
黄浦江浮游动物群落结构及其与环境因子的关系
2018-07-04 01:59:46吴晓敏郝瑞娟潘宏博魏华王丽卿
生态环境学报 2018年6期
吴晓敏 ,郝瑞娟 ,潘宏博 ,魏华 ,王丽卿 *
浮游动物是一类小型的水生生物,作为水域生态系统的重要组成部分,其在水域生态系统的物质循环、能量流动和信息传递中都起着十分重要的作用(孟顺龙等,2016;郭沛涌等,2003;颜庆云等,2005;高原等,2013)。它们既可为鲢鳙鱼等滤食性鱼类提供饵料,又可控制浮游藻类的生长和繁殖,从而一定程度上缓解水体富营养化程度(刘建康,1990)。同时,研究表明浮游动物的种类、数量、多样性指数、优势种等的变化,可以指示水体的水质状况或健康水平(刘建康,2002)。因此,研究浮游动物群落的动态变化对水生态系统的结构、功能、生产力等相关工作的开展具有非常重要的意义。
黄浦江是历史上最早的人工修凿疏浚的河流之一,位于长江下游入海口的一个支流,全长 113 km,宽300~700 m,上游始于上海市青浦区的淀山湖,在下游吴淞口与长江相汇,流入东海,横穿整个城市,是上海市的重要水道。该水体不仅有农田灌溉、船舶运输的功能,还有旅游观光、排污通道等功能,因此黄浦江的生态健康受到人们的广泛关注(吴波等,2007)。
目前有一些学者对黄浦江水体的鱼类、有机农药、浮游植物等进行了研究(吴波等,2007;吴波,2006;陈小华等,2008;夏凡等,2006),还有一些学者对黄浦江支流苏州河(顾笑迎等,2008;刘冬燕等,2009;俞秋佳等,2014;汪飞等,2007)以及一些长江口的浮游动植物进行过报道(郭沛涌等,2008;徐兆礼等,2005;纪焕红等,2006),这些报道显示枯水期与丰水期浮游动物群落结构在密度、生物量、种类数有很大的差异。
然而,目前还没有学者全面地对黄浦江的浮游动物群落结构及其与环境因子的关系进行过相关研究。为填补黄浦江浮游动物群落结构研究的不足,了解枯水期、丰水期的浮游动物群落结构,于2013年11月(枯水期)与2014年7月(丰水期)对黄浦江水域11个断面(共82个采样点)浮游动物物种多样性和现存量在枯、丰水期的变化进行调查研究,运用多元统计方法中的聚类分析(Cluster analysis)对黄浦江上游支流、上游干流、中游及下游的各个采样断面进行空间分布特征分析,探究黄浦江水域浮游动物群落结构特征与水体环境因子的关系,以期为黄浦江的水环境评价和生态保护提供基础资料。
1 材料与方法
1.1 采样点的设置及采样时间
本研究于2013年11月枯水期和2014年7月丰水期对黄浦江浮游动物进行两次采样。根据黄浦江的水文特征分区、河岸邻近的陆域土地利用状况差异分区以及水体功能分区等因素,从其上游到下游共设置11个采样断面(每个断面3~5个采样点,共41个采样点),断面点(St1-St11)分布见图1。其中,上游支流为 St1-St3,分别为夏字圩、三角渡、泖港大桥,上游干流为St4和St5,分别为松浦大桥、奉浦大桥;中游为St6-St8,分别为吴泾、长桥水桥、南市水厂;下游为St9-St11,分别为杨浦水厂、共青森林公园、吴淞口。St1、St2周边用地为农业用地和居民用地;St3为农业用地、居民用地、工业用地和裸地;St4、St6周边用地为农业用地、居民用地、裸地;St5、St7、St8、St9周边用地为居民用地;St10和St11周边用地为居民用地和裸地。
1.2 水质理化指标的测定
水温(Water temperature,t)、溶解氧(Dissolved oxygen,DO)、电导率(Conductivity,Spc)、盐度(Salinity,Sal)、pH、氨氮(Ammonia nitrogen,NH4+-N)和浊度(Turbidity,Turb)等水质理化指标用便携式YSI Proplus(美国)现场测定,透明度(Transparency,SD)用透明度盘现场测定。用5 L有机玻璃采水器采集混合水样,取1 L混合水样于聚乙烯瓶中,并置于4 ℃冷藏箱中,4 h内测定总氮(Total nitrogen,TN)(国标法:GB/T11894—1989)、总磷(Total phosphorus,TP)(国标法:GB/T11893—1989)、高锰酸盐指数(Permanganate index,CODMn)(国标:GB/T11892—1989)和叶绿素 a(Chlorophyll a,Chl-a)(丙酮萃取分光光度法)等水质理化指标,具体测定方法参照(国家环境保护总局,2002)。

图1 黄浦江采样断面图Fig. 1 Sampling section in Huangpu River
1.3 样品采集和处理
1.3.1 定性标本
定性样品用25号浮游生物网(孔径64 μm)在水面下0~0.5 m水层作“∞”字拖取5 min进行采集,然后将采集的样品装入样品瓶中,经福尔马林固定后在Olympus BX53显微镜下镜检观察鉴定(张武昌等,2012;诸葛燕,1997;韩茂森等,1995)。
1.3.2 定量样品
浮游甲壳动物:在各个采样断面,用规格为5 L的有机玻璃采水器在不同深度共采集水样20 L,然后用13号浮游生物网过滤到500 mL样品瓶中,按照《淡水浮游生物研究方法》(章宗涉等,1991)进行室内固定浓缩,最后处理好的样品在显微镜下进行全部计数。生物量根据体长-体重回归方程计算(黄祥飞等,1986;陈雪梅,1981)。
原生动物和轮虫:轮虫和原生动物定量样品用5 L有机玻璃采水器在水面下0.5 m和1.5 m处水层各采集5 L,等量混合后取1 L,用福尔马林及鲁哥氏碘液固定,静置48 h后浓缩至50 mL。吸取1 mL浓缩匀液至1 mL计数框,在10×10倍显微镜下进行全片镜检(设3次平行),观察并记录轮虫数量;吸取0.1 mL浓缩匀液至0.1 mL计数框进行全片镜检计数,观察并记录原生动物数量。原生动物、轮虫生物量采用体积法进行统计。
1.4 数据统计和分析
1.4.1 数据统计
本研究采用优势度指数(Y)(徐兆礼等,1989)、Shannon-Wiener多样性指数(H′)(Wilhm,1968)、Margalef丰富度指数(D)(Margalef,1958)和Pielou均匀度指数(J′)(Pielou,1966)分析黄浦江浮游动物群落结构的多样性,计算公式如下:
优势度(Y):

Shannon-Wiener多样性指数(H′):

Margalef丰富度指数(D):

Pielou均匀度指数(J′):

式中,Pi=Ni/N,以优势度指数 Y≥0.02作为优势种;Ni为浮游动物群落中第 i种物种的密度;N为群落中物种的总密度;fi代表本次样品采集中第i种物种在采样断面出现的频率;S代表总种类数。
1.4.2 数据分析软件
采用Origin 2016绘制统计图;采用IBM SPSS Statistics 23软件进行Pearson相关性分析(Pearson correlation analysis);采用PRIMER 6软件对黄浦江的浮游动物生物量进行 Log(X+1)转化并进行聚类分析(Cluster analysis),建立树状图。
2 结果
2.1 水体环境因子
黄浦江水域营养盐(氮、磷)浓度总体偏高,pH显示黄浦江水域呈弱碱性。枯水期各站点间水温变化幅度较小,在15.3~20.42 ℃之间变化,平均温度为17.42 ℃;DO平均值为3.95 mg·L-1,变化范围为 2.36~4.63 mg·L-1;NH4+-N 变化范围为 0.38~0.73 mg·L-1,平均为 0.62 mg·L-1;SD 平均为 32.17 cm,变化范围为 22~40 cm;TN 变化范围为 3.27~8.30 mg·L-1,平均为 5.19 mg·L-1;Chl-a 平均为 1.97 μg·L-1,变化范围为 0.93~3.13 μg·L-1。丰水期各站点间的平均水温为 25.92 ℃,变化范围为 25.30~26.58 ℃;DO 平均值为 2.11 mg·L-1,变化范围为 0.76~3.65 mg·L-1;NH4+-N 变化范围为 0.34~0.97 mg·L-1,平均为 0.50 mg·L-1;SD 平均为 39.10 cm,变化范围为32.00~43.00 cm;TN 变化范围为 2.86~5.08 mg·L-1,平均为 3.48 mg·L-1;Chl-a 平均为 5.44 μg·L-1,变化范围为 2.83~11.44 μg·L-1(图 2)。
2.2 浮游动物群落结构的动态变化
2.2.1 浮游动物种类数
调查共检出109种浮游动物,其中原生动物35种,轮虫33种,枝角类18种,桡足类23种。就时间格局而言,枯水期共检出浮游动物 74种,其中原生动物、轮虫、枝角类和桡足类分别有21、19、13和21种,分别占总种类数的28%、26%、18%和28%。丰水期共检出79种,其中原生动物、轮虫、枝角类和桡足类分别有25、25、13和16种,分别占总种类数的32%、32%、16%和20%。
2.2.2 浮游动物优势种
经两次调查,共发现优势种 20种(浮游甲壳类12种,轮虫4种,原生动物4种),各个类群的第一优势种分别为桡足幼体(Copepodid)、脆弱象鼻溞(Bosmina fatalis)、广布多肢轮虫(Polyarthra vulgaris)和淡水麻铃虫(Leprotintinnus fluviatile)。其中,枯水期优势种13种,丰水期优势种18种。两个时期共有优势种11种,包括浮游甲壳类6种,轮虫1种,原生动物4种(表1)。

图2 黄浦江枯水期(11月)和丰水期(7月)环境因子的变化Fig. 2 The variation of environmental factors in the Huangpu River during dry season (November ) and wet season (July)
丰水期与枯水期的优势种略有差异,如:长肢秀体溞(Diaphanosoma leuchtenbergianum)仅在枯水期成为优势种;而等刺温剑水蚤(Thermocyclops kawamurai)、多刺裸腹溞(Moina macrocopa)、角突网纹溞(Ceriodaphnia cornuta)、多刺秀体溞(Diaphanosoma sarsi)、长三肢轮虫(Filinia longisela)、顶生三肢轮虫(Filinia major)、角突臂尾轮虫(Brachionus angularis)等7个种仅在丰水期成为优势种(表1)。

表1 黄浦江浮游动物优势种的密度及优势度Table 1 Dominant species abundances and dominance of zooplankton in Huangpu River
2.2.3 浮游动物密度的时空变化
黄浦江浮游动物两次采样各采样断面的密度变化范围为 1671.8~70428.4 ind.·L-1,总平均密度为17931.7 ind.·L-1,以原生动物为主。
黄浦江水域浮游动物密度的时空分布为:枯水期浮游动物平均密度为12817.1 ind.·L-1(变化范围为 1671.8~37889.0 ind.·L-1),以原生动物为主,平均密度为 12726.5 ind.·L-1,占总平均密度的99.29%。丰水期浮游动物平均密度为 23046.3 ind.·L-1(变化范围为 2074.2~70428.4 ind.·L-1),原生动物平均密度为 22794.7 ind.·L-1,占总密度的98.91%(图3)。
2.2.4 浮游动物生物量的时空变化
黄浦江两次采样各采样断面的生物量(湿重)变化范围为0.26~14.97 mg·L-1,平均生物量为3.54 mg·L-1,以原生动物为主。
黄浦江水域浮游动物生物量的时空分布为:枯水期浮游动物的平均生物量为2.36 mg·L-1,生物量比较大的种类有:淡水麻铃虫(Leprotintinnus fluviatile)(1.80 mg·L-1,占总生物量的76.27%)、淡水筒壳虫(Tintinnidium fluviatile)(0.13 mg·L-1)、小筒壳虫(Tintinnidium pusillum)(0.10 mg·L-1)、汤匙华哲水蚤(Sinocalanus dorrii)(0.09 mg·L-1)。丰水期浮游动物平均生物量为4.72 mg·L-1,生物量较大的种类有:淡水麻铃虫(3.65 mg·L-1,占总生物量的 77.33%),淡水筒壳虫(0.25 mg·L-1)、小筒壳虫(0.12 mg·L-1)、桡足幼体(0.11 mg·L-1)(图3)。
2.3 浮游动物多样性指数的变化
黄浦江枯、丰水期浮游动物物种丰富度指数Margalef(D)分别为2.79、3.08,根据评价标准,D值显示黄浦江水体为轻度污染或清洁水体;Shannon-Wiener多样性指数(H′)分别为 1.24、1.44,H′值表明黄浦江水体为中度污染;物种均匀度指数Pielou(J′)分别为0.39、0.43,表明黄浦江水体为中度污染。总体而言,黄浦江水体处于中度污染状态。浮游动物丰水期的多样性指数均大于枯水期,其中,枯水期浮游动物H′值变化范围为0.79~1.69;浮游动物 D值变化范围为 1.48~3.43;浮游动物 J′值变化范围为0.26~0.54。由此可判断,枯水期黄浦江已经处于中度污染,此外,J′值和H′值从黄浦江上游干流到下游整体呈下降趋势,说明上游水质高于下游水质。丰水期浮游动物 H′值变化范围为0.80~2.26;浮游动物D值在2.38~4.72之间变化;浮游动物J′值变化范围为0.23~0.63(图4)。从两个时期浮游动物多样性指数看,黄浦江丰水期水质优于枯水期。
2.4 浮游动物群落结构的聚类分析
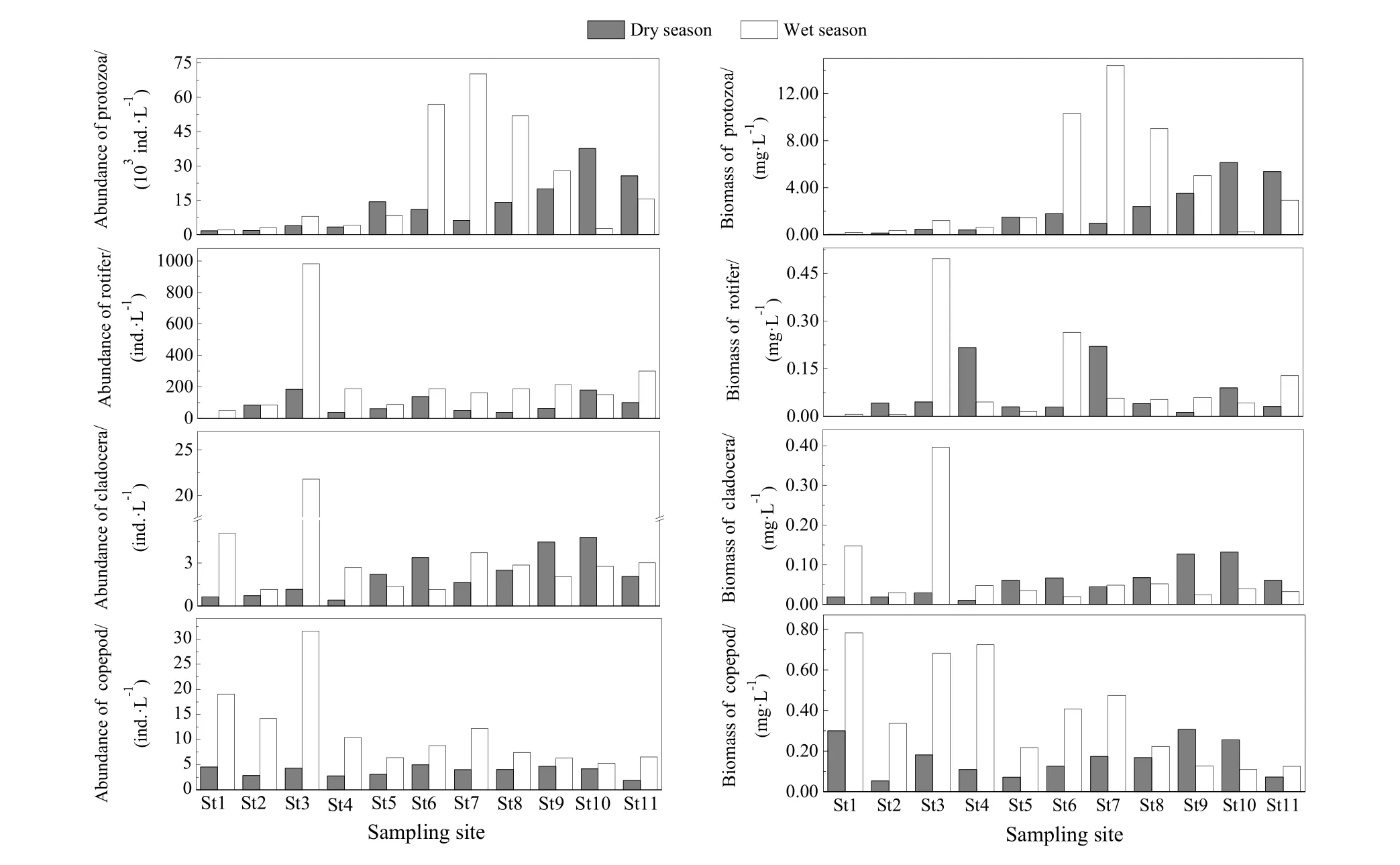
图3 浮游动物密度和生物量的时空变化Fig. 3 Temporal and spatial variation of zooplankton abundance and biomass in Huangpu River
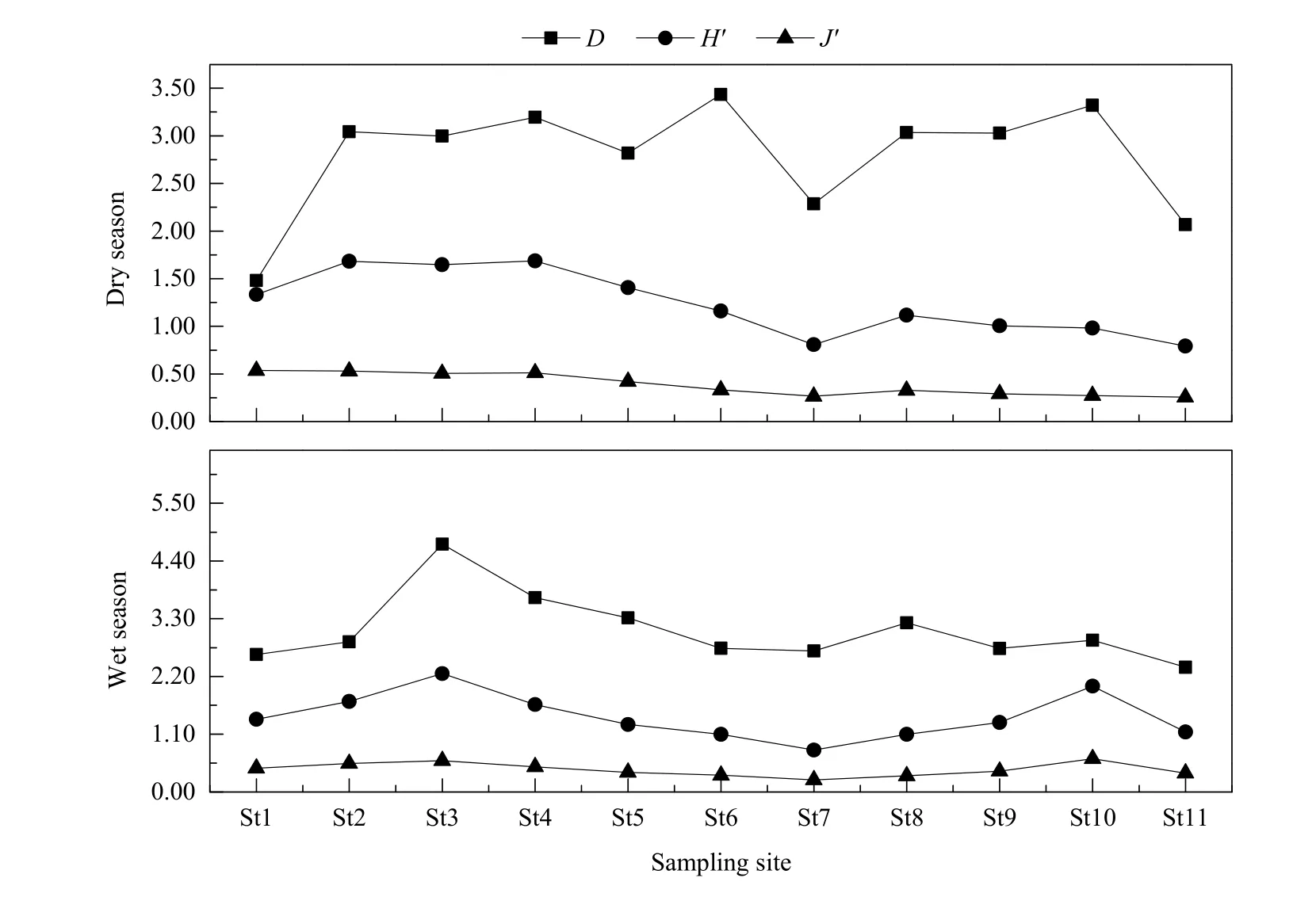
图4 浮游动物多样性指数Fig. 4 Diversity index of zooplankton
聚类分析(Cluster analysis)(图5和图6)可反映研究期间黄浦江水域各个断面浮游动物的空间分布情况。在相似系数为65%时,枯水期黄浦江水域的浮游动物群落被划分为5个类群:类群1,包含St3-St4;类群2,包含St5-St7;类群3,包含St8-St11;类群4,仅包含St1;类群5,仅包含St2。这表明,枯水期黄浦江水域从上游到下游存在较大差异性,基本呈现为上游支流、上游干流及中游为一个类群,下游为一个类群。在相似系数为38%时,丰水期黄浦江水域浮游动物群落分为3个类群,类群1,包含St1-St5;类群2,包含St6-St9和St11;类群3,仅包含St10。这表明,丰水期上游支流和上游干流浮游动物群落相似性较高,中下游段浮游动物群落为一个类群。

图5 枯水期各断面浮游动物群落的聚类分析Fig. 5 The cluster analysis of zooplankton community in the dry season
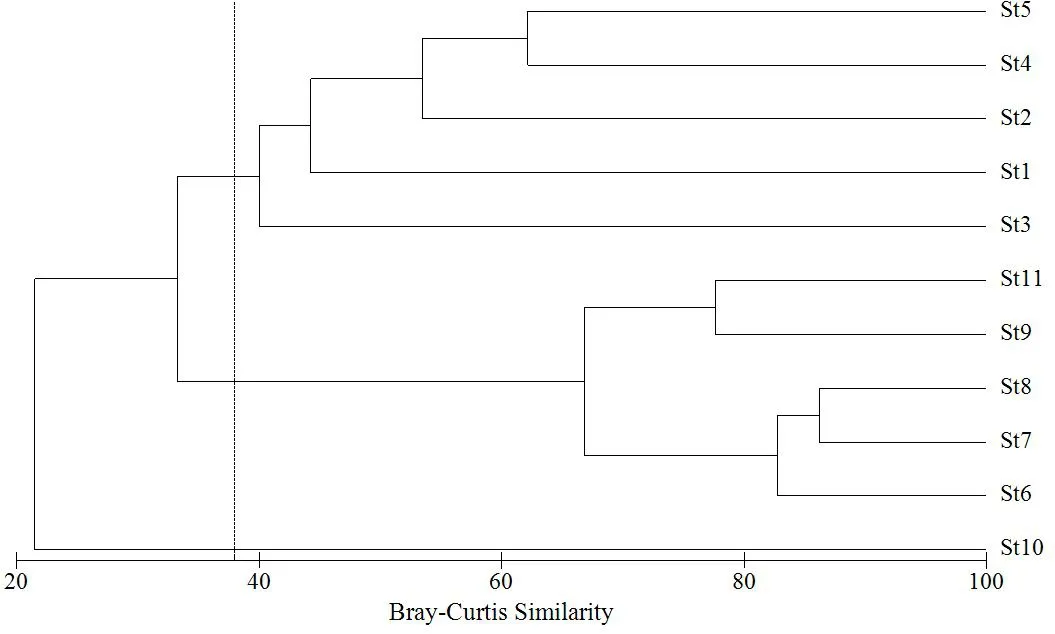
图6 丰水期各断面浮游动物群落的聚类分析Fig. 6 The cluster analysis of zooplankton community in wet season
2.5 浮游动物类群及其优势种与水体环境因子的关系
浮游动物4个类群及其优势种与水体环境因子的Pearson相关性分析表明,轮虫、枝角类与桡足类均与叶绿素呈极显著正相关,此外,轮虫(Rotifer)与总磷呈显著正相关,桡足类(Copepods)与水温呈极显著正相关,枝角类(Cladocera)与氨氮呈显著正相关;原生动物(Protozoa)与氨氮呈显著负相关;淡水麻铃虫(Leprotintinnus fluviatile)密度与溶解氧和氨氮均呈显著负相关;淡水筒壳虫(Tintinnidium fluviatile)与溶解氧呈极显著负相关,与氨氮呈显著负相关,与总磷呈显著正相关;广布多肢轮虫(Polyarthra vulgaris)与总磷和叶绿素a均呈极显著正相关,与溶解氧呈显著负相关;脆弱象鼻溞(Bosmina fatalis)与总氮呈极显著正相关;微型裸腹溞(Moina micrura)、桡足幼体(Copepodite)、无节幼体(Nauplii)、台湾温剑水蚤(Thermocyclops taihokuensis)均与叶绿素呈极显著正相关,其中微型裸腹溞与氨氮呈显著正相关,无节幼体、台湾温剑水蚤还与水温呈极显著正相关,桡足幼体与水温呈显著正相关;球状许水蚤(Schmackeria forbesi)与水温和 pH均呈显著正相关;此外,无节幼体、台湾温剑水蚤和桡足类均与浊度呈极显著或显著负相关(表2)。
3 讨论
3.1 黄浦江浮游动物群落分布特征
黄浦江枯水期及丰水期的浮游动物的种类数及密度均以小型的原生动物和轮虫为主,而后生的枝角类、桡足类所占的比例较小。本研究与其他淡水水体如惠州西湖、晋江流域、武汉东湖、黑龙江水系等的浮游动物(陈光荣等,2008;谢进金等,2005;杨宇峰等,1994;姜作发等,2006)的调查结果大体相同。值得注意的是,黄浦江浮游动物种类数显著高于松花江哈尔滨段(40种)、瓯江口水域春夏季水域(42种)和浅水型亚热带河口(54种)等水体中的浮游动物种类数(鞠永富等,2017;张亚洲等,2016;Nicola et al.,2015),这可能与黄浦江的St1-St10站点均远离长江口的入海口处,水体较稳定有关。然而,本研究中原生动物和轮虫的种类数均应高于实际的调查结果,这与采样方法中对原生动物和轮虫进行了固定导致收缩后的种类难于准确鉴定有关。生物量以原生动物为主,其次是桡足类,这主要是由于原生动物生长周期短、繁殖快,其密度大,而桡足类由于个体大,因而平均生物量较大;此外鱼类对浮游动物进行摄食时,更偏好体型较大的浮游甲壳动物(Hall et al.,1976),这进一步造成黄浦江浮游动物以小型的原生动物和轮虫为主。

表2 不同浮游动物类群、优势种密度与环境因子的Pearson相关性总结Table 2 Pearson correlations between different groups and dominant species abundances of zooplankton with environmental factors in the Huangpu River
黄浦江丰水期浮游动物总密度及原生动物、轮虫、枝角类和桡足类各类群的密度均高于枯水期,且都高出将近2倍。对浮游动物生物量而言,丰水期浮游动物生物量同样约是枯水期的2倍多。浮游动物群落的这一特征正好反映了其对不同时期水质环境变化的适应性。温度是影响浮游动物的生长、发育、群落组成等极为重要的环境因子(陈光荣等,2008;Wang et al.,2004;Yu et al.,2008),而本研究中,丰水期与枯水期的温度有显著的差异,造成浮游动物数量变化的主要原因可能是温度。此外,黄浦江的轮虫、枝角类、桡足类均与叶绿素呈极显著正相关(表 2),而丰水期的叶绿素含量显著高于枯水期(图 2),叶绿素含量代表着水体的初级生产力,在一定程度上能反映藻类的密度和生物量。相关研究表明,浮游植物的密度与浮游动物群落结构密切相关(王庆等,2010),蔡庆华(1995)、陈光荣等(2010)研究也表明叶绿素a的含量与浮游动物密度和生物量有关。这也导致丰水期浮游动物密度和生物量较高。郭沛涌等(2008)研究表明,在枯水期,长江径流量少,河口区浮游动物种类、数量均较少。相反,由于丰水期黄浦江水域随着降水量的增多,也使得其水系连通程度增加,而水系连通性的提高,客观上为浮游动物的生活环境提供了多样性,使得浮游动物多样性得以提高。本研究原生动物密度与氨氮呈显著负相关(表 2),且这一负相关性主要体现在丰水期St6-St8(图2和图3),这可能是由于该段水域属于生活污染区,排放的氨氮较多,原生动物的密度很大,其大量繁殖极大地消耗了该居民区排放的氨氮。
广生性的广布多肢轮虫与叶绿素和总磷呈极显著正相关,这表明广布多肢轮虫更加适应叶绿素含量高、总磷较高的环境,这可能与夏秋季藻类在水体中含量丰富,为轮虫在夏秋季繁殖提供了丰富的食物来源有关(王庆等,2010)。此外,本研究中,微型裸腹溞、桡足幼体、无节幼体和台湾温剑水蚤等浮游甲壳动物与叶绿素呈极显著正相关,且无节幼体、台湾温剑水蚤和桡足幼体均与水温呈极显著正相关或显著正相关,这可能是由于温度升高(袁林等,2013)导致水体中浮游藻类大量繁殖,叶绿素 a浓度升高,给浮游甲壳动物创造了广泛的食物来源,使得其在夏秋季成为优势种。
3.2 黄浦江周边用地类型对浮游动物群落分布的影响
有学者指出,若流域内的周边用地为居民用地、工业用地、商业用地为主时,径流主要污染物为总氮、总磷等营养盐(Ren et al.,2003)。流域尺度上的土地利用类型和空间分布影响水质的污染状况(Schoonover et al.,2006;Hwang et al.,2007),而营养盐的含量直接影响浮游动物群落结构。黄浦江各个断面的周边用地类型以及其水质空间分布均具有异质性(李典宝等,2015),而丰水期雨量较大,水量充沛,丰富的水量使地表径流增加,导致大量周边用地的营养盐等陆源物质进入黄浦江径流,加剧水质空间分布的异质性。本研究对丰水期及枯水期的群落结构类群的划分,与所报道出的周边用地类型(李典宝等,2015)密切相关。其中,丰水期的浮游动物结构基本表现为上游支流及上游干流为一个类群(包括St1、St2、St3、St4、St5),主要是由于这几个站点均为农业用地、裸地或居民用地等污染较小的站点,对浮游动物的群落影响较相似而被聚为一类;而中下游段(包括St6-St9和St11)的浮游动物群落结构相似性较高,可能与这几个站点的周边大多为居民用地,使得流入这几个站点的营养盐等陆源物质类型相似有关。此外,St11附近有部分裸地,且其处于入海口处,使得该站点的水体易受海水的扰动并与中游流下的水流进行汇合,因此,中下游段几个站点的浮游动物群落结构相似性较高。枯水期浮游动物群落基本呈现为上游支流、上游干流及中游(包括St3-St7)几个站点的群落相似性较高,下游(St9-St11)浮游动物群落相似性较高,这可能与枯水期雨量较小,地表径流较小,浮游动物群落受周边用地类型的影响也相对较小有关。
3.3 黄浦江浮游动物优势种对水质的指示作用
一般情况下,浮游动物在清洁型的水体中的主要的特点表现为种类多、数量少;在富营养化的水体中一些浮游动物耐污种会形成优势种群;而几乎所有水生生物在重度污染的水体都难以生存(陈光荣等,2008)。本研究中,淡水麻铃虫与水体中的氨氮浓度呈显著负相关,这可能与砂壳纤毛虫的丰度以及群落结构易受水体中氮浓度的影响有关(于凌宇等,2014)。本研究中,广布多肢轮虫与水体中的总磷浓度和叶绿素a含量均呈极显著正相关,而王庆等(2010)对南方某水库的研究表明,广布多肢轮虫在调查期间各月的相对丰度都比较高,但其与水体环境因子的相关性不大。此外,在本研究中,脆弱象鼻溞和球状许水蚤等寡污带物种(李强等,2015)在丰、枯水期均成为优势种,可能与黄浦江的水质状况较好有关。已有学者提出(刘歆璞等,2013),桡足类的剑水蚤(本研究丰、枯水期的优势种为台湾温剑水蚤)适宜生活在中度污染水体中,而本次调查期间黄浦江的水体正处于中度污染状态。Pearson相关性分析表明,淡水麻铃虫、淡水筒壳虫、广布多肢轮虫、脆弱象鼻溞、微型裸腹溞、桡足幼体、无节幼体、台湾温剑水蚤和球状许水蚤等9个种的存在与否以及是否成为调查水体的优势种可以作为评定黄浦江等河流水体富营养化水平高低及污染程度轻重的指示物种,但这一结论仍有待进一步实验验证。
4 结论
黄浦江丰水期浮游动物总密度及原生动物、轮虫、枝角类、桡足类等各类群密度均高于枯水期,且都高出接近2倍;浮游动物丰水期的生物量同样约为枯水期的2倍。多样性指数显示黄浦江丰水期的水质优于枯水期。
黄浦江的浮游动物群落结构与采样点周边用地类型以及地表径流密切相关,主要表现为:丰水期的浮游动物群落基本呈现为上游支流及上游干流段一个类群、中下游段为一个类群,而枯水期的浮游动物群落基本呈现为上游支流、上游干流及中游段为一个类群,下游段为一个类群。
淡水麻铃虫、淡水筒壳虫、广布多肢轮虫、脆弱象鼻溞、微型裸腹溞、桡足幼体、无节幼体、台湾温剑水蚤和球状许水蚤等9个种的存在与否以及是否成为调查水体的优势种可以作为评定黄浦江等河流水体富营养化水平高低及污染程度轻重的指示物种,但这一结论仍有待进一步实验验证。
HALL D J, THRELKELD S T, BURNS C W, et al. 1976. The size-efficiency hypothesis and the size structure of zooplankton communities [J]. Annual Review of Ecology and Systematics, 7:177-208.
HWANG S J, LEE S W, SON J Y, et al. 2007. Moderating effects of the geometry of reservoirs on the relation between urban land use and water quality [J]. Landscape and Urban Planning, 82(4): 175-183.
MARGALEF R. 1958. Information theory in ecology [J]. International Journal of General Systems, 3: 36-71.
NICOLA K C, RENZO P. 2015. Zooplankton community structure during a transition from dry to wet state in a shallow, subtropical estuarine lake[J]. Continental Shelf Research, 111(Part B): 294-303.
PIELOU E C. 1966. Species-diversity and pattern-diversity in the study of ecological succession [J]. Journal of Theoretical Biology, 10(2):370-383.
REN W W, ZHONG Y, MELIGRANA J, et al. 2003. Urbanization, land use,and water quality in Shanghai 1947—1996 [J]. Environment International,29(5): 649-659.
SCHOONOVER J E, LOCKABY B G. 2006. Land cover impacts on stream nutrients and fecal coliform in the lower Piedmont of West Georgia [J].Journal of Hydrology, 331(3-4): 371-382.
WANG J, XIE P, TAKAMURA N, et al. 2004. The picophytoplankton in three Chinese lakes of different trophic status and its relationship to fish populations [J]. Journal of Freshwater Ecology, 19(2): 285-293.
WILHM J L. 1968. Use of biomass units in Shannon's Formula [J]. Ecology,49(1): 153-156.
YU Y H, YAN Q Y, FENG W S. 2008. Spatiotemporal heterogeneity of plankton communities in Lake Donghu, China, as revealed by PCR-denaturing gradient gel electrophoresis and its relation to biotic and abiotic factors [J]. Fems Microbiology Ecology, 63(3):328-337.
蔡庆华. 1995. 武汉东湖浮游生物间相互关系的多元分析[J]. 中国科学院研究生院学报, 12(1): 97-102.
陈光荣, 雷泽湘, 谭镇, 等. 2010. 环境因子对广东城市湖泊后生浮游动物的影响[J]. 水生态学杂志, 3(4): 28-32.
陈光荣, 钟萍, 张修峰, 等. 2008. 惠州西湖浮游动物及其与水质的关系[J]. 湖泊科学, 20(3): 351-356.
陈小华, 李小平, 程曦. 2008. 黄浦江和苏州河上游鱼类多样性组成的时空特征[J]. 生物多样性, 16(2): 191-196.
陈雪梅. 1981. 淡水桡足类生物量的测算[J]. 水生生物学集刊, 7(3):397-408.
高原, 赖子尼, 李捷, 等. 2013. 连江浮游动物多样性的空间分布[J]. 生物多样性, 21(6): 699-708.
顾笑迎, 由文辉. 2008. 苏州河浮游植物群落结构与水质评价[J]. 海洋湖沼通报, (1): 66-73.
郭沛涌, 沈焕庭, 刘阿成, 等. 2003. 长江河口浮游动物的种类组成、群落结构及多样性[J]. 生态学报, 23(5): 892-900.
郭沛涌, 沈焕庭, 刘阿成, 等. 2008. 长江河口中小型浮游动物数量分布、变动及主要影响因素[J]. 生态学报, 28(8): 3517-3526.
国家环境保护总局. 2002. 水和废水监测分析方法[M]. 第 4版. 北京:中国环境科学出版社.
韩茂森, 束蕴芳. 1995. 中国淡水生物图谱[M]. 北京: 海洋出版社.
黄祥飞, 胡春英. 1986. 淡水常见枝角类体长-体重回归方程式[C]//甲壳动物学论文集编辑委员会. 甲壳动物学论文集. 北京: 科学出版社:147-157.
纪焕红, 叶属峰. 2006. 长江口浮游动物生态分布特征及其与环境的关系[J]. 海洋科学, 30(6): 23-30.
姜作发, 唐富江, 董崇智, 等. 2006. 黑龙江水系主要江河浮游动物种群结构特征[J]. 东北林业大学学报, 34(4): 64-66.
鞠永富, 于洪贤, 于婷. 2017. 松花江哈尔滨段浮游动物群落结构特征与水质评价[J]. 湖泊科学, 29(3): 646-653.
李典宝, 张玮, 王丽卿, 等. 2015. 2013年上海市河流秋季水质空间分布特征[J]. 生态与农村环境学报, 31(1): 50-58.
李强, 田华, 姜民, 等. 2015. 淀山湖浮游动物群落结构特征及其影响因子[J]. 水生态学杂志, 36(4): 69-77.
刘冬燕, 林文鹏, 赵敏. 2009. 苏州河浮游植物群落结构特征[J]. 长江流域资源与环境, 18(10): 914-918.
刘建康. 1990. 东湖生态学研究(一)[M]. 北京: 科学出版社.
刘建康. 2002. 高级水生生物学[M]. 北京: 科学出版社.
刘歆璞, 王丽卿, 张宁, 等. 2013. 青草沙水库后生浮游动物群落结构及其与环境因子的关系[J]. 生态学杂志, 32(5): 1238-1248.孟顺龙, 肖代, 陈小丽, 等. 2016. 丰、枯水期里下河腹地典型水体浮游动物群落结构与水质评价[J]. 水生态学杂志, 37(6): 76-83.
汪飞, 吴德意, 王灶生, 等. 2007. 以浮游生物为指示生物的苏州河生态安全评价[J]. 环境科学与技术, 30(3): 52-54, 118.
王庆, 刘丽君, 杨宇峰. 2010. 南方某水库轮虫群落特征及其对环境因子的响应[J]. 生态学报, 30(13): 3385-3395.
吴波, 陈德辉, 吴琼, 等. 2007. 黄浦江浮游植物群落结构及其对水环境的指示作用[J]. 武汉植物学研究, 25(5): 467-472.
吴波. 2006. 上海苏州河、黄浦江浮游植物群落结构及其对环境指示作用的研究[D]. 上海: 上海师范大学.
夏凡, 胡雄星, 韩中豪, 等. 2006. 黄浦江表层水体中有机氯农药的分布特征[J]. 环境科学研究, 19(2): 11-15.
谢进金, 许友勤, 陈寅山, 等. 2005. 晋江流域水质污染与浮游动物四季群落结构的关系[J]. 动物学杂志, 40(5): 8-13.
徐兆礼, 陈亚瞿. 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15, 19.
徐兆礼, 沈新强. 2005. 长江口水域浮游动物生物量及其年间变化[J].
长江流域资源与环境, 14(3): 282-286.
颜庆云, 余育和, 冯伟松. 2005. 洞庭湖浮游生物群落DNA指纹拓扑结构与物种组成对应关系[J]. 生态学报, 25(9): 2221-2226.
杨宇峰,黄祥飞. 1994. 武汉东湖浮游动物群落结构的研究[J]. 应用生态学报, 5(3): 319-324.
于凌宇, 张文静, 林元烧, 等. 2014. 广西铁山港海域砂壳纤毛虫生态学研究[J]. 安全与环境工程, 21(3): 12-17, 27.
俞秋佳, 徐凤洁, 蒋跃, 等. 2014. 夏秋季苏州河浮游植物群落特征及其影响因子[J]. 生态与农村环境学报, 30(4): 450-457.
袁林, 吴惠仙, 彭自然, 等. 2013. 阳澄湖围网内外浮游甲壳动物群落结构比较[J]. 上海海洋大学学报, 22(4): 586-592.
张武昌, 丰美萍, 于莹, 等. 2012. 砂壳纤毛虫图谱[M]. 北京: 科学出版社.
张亚洲, 蒋日进, 李凯, 等. 2016. 瓯江口水域春、夏季浮游动物群落结构特征[J]. 浙江海洋学院学报(自然科学版), 53(3): 201-206.
章宗涉, 黄翔飞. 1991. 淡水浮游生物研究方法[M]. 北京: 科学出版社.
诸葛燕. 1997. 中国典型地带轮虫的研究[D]. 武汉: 中国科学院水生生物研究所.
猜你喜欢
水生态学杂志(2023年4期)2023-04-29 00:44:03
小主人报(2022年8期)2022-08-18 01:38:34
科学养鱼(2020年10期)2020-11-23 09:04:10
现代装饰(2020年4期)2020-05-20 08:56:06
课外语文·中(2020年2期)2020-03-24 22:18:09
课外语文(2020年5期)2020-03-14 08:16:40
知识窗(2019年4期)2019-04-26 03:16:02
教育教学论坛(2019年7期)2019-03-18 11:50:12
安徽农学通报(2018年19期)2018-12-25 10:57:36
水利规划与设计(2017年11期)2017-12-23 06:34:11
