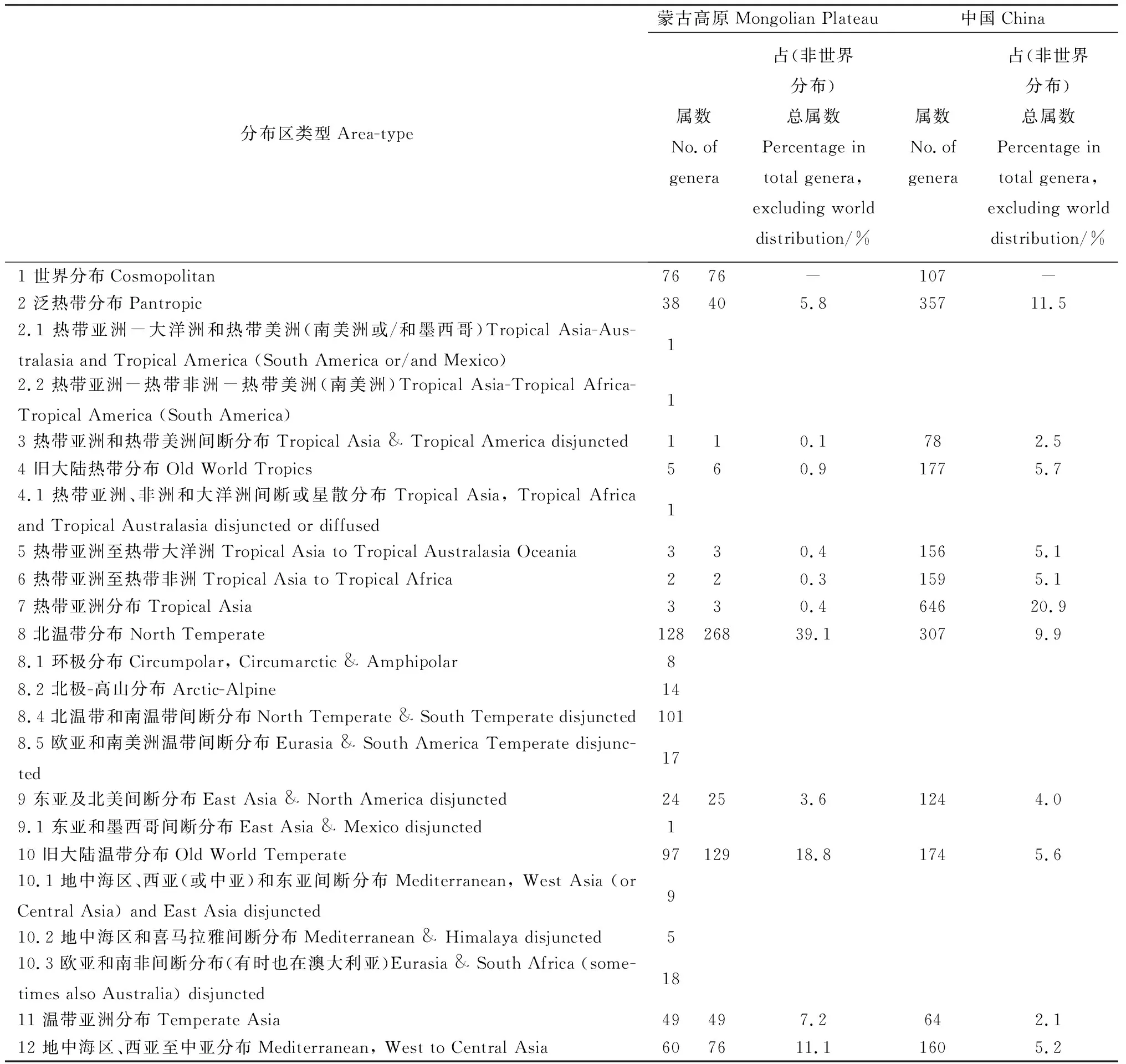
表3 蒙古高原种子植物属的分布区类型Table 3 The distribution types of spermatophyte genera in Mongolian Plateau

续表3
2.3.1世界分布属 本类型蒙古高原有76属,占总属数的10.0%,远高于我国该类型所占的比例(3.5%)[22]。从生态型来看,该类型的属是以温带起源的喜湿或中生草本植物为主[23],且广布于蒙古高原。如在各地草甸分布的有千里光属(Senecio)、龙胆属(Gentiana)、老鹳草属(Geranium)、酸模属(Rumex)、黄芩属(Scutellaria)、远志属(Polygala)、毛茛属(Ranunculus)和苔草属(Carex)等;在各地湖沼及河岸浅水中分布的有莎草属(Cyperus)、藨草属(Scirpus)、灯心草属(Juncus)、香蒲属(Typha)、浮萍属(Lemna)、眼子菜属(Potamogeton)、狸藻属(Utricularia)、茨藻属(Najas)等。区域内还有少数含木本植物的属,如槐属(Sophora)、悬钩子属(Rubus)、鼠李属(Rhamnus)、蒿属(Artemisia)、卫矛属(Euonymus)和猪毛菜属(Salsola),是灌丛的主要组成成分,唯有槐属和卫矛属可达到乔木的高度。其中不乏有少数属,如猪毛菜属、旋花属(Convolvulus)、蒿属、碱蓬属(Suaeda)、盐角草属(Salicornia)、黄芪属(Astragalus)、和补血草属(Limonium)等,主要产于干旱地区或盐渍化的生境上,是荒漠植被或盐生草甸的主要植被。
2.3.2泛热带分布 本类型蒙古高原有40属,其中热带亚洲-大洋洲和热带美洲分布变型1属,热带亚洲-热带非洲-热带美洲分布变型1属,共占蒙古高原总属数的5.8%,低于我国该类型所占的比例(11.5%)[22],这些属中没有仅限于热带地区分布的属。该类型的木本属只有朴属(Celtis)、一叶萩属(Securinega)、牡荆属(Vitex)、枣属(Zizyphus)和醉鱼草属(Buddleja),它们主要分布于蒙古高原温带地区的森林、山地灌丛及沟谷,几乎不进入寒温带地区。该类型的草本植物属多为单种属或寡种属,可见这些泛热带属在蒙古高原这一辽阔的地区种系发育得并不好。
2.3.3热带亚洲和热带美洲间断分布 本类型蒙古高原仅有1属,为地榆属(Sanguisorba),常生于草原、草甸、山坡草地、灌丛中或疏林下,占蒙古高原总属数的0.1%,明显低于我国该类型所占的比例(2.5%)[23]。可见,蒙古高原植物区系与热带亚洲和热带美洲植物区系曾有共同的渊源,这是由于热带美洲(南美洲)本来位于古南大陆的西部,最早于侏罗纪末期和非洲开始分离,至白垩纪末期与之完全分离,之后热带亚洲和热带美洲的植物又都经过漫长的演化和发展历程,使种群逐步扩散到温带地区,造就了蒙古高原种子植物区系与热带亚洲和热带美洲植物区系的微弱联系[23]。
2.3.4旧大陆热带分布 本类型蒙古高原有6属,其中热带亚洲、非洲和大洋洲间断或星散分布变型1属,共占总属数的0.9%,远低于我国同类型属数的比例(5.7%)[22]。其中,天门冬属(Asparagus)、百蕊草属(Thesium)广泛分布于蒙古高原各类生境中。槲寄生属(Viscum)、虎舌兰属(Epipogium)主要分布在蒙古高原的森林地区。牛鞭草属(Hemarthria)、荩草属(Arthraxon)常见于蒙古高原的各类草甸中。
2.3.5热带亚洲至热带大洋洲 本类型蒙古高原有3属,占总属数的0.4%,明显低于我国同类型属所占的比例(5.1%)[22]。其中臭椿属(Ailanthus)是该类型中唯一一个作为优势树种能够分布到蒙古高原南部地区的森林植被中[23]。大豆属(Glycine)和犁头尖属(Typhonium)作为伴生种,常分布于蒙古高原地区的河岸、灌丛、山地或田野。
2.3.6热带亚洲至热带非洲 本类型蒙古高原有2属,分别为杠柳属(Periploca)和芒属(Miscanthus),其中的2个变型(华南、西南到印度和热带非洲间断分布和热带亚洲和东非间断分布)均缺,占总属数的0.3%,明显低于我国同类属所占的比例(5.1%)[22],这与蒙古高原所处的气候带有关。
2.3.7热带亚洲分布 本类型蒙古高原仅有3属,分别为赤爮属(Thladiantha)、苦荬菜属(Ixeris)和小苦荬属(Ixeridium),占总属数的0.4%,远低于全国同类型属数的比例(20.9%)[22]。它们在蒙古高原常出现在林缘、山坡草地、河边、沟谷甚至是沙地,这一类型在蒙古高原植物区系中所起的作用很小。
2.3.8北温带分布 本类型蒙古高原有268属,其中环极分布变型8属、北极-高山分布变型14属、北温带和南温带间断分布变型101属、欧亚和南美洲温带间断分布变型17属,共占总属数的39.1%,明显高于全国同类属所占的比例(9.9%)[22]。这一类型在属和种的数量上均占据着最高的比例,在蒙古高原植物区系组成上占有极为重要的作用。它们的水分生态型多为中生的木本或草本植物。前者如裸子植物的冷杉属(Abies)、云杉属(Picea)、落叶松属(Larix)、松属(Pinus)、刺柏属(Juniperus)、圆柏属(Sabina)均为山地针叶林植被的重要组成成分,尤以落叶松属和松属最为突出,可作为蒙古高原山地植物群落的优势种和建群种,也呈斑块状少量进入森林草原地带;被子植物的杨属(Populus)、桦木属(Betula)、栎属(Quercus)、榆属(Ulmus)、槭树属(Acer)、椴树属(Tilia)、接骨木属(Sambucus)、花楸属(Sorbus)、桑属(Morus)是构成蒙古高原落叶阔叶林和针阔叶混交林的主要成分,其中多数种是区域森林植被的建群种或优势种。还有一些灌木属如山梅花属(Philadelphus)、榛属(Corylus)、越橘属(Vaccinium)、蔷薇属(Rosa)、杜鹃花属(Rhododendron)、绣线菊属(Spiraea)、茶藨属(Ribes)、荚蒾属(Viburnum)、忍冬属(Lonicera)、胡颓子属(Elaeagnus)是构成植物群落下木层的主要成分或山地落叶灌丛的主要建群植物。溪谷、河岸两侧及河流下游的河床上常分布有柳属(Salix)植物,部分地区还可见梾木属(Swida)、桤木属(Alnus)、绣线菊属和蔷薇属植物。此外,该类型的草本属更是发达。双子叶植物如龙芽草属(Agrimonia)、蚊子草属(Filipendula)、舞鹤草属(Maianthemum)、委陵菜属(Potentilla)、迷果芹属(Sphallerocarpus)、景天属(Sedum)等及单子叶植物如铃兰属(Convallaria)、舞鹤草属(Maianthemum)、黄精属(Polygonatum)等均为山地林下或山地草甸植物的主要代表;双子叶植物远志属(Polygala)、棘豆属(Oxytropis)和单子叶植物针茅属(Stipa)、碱茅属(Puccinellia)、披碱草属(Elymus)、赖草属(Leymus)和拂子茅属(Calamagrostis)等旱生植被,是蒙古高原典型草原和山坡草地的优势成分。岩高兰属(Empetrum)、杜香属(Ledum)、北极花属(Linnaea)、毛蒿豆属(Oxycoccus)、松毛翠属(Phyllodoce)、冰沼草属(Scheuchzeria)、熊果属(Arctostaphylos)、北极果属(Arctous)、肉果荠属(Braya)、单侧花属(Orthilia)、裂稃茅属(Schizachne)、岩须属(Cassiope)和山嵛菜属(Eutrema)等,常常是山地冻原、高山植被或寒温带针叶林下的代表植物或重要成分。
2.3.9东亚和北美间断分布 本类型蒙古高原有25属,包括东亚和墨西哥间断分布变型1属,占总属数的3.6%,与全国同类属所占的比例(4.0%)[22]相近。含木本的属仅有珍珠梅属(Sorbaria)、罗布麻属(Apocynum)、八仙花属(Hydrangea)、胡枝子属(Lespedeza)和五味子属(Schisandra)5属。除罗布麻属会分布到较为干旱的盐碱荒地、沙漠边缘、戈壁荒滩及河流两岸、冲击平原、河湖周围外,其余4属主要是蒙古高原森林中的重要成分,但胡枝子属的草本种也常分布到草原中。这一间断分布的成分或在联合古陆解体之前的白垩纪就已存在于北美及东亚,或在联合古陆分离后继续演化发展而来,并呈现出现今的间断分布格局[24]。
2.3.10旧大陆温带分布 本类型蒙古高原有129属,包括地中海区、西亚(或中亚)和东亚间断分布变型9属、地中海区和喜马拉雅间断分布变型5属、欧亚和南非间断分布变型18属,共占总属数的18.8%,远高于全国同类属所占的比例(5.6%)[22],这一类型及变型在蒙古高原植物区系中占的比例仅次于北温带分布型及其变型。从生活型上看,木本属仅有沙棘属(Hippophae)、柽柳属(Tamarix)、水柏枝属(Myricaria)、梨属(Pyrus)、栒子属(Cotoneaster)、桃属(Amygdalus)、木蓼属(Atraphaxis)、鲜卑花属(Sibiraea)和丁香属(Syringa)9属,占该类型属数的7.0%,其中柽柳属、水柏枝属和木蓼属是蒙古高原荒漠地区极为广布的重要的优势属和建群属,其它木本属均为森林的重要组成成分。中生或旱中生的草本属占大多数,如芨芨草属(Achnatherum)、冰草属(Agropyron)、隐子草属(Cleistogenes)常成为典型草原地区的优势或建群植物,其他草本属如沙参属(Adenophora)、荆芥属(Nepeta)、聚合草属(Symphytum)、白鲜属(Dictamnus)、滇紫草属(Onosma)、菊属(Dendranthema)、蓝盆花属(Scabiosa)、乳菀属(Galatella)、蓍属(Achillea)、萱草属(Hemerocallis)、沼兰属(Microstylis)、芯芭属(Cymbaria)、败酱属(Patrinia)、石竹属(Dianthus)、麻花头属(Serratula)、樟味藜属(Camphorosma)、蜡菊属(Helichrysum)等,为林下草本层、高山灌丛、山地草甸、草甸、典型草原、荒漠草原的重要组成成分。
2.3.11温带亚洲分布 本类型蒙古高原有49属,占总属数的7.2%,远高于全国同类属所占的比例(2.1%)[22]。含木本的属仅有锦鸡儿属(Caragana)、杏属(Armeniaca)和亚菊属(Ajania)3属,其余均为草本植物,主要有线叶菊属(Filifolium)、马兰属(Kalimeris)、狗娃花属(Heteropappus)、米口袋属(Gueldenstaedtia)、防风属(Sapoghnikovia)、轴藜属(Axyris)、裂叶荆芥属(Schizonepeta)、狼毒属(Stellera)、细柄茅属(Ptilagrostis)等。其中,锦鸡儿属、亚菊属、线叶菊属、细柄茅属等常是草原中的建群植物。
2.3.12地中海区、西亚至中亚分布 本类型蒙古高原有76属,包括地中海区至中亚和南非和/或大洋洲间断分布变型8属、地中海区至西亚或中亚和墨西哥或古巴间断分布变型1属、地中海区至温带-热带亚洲、大洋洲和/或北美南部至南美洲间断分布变型3属、地中海区至北非、中亚-蒙古、西南非洲、西南澳大利亚、北美西南部、智利西部(泛地中海)星散分布变型3属、马加罗尼西亚、地中海欧洲、索科特拉、西亚、南至索马里分布变型1属,共占总属数的11.1%,远高于全国同类属所占的比例(5.2%)[22],这一类型及变型在蒙古高原植物区系中所占的比例居第三位。这一成分中单型属和少型属很丰富。从生态类型上看,多是旱生和盐生的种类,它们是蒙古高原荒漠草原、荒漠以及盐沼、碱地的隐域植被的重要组成者,在蒙古高原植物区系中起着十分重要的作用,常常成为群落的优势植物或建群植物,甚至形成单优势群落。如红砂属(Reaumuria)、半日花属(Helianthemum)、假木贼属(Anabasis)、梭梭属(Haloxylon)、驼绒藜属(Ceratoides)、小蓬属(Nanophyton)、盐爪爪属(Kalidium)、单刺蓬属(Cornulaca)、裸果木属(Gymnocarpos)、沙拐枣属(Calligonum)、霸王属(Zygophyllum)、白刺属(Nitraria)、骆驼蓬属(Peganum)、苓菊属(Jurinea)、补血草属(Limonium)、花花柴属(Karelinia)及禾本科的獐茅属(Aeluropus)等。还有一些短命、类短命植物及珍贵的肉质寄生植物,如锁阳(Cynomorium)及肉苁蓉属(Cistanche)等。
2.3.13中亚分布 本类型蒙古高原有52属,包括中亚东部分布变型19属、中亚东部至喜马拉雅和中国西南部变型10属、西亚至喜马拉雅和西藏分布变型2属、中亚至喜马拉雅-阿尔泰和太平洋北美洲间断分布变型1属,共占总属数的7.6%,明显高于全国同类属数所占的比例(4.0%)[22]。这一类型单型属和少型属更为丰富,它们多是旱生或耐旱的草本或木本,多生于山前荒漠、砾石戈壁、丘间地。典型的中亚分布属有矮刺苏属(Chamaesphacos)、翅膜菊属(Alfredia)、短舌菊属(Brachanthemum)、寒蓬属(Psychrogeton)、琉苞菊属(Hyalea)、绒藜属(Londesia)、四齿芥属(Tetracme)、兔唇花属(Lagochilus)、蛛丝蓬属(Micropeplis)、紫菀木属(Asterothamnus)等;分布于亚洲中部的戈壁藜属(Iljinia)、革苞菊属(Tugarinovia)、合头草属(Sympegma)、沙蓬属(Agriophyllum)、沙冬青属(Ammopiptanthus)、栉叶蒿属(Neopallasia)、沙芥属(Pugionium)等属均是比较古老的荒漠植物。扁芒菊属(Waldheimia)、高河菜属(Megacarpaea)、角蒿属(Incarvillea)、长柱琉璃草属(Lindelfia)、拟耧斗菜属(Paraquilegia)等,主要是蒙古高原高山上衍生的比较年轻的成分。这类成分均为地中海区、西亚至中亚分布成分在亚洲地区经过强烈旱化的衍生物,由于天山山脉和青藏高原的抬升,产生山地特有现象,从北温带和旧大陆温带广布的大属中进一步分化出一些新属,如女蒿属(Hippolytia)和沙鞭属(Psammochloa)分别是从菊蒿属(Tanacetum)和芨芨草属(Achnatherum)分化衍生而来。个别的如角蒿属(Incarvillea)则远达四川和贵州,可以说这些高原高山成分是邻近亚热带森林区系经过高山旱化和特化的结果[23]。
2.3.14东亚分布属 本类型蒙古高原有21属,包括中国-喜马拉雅分布变型6属、中国-日本分布变型7属,共占总属数的3.1%,明显低于全国同类属数的比例(10.2%)。本类型以森林成分为主,如侧柏属(Platycladus)在蒙古高原南部山地往往形成单优势群落;五加属(Eleutherococcus)、锦带花属(Weigela)、斑种草属(Bothriospermum)、翅果菊属(Pterocypsella)、党参属(Codonopsis)、裂瓜属(Schizopepon)、兔儿伞属(Syneilesis)、翠菊属(Callistephus)、黄瓜菜属(Paraixeris)、桔梗属(Platycodon)、松蒿属(Phtheirospermum)等成为林下或林缘草甸的常见成分。也有分布在干旱、半干旱区山地的属,如野丁香属(Leptodermis)、莸属(Caryopteris)、地黄属(Rehmannia)。还有部分属是组成山地草甸或河滩草甸的重要成分,如甘松属(Nardostachys)、微孔草属(Microula)、肉果草属(Lancea)等。
2.3.15蒙古高原特有属 蒙古高原仅有10个特有属,其中,真特有属[25]有沙芥属(Pugionium)、沙鞭属(Psammochloa)、连蕊芥属(Synstemon)、绵刺属(Potaninia)[26]、四合木属(Tetraena)、革苞菊属(Tugarinovia)、百花蒿属(Stilpnolepis)、紊蒿属(Elachanthemum);准特有属[25]有脓疮草属(Panzeria)和芯芭属(Cymbaria)。
3 讨论
3.1 特有现象
蒙古高原分布的761属中仅有10个属是蒙古高原特有属,只占总属数的1.5%,远低于全国特有属所占的比例(8.1%)[22],特有程度较低。根据各特有属分布区域的不同,又划分为7个特有分布变型。如沙芥属(Pugionium)和沙鞭属(Psammochloa)为蒙古高原沙地分布的特有属;连蕊芥属(Synstemon)为阿拉善荒漠南缘低山丘陵分布的特有属;绵刺属(Potaninia)[26]为阿拉善荒漠特有属;四合木属(Tetraena)和革苞菊属(Tugarinovia)为东阿拉善-西鄂尔多斯特有属;百花蒿属(Stilpnolepis)为东阿拉善-鄂尔多斯特有属;紊蒿属(Elachanthemum)为戈壁蒙古分布的特有属;脓疮草属(Panzeria)和芯芭属(Cymbaria)[28]则是主要分布在蒙古高原的准特有属。
10个特有属在区系上全部都是单种属或双种中群,分类上比较孤立,但全部是被子植物中各个所属科中的最进化类群,在生态上绝大多数是荒漠种和半荒漠种,均为旱生植物,且沙生植物占优势,它们的生活型以草木植物占绝对优势。这10个属在地理分布上集中分布于蒙古高原的阿拉善-鄂尔多斯地区,东阿拉善-西鄂尔多斯是其分析中心,起源上多数是东亚与地中海植物区系的后裔。因此,无论在区系分类上,还是生态生活型上,这些特有属都是比较年轻的类群[27]。
3.2 区系地理联系
蒙古高原种子植物区系与世界大部分地区均有一定联系,尤与北温带的关系最为密切,如前文所述,北温带成分不仅广布于蒙古高原各地区,而且大多数北温带成分的乔木和灌木能够形成森林植被和灌丛植被的优势成分或建群成分,表现了北温带区系与蒙古高原区系有着非常密切的历史渊源。其次是与地中海、西亚至中亚区系之间的联系,蒙古高原属于与地中海、西亚和中亚相联系的属有137属,占本区温带性质成分的21.7%,这些地中海、西亚和中亚的成分是地中海脱离海侵局面之后,第三纪的热带成分、东亚成分、温带成分逐步向西北迁移与演变以及适应地中海区气候旱化发生蜕变和演化而来的,这与地中海孑遗成分有着共同的渊源。再次是与东亚及北美大陆温带区系之间的联系,共有25属,这些成分可能与第三纪晚期至第四纪气候变冷之后的温带植物南移与演变以及间冰期植物的南北、东西移动有着密切的联系。最后是与北极的联系,蒙古高原与北极相联系的有22属,这些成分与在受第四纪多次冰期影响的气候变化条件下的植物进退有着密切关系。关于蒙古高原与热带亚洲、热带非洲、热带大洋洲和热带美洲一些地区存在微弱的联系,蒙古高原的热带成分属的比例为7.2%,提供了蒙古高原植物区系在发展历史上与热带区系相联系的遗迹和证据。
3.3 起源与演化
蒙古高原的现代种子植物区系是古生物区系的延续和残遗,蒙古高原大部分地区曾被Tethys海所占据,经历了始新世中期的喜马拉雅造山运动后,大陆性气候愈加强烈[29],至此,许多成分便在干旱的古地中海沿岸或遗迹上发生。如分布于蒙古高原中央戈壁区的麻黄属(Ephedra)植物-膜果麻黄(Ephedraprzewalskii)在三叠纪时便发源于古地中海沿岸[30]。白垩纪尤其是老第三纪孑遗种或者是古老的或分类上孤立的残遗植物如绵刺(Potaniniamongolica)、沙冬青(Ammopiptanthusmongolicus)等均是起源于古地中海沿岸或遗迹上[31]。本区分布的古地中海孑遗植物如沙拐枣(Calligonummongolicum)、盐节木(Haloenemumstrobilaceum)、花花柴(Kareliniacaspia)、甘草(Glycyrrhizauralensis)[32]等无疑是在古地中海沿岸及遗迹上发生的。中亚也是Tethys海遗迹的一部分[33],因而起源于古地中海沿岸或遗迹上的中亚分布类型无疑也是古地中海植物区系的孑遗,或者说是一些植物种在干旱的荒漠环境中蜕变演化出来的植物种类,并不是古老区系的起源中心。
第三纪后半期,随着喜马拉雅的上升,蒙古高原脱离了海侵的局面,东亚植物区系的温带成分得以向西迁移[34],并在此独特环境中,物种的形成过程朝向旱生化发展,产生了现代地中海植物区系的基本成分,典型的代表有锦鸡儿属(Caragana)、野丁香属(Leptodermis)和罗布麻属[35]。如锦鸡儿属是发生于始新世早期亚洲东部森林成分,后沿古老的“Зысса”山系的山脊向西迁移,随着喜马拉雅和其他第三纪褶皱山系的形成,其中,迁向西北的一支向着适应干旱荒漠气候发展,产生了小叶系(Ser.Microphyllae)较进化的类型[36],其中有些种已经成为适应干旱气候的荒漠灌木[37]。又如仅局限分布于阿拉善荒漠区的绵刺作为一个蒙古高原古老的残遗旱生植物区系成分也是由东亚中生植物区系成分的银露梅(Potentillaglabra)演化而来[38]。
蒙古高原的温带成分发源于中国亚热带,亚热带常绿阔叶林成分向西北迁移,在适应本地区逐渐变旱的气候,进而蜕变发展形成适应此特殊生境的次生植物种类[33]。如沙冬青是亚洲荒漠地区仅有的常绿阔叶灌木,这应该是亚热带常绿阔叶林植物种干旱变型的有效证据[39];广布于中央戈壁的膜果麻黄也是属于老第三纪亚热带常绿植被干旱变型的残遗成分[39]。此外,北温带成分在蒙古高原大量存在,且多数的木本植物在森林和灌丛植被中,往往形成群落的优势成分或建群成分,由此可见温带成分在本区植物区系中占重要地位,反映了蒙古高原所处的地理位置(主要是纬度和海拔高度)与其气候条件相适宜的特点。
蒙古高原植物区系还有部分是从本地的荒漠环境中发展起来的,如蒺藜科(Zygophyllaceae)的霸王属(Zygophyllum)、骆驼蓬属(Peganum),藜科(Chenopodiaceae)的盐爪爪属(Kalidium)、盐穗木属(Halostachys)、猪毛菜属,石竹科(Caryophyllaceae)的刺叶属(Acanthophyllum),菊科(Compositae)的刺头菊属(Cousinia)、粉苞苣属(Chondrilla),百合科(Liliaceae)的郁金香属(Tulipa)及蓼科(Polygonaceae)的沙拐枣属(Calligonum)和木蓼属(Atraphaxis)等是在干旱的荒漠环境中形成的[39]。蒙古高原特有属中的新生进化成分亦是如此,因为它们并不分布到蒙古高原以外的地方,它们是蒙古高原植物区系的一部分。
4 结论
蒙古高原处于北温带半干旱和干旱的内陆地区,自然环境较为严酷,种子植物仅115科761属4 280种,占世界植物同类群总种数的1.9%,植物区系较贫乏,特有程度较低。蒙古高原所处的地理位置决定了其区域植物具有典型的温带性质,其区系地理成分复杂,联系广泛,一定程度上亦受到热带成分的影响,与热带成分具有一定的亲缘关系。植物区系优势现象不显著,以单型属和少型属为主。区域内植被特点是以旱生和超旱生的灌木、小灌木植物占优势,其中有很多是古老的或分类上孤立的残遗植物,使得蒙古高原植物区系表现出强烈的旱生性和古老性。从植物区系发生角度分析,蒙古高原植物区系的起源可归结为古地中海成分、东亚成分、温带成分和本地起源。
猜你喜欢
数学大王·趣味逻辑(2022年8期)2022-07-10
耕作与栽培(2020年3期)2020-09-18
花卉(2020年12期)2020-01-09
世界热带农业信息(2019年11期)2019-01-05
天然产物研究与开发(2018年9期)2018-10-08
小资CHIC!ELEGANCE(2018年24期)2018-08-13
浙江农业科学(2017年5期)2017-06-21
华东师范大学学报(自然科学版)(2017年1期)2017-02-27
网络空间安全(2016年3期)2016-06-15
安徽农业科学(2015年10期)2015-02-27
