
miRNA-155在食管癌中的表达及与其临床病理特征的关系
2018-07-12 01:16周方正潘东风张东升王小聪郭思言
实用癌症杂志 2018年6期
聂 龙 周方正 童 琳 潘东风 张东升 王小聪 谢 辉 郭思言
微小RNA(miRNA)是1种能够参与蛋白的合成以及调控机体细胞活动的小分子非编码RNA,其长度大约在21~25个核苷酸[1]。据研究报道,miR-155在肺癌[2],乳腺癌[3]和胰腺癌[4]等多种肿瘤中均存在高表达。但是还未见miR-155在食管癌中表达的相关研究报道。因此本研究通过采用RT-PCR方法检测miR-155在食管癌组织以及癌旁组织中的表达差异,并分析两者的表达与食管癌患者临床病理特征的相关性,进而探讨其对食管癌发生,转移的影响。
1 材料与方法
1.1 一般资料
选取2014年5月至2016年6月本院食管癌手术切除标本及对应的癌旁组织各80例,其中男性52例,女性28例,平均年龄42~76岁,所有患者术前均未接受化疗,放疗等其他疗法,所有组织标本分别于癌旁组织4 cm以内、无坏死癌灶及正常黏膜组织处取材。组织经4%多聚甲醛固定,常规脱水,包埋后连续切片,切片厚度控制在4 μm,进行苏木精-伊红染色(HE)染色,经专业病理科老师诊断确诊为食管癌。按WHO分类标准(1996)进行组织学分级:高分化34例,中分化25例,低分化21例。淋巴结转移者38例,无淋巴结转移者42例。标本采集经本院医院伦理委员会的审批和认可。
1.2 方法
1.2.1主要试剂Trizol Reagent购自Invitrogen公司,逆转录试剂盒和cDNA扩增试剂盒均购自MBI Fermentas公司。BioRAD-IQ5 qRT-PCR仪购自美国BioRad公司。
1.2.2RT-PCR检测组织中miR-155的表达取对数生长期细胞,采用Trizol试剂一步法提取细胞中的RNA,将细胞RNA反转录成cDNA,然后扩增成DNA(所有步骤严格按照逆转录试剂盒和扩增试剂盒说明书操作)。miR-155引物序列:上游:5-CTGTATCAAAAGGCCAACTGAA-3,下游:5-GTGTCTATCCTTATGAATCGCCA-3;以GAPDH作为内参,引物序列:上游:5-CATGGGGTGTGAACCATGAGA-3;下游:5-GTCTTCTGGGTGGCAGTGAT-3。所有引物序列均由上海生物工程技术有限公司合成。RT-PCR反应条件如下:95 ℃预变性10 min,然后95 ℃变形30 s,50 ℃退火30 s,70 ℃延伸1 min,40个循环,最后70 ℃延伸10 min结束。扩增产物经1%琼脂糖凝胶电泳,凝胶成像分析系统(Bio-Rad,USA)测条带灰度值。
1.2.3miR-155表达水平的计算方法以目的基因miR-155与内参基因GAPDH灰度值的比值反映miR-155的相对表达量。至少选取3例数据求取平均值。取食管癌患者癌旁正常组织miR-155的表达水平为1,以此为对照,计算食管癌患者癌组织中miR-155的表达水平(以倍数表示)[5]。
1.3 统计学方法
应用SPSS 22.0统计分析软件对数据进行统计分析。miR-155表达与食管癌患者临床病理特征之间的关系采用Mann-Whitney检验,计量资料用均数±标准差表示,组间的比较采用t检验,以P<0.05为差异具有统计学意义。
2 结果
2.1 miR-155在食管癌组织与癌旁组织中的表达
RT-PCR结果显示,食管癌组织中miR-155的表达水平为0.437±0.091,与癌旁组织相比差异具有统计学意义(t=25.184,P=0.007),见图1。
2.2 miR-155表达与食管癌临床病理特征的关系
食管癌组织中miR-155表达与患者性别和年龄无关(P>0.05);与食管癌组织学分级,肿瘤浸润深度,淋巴结转移和TNM分期均有一定的关系(P<0.05),见表1。

1为食管癌组织;2为癌旁组织
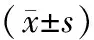
表1 miR-155表达与食管癌临床病理特征的关系
3 讨论
Has-miR-155是非编码基因BIC(B-cell integration cluster,BIC)转录所形成。近年的研究更多的聚焦于机体炎症反应调控以及血液性肿瘤的研究,但是关于miR-155与实体肿瘤的相关性研究少见报道。本研究结果显示,食管癌患者组织中miR-155的表达水平显著低于对应的癌旁组织,并且差异具有统计学意义。同时对80例食管癌组织中miR-155的表达水平与患者临床病理之间的相关性进行分析,结果发现食管癌组织中miR-155的表达与患者的性别和年龄无关(P>0.05);与患者食管癌的组织学分级,肿瘤的浸润深度,淋巴结转移和TNM分期均有一定的关系(P<0.05)。
有研究发现miR-155在肺癌[6],胃癌[7]和乳腺癌[8]等实体肿瘤中呈现高表达,并且与肿瘤的发生,发展密切相关。同时报道miR-155的高表达与其抑癌作用存在一定的关系。Dorsett等[9]研究发现,高表达的miR-155通过抑制AID(activation-induced cytidine deaminase)产生的潜在癌基因转位来发挥抗癌的作用。同期研究中也有报道miR-155高表达与促癌作用存在一定的联系。Ovcharenko等[10]研究发现,高表达的miR-155可以通过抑制凋亡基因caspase-3的表达来抑制肿瘤的凋亡,从而促进肿瘤细胞的生长。Valeri等[11]研究发现在结肠癌患者组织中miR-155的表达水平与错配修复蛋白MLH1或MSH2表达呈现负相关,结果提示miR-155可以通过调控错配修复蛋白MLH1或MSH2表达来促进结肠癌肿瘤的发生与发展。Pedersen等[12]在造血细胞增殖机制的研究发现,SHIP1是调节造血细胞增殖与分化的因子,而miR-155可以作用了靶基因SHIP1进而发挥促进细胞生长的作用。Jiang等[13]在乳腺癌细胞中研究发现miR-155可以通过抑制细胞因子信号转导蛋白1来促进乳腺癌细胞的增殖与生长。
近年的研究也证实,miR-155在炎症反应和免疫中发挥中重要的作用。miR-155可以通过上调炎性T细胞的活性来促进炎症反应的发生[14]。Vigorito等[15]研究也发现miR-155在淋巴细胞免疫反应中发挥着关键性的作用。另外,miR-155基因敲除的老鼠细胞因子的分泌降低并且T细胞依赖的抗体反应减弱[16]。而炎症细胞和炎症反应在肿瘤的发生发展中具有重要作用。因此我们推测食管癌患者组织中miR-155表达的降低可能是导致机体炎症反应以及抗癌机制的失衡,从而促进肿瘤细胞的生长。
综上,食管癌患者组织中miR-155的相对表达降低,可能抑制了机体抗癌作用,从而促进食管癌的发生与发展,但是具体的作用机制还需要进一步的研究。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30
保健医苑(2021年7期)2021-08-13
中国毕业后医学教育(2020年5期)2020-12-06
天津医科大学学报(2019年6期)2019-08-13
现代检验医学杂志(2016年2期)2016-11-14
中华老年多器官疾病杂志(2016年9期)2016-04-28
西南军医(2016年1期)2016-01-23
中国继续医学教育(2015年4期)2016-01-07
川北医学院学报(2015年5期)2015-12-05
现代检验医学杂志(2015年6期)2015-02-06
