
拉萨河中下游原生动物群落物种分布格局及其驱动因素
2023-04-29 00:44:03达珍张鹏安瑞志潘成梅乔楠茜巴桑
水生态学杂志 2023年4期
达珍 张鹏 安瑞志 潘成梅 乔楠茜 巴桑

摘要:了解拉萨河中下游原生动物群落物种分布格局及其驱动因素,为拉萨河中下游生物多样性保护及可持续利用提供基础资料。选取了拉萨河中下游17个样点,于2015年5月、8月和2016年11月进行原生动物定性和定量样品采集,共采集水样153份,分析物种组成、生物多样性及其与环境因子之间的关系,确定该流域原生动物类群组成中稀有种、偶见种和常见种对物种多样性的贡献、生物多样性格局及群落与理化因子相关性。结果表明:(1)3个季节共调查到稀有种80种,占总物种数的40%;偶见种95种,占48%;常见种21种,占11%;恒见种1种,占0.5%。(2)拉萨河中下游原生动物偶见种是原生动物群落物种多样性格局的重要贡献者。(3)多样性指数表明,Richness指数和Margalef指数春季和夏季高于秋季,Shannon-Wiener指数和Simpson指数春季高于秋季高于夏季。(4)基于Bray-Curtis距离的主坐标分析和相似性分析结果表明,拉萨河中下游原生动物群落结构在不同季节和不同流域均具有显著差异,且流域差异相比季节差异较大。(5)根据Mantel检验分析原生动物与环境因子的相关性,在所测定环境因子中,水温(WT)、总溶解性固体(TDS)、盐度(Salt)、电导率(EC)、总磷(TP)及溶解氧(DO)是影响原生动物群落结构的主要环境因子。
关键词:原生动物;物种多样性;环境因子;拉萨河
中图分类号:Q178.1 文献标志码:A 文章编号:1674-3075(2023)04-0010-08
原生动物是动物界中一大类单细胞真核生物的统称,其个体小,生长发育快,繁殖能力强,在水生态系统物质循环、能量传递与生物资源利用过程中发挥着十分重要的作用,其物种组成、优势种以及多样性能直接反映水环境的状况(Pawlowski,2014;安瑞志等,2021)。物种分布格局及其驱动因素一直是生态学家关注的焦点(Lennon et al,2004;王思齐和朱章明,2020)。Moura (2021)根据物种的出现频率,将物种划分为恒有种、常见种、偶见种和稀有种,其中有关稀有种、偶见种和常见种对物种多样性格局相对贡献的研究较多,主要集中于植物群落(刘旻霞等,2020)和浮游植物群落(Moura et al,2021;索雨泽等,2021),然而很少有人将此分析方法应用于原生动物群落的相关研究。
雅鲁藏布江位于青藏高原西南部,是我国水能资源最丰富的河流(彭定志等,2008)。雅鲁藏布江流域由于高寒环境、地形复杂、人口稀疏、交通不便等因素,保持着相对稳定的原始生态系统,因而备受国内外研究生物起源、进化、区系和迁徙的学者的广泛关注(Zhang et al,2018;安瑞志等,2020)。拉萨河是雅鲁藏布江五大支流之一,在调节区域气候、水资源供给和维持生物多样性等方面起着重要的生态安全屏障作用。目前,有关拉萨河的水生生物学研究主要集中在浮游植物(君珊等,2019)和原生动物(巴桑等,2017;杨欣兰等,2019)等方面,但针对拉萨河中下游原生动物的系统和持续研究仍较少。
为了解拉萨河中下游原生动物群落物种分布格局及其驱动因素,于2015年5月、8月和2016年11月对拉萨河中下游原生动物进行了定性和定量样品采集,分析其物种组成、生物多样性及其与环境因子之间的关系,确定该流域原生动物类群组成中稀有种、偶见种和常见种对物种多样性的贡献、生物多样性格局及群落与理化因子相关性,以期为拉萨河中下游生物多样性保护及可持续利用提供基础资料。
1 研究方法
1.1 区域概况
拉萨河位于青藏高原的中南部,是雅鲁藏布江长度最长、面积最大的支流,发源于念青唐古拉山南麓(安宝晟等,2021)。根据河谷形态和地质情况,分为上游、中游、下游3段(何柳,2019)。自源头至桑曲县汇入口为上游段,位于自然区域,全长256 km,气候常年寒冷干燥,空气稀薄,人口稀少,多为牧民,生态环境较为原始(刘久潭等,2020)。从桑曲到直孔为中游段,全长138 km,为高原温带半干旱季风气候,日照充足,昼夜温差大,人类活动较少(陈歆等,2019)。直孔以下为下游段,全长157 km,地势平缓,河谷宽广,相比中上游,人类活动干扰较强,沿拉萨市流域城镇数量变多,人口密集,且该流域为拉萨市区主要粮食产区(陈丽等,2019)。
1.2 样点设置
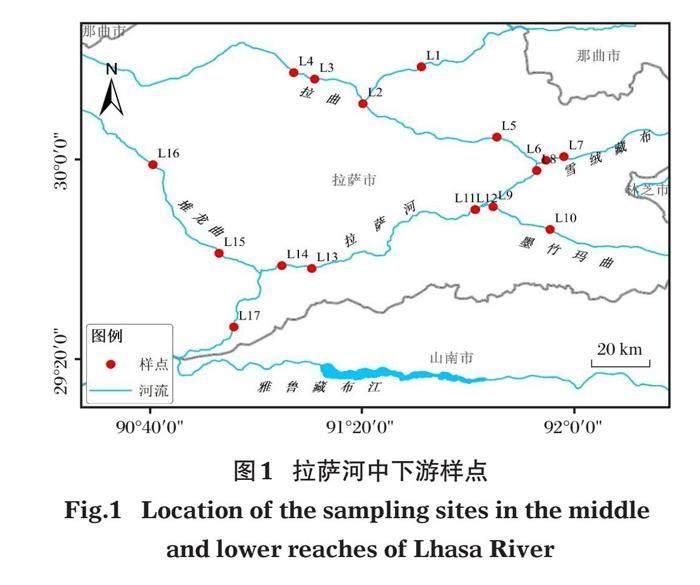
依据《水域生态系统观测规范》,在拉萨河中下游河段共计设置17个样点(图1),样点基本信息见表1。分别于2015年5月、8月和2016年11月进行采样,共采集样本153份。
1.3 水体理化因子测定
水体pH、水温(WT)、总溶解性固体(TDS)、盐度(Salt)及电导率(EC)现场用PSCTestr35防水型多参数笔测试仪(EUTECH,美国)测定;溶解氧(DO)现场用HI9146便携式溶解氧测定仪(HANNA,德国)测定;水流速度(WS)用FP111直读式流速仪(HANNA,美国)测定;浊度(Tur)用HI98703高精度数据型浊度测定仪(HANNA,意大利)测定;总氮(TN)、总磷(TP)、重铬酸盐指数(CODCr)、氨氮(NH3-N)和硝态氮(NO3-N)送检测公司测定,测定方法参照国家环境保护总局《水和废水监测分析方法》和《地表水环境质量标准》(GB3838-2002)送检测公司进行测定。
1.4 样品采集与处理
样品采集标准及实验室处理方法均参照《淡水浮游生物研究方法》(章宗涉和黄祥飞,1991)进行。定性样品用25号浮游生物网(孔径64 μm)在水面下0.8~1.2 m处作“∞”字型捞取5~10 min,将浓缩样品装入标本瓶,在室内利用Olympus CX23研究型光学显微镜,采用活体镜检和固定染色法鉴定物种(中国科学院青藏高原综合科学考察队,1981;沈韫芬,1999)。定量样品在水面下0.8~1.2 m采集原水1 L,加10 mL(10%)鲁哥氏液固定,实验室静置24 h沉淀浓缩至50 mL。移取0.1 mL浓缩液至0.1 mL计数框,在10×40倍显微镜下观察,全片平行计数3次(计数差异不超过15%)。
1.5 数据统计与分析
参照Moura(2005)计算原生动物在群落中的出现频率(FO),分为恒见种(FO≥70%)、常见种(30%≤FO<70%)、偶见种(10%≤FO<30%)和稀有种(FO<10%)。
利用Excel 2016软件进行原始数据的基础统计处理;利用R语言软件Vegan包计算Shannon-Wiener指数(H')、Margalef指数(dM)、Richness指数(R)、Simpson指数(D)、单因素方差分析(one-way ANOVA);Tidyverse、Vegan包进行Mantel检验分析;ggplot2、ggcor包进行绘图;利用Origin pro 2019分析原生动物稀有种、偶见种和常见种对群落物种多样性的相对贡献;利用ArcGIS10.7软件绘制研究区域地图。
2 结果与分析
2.1 水环境因子
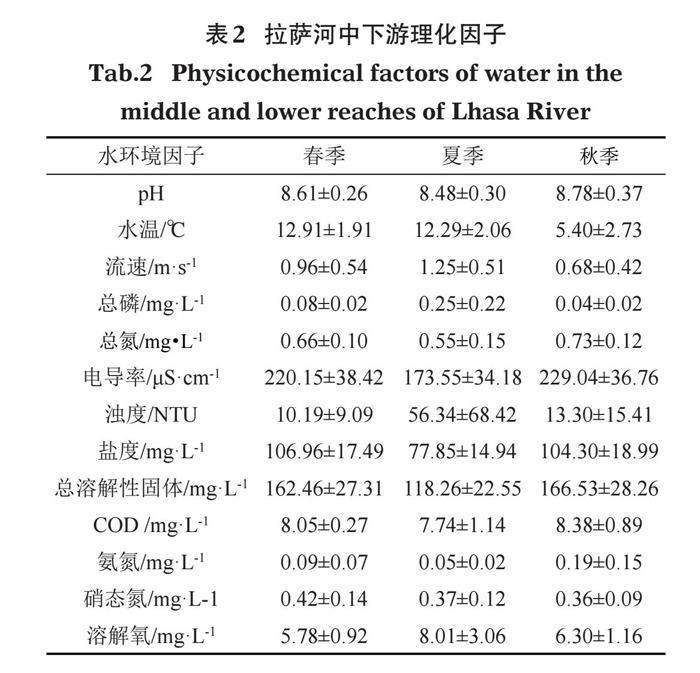
拉萨河中下游春季、夏季、秋季的水环境因子如表2所示。
由表2可得,pH、总氮、电导率、总溶解性固体和氨氮的值秋季>春季>夏季;水流速度、总磷的值夏季>春季>秋季;浊度、溶解氧的值夏季>秋季>春季;盐度值春季>秋季>夏季;硝态氮值春季>夏季>秋季。
2.2 原生动物群落物种组成
拉萨河中下游3个季节共鉴定出原生动物197种,其中,春、夏、秋分别鉴定出97、106、107种。各季节稀有种、偶见种、常见种数量及占比见表3。
2.3 原生动物群落多样性
春、夏、秋3个季节多样性指数结果见表4。其中,Richness指数(R)和Margalef指数(dM)春季[≥]夏季>秋季,Shannon-Wiener指数(H')和Simpson指数(D)春秋季节>夏季。
拉萨河中下游各样点不同季节多样性指数如图2所示,3个季节各样点多样性指数比较发现,整体上,中游段各样点(L1、L5、L6)指数高于下游段各样点(L14、L9、L12、L13)。
2.4 出现频率对物种多样性的相对贡献
原生动物稀有种、偶见种和常见种对物种多样性的相对贡献如图3所示。从各样点看,春季去除偶见种引起的Richness指数、Shannon多样性指数和Simpson指数减小效应最强,其次为去除稀有种和常见种。夏季去除偶见种引起的Richness指数、Shannon多样性指数和Simpson指数减小效应最强,其次为去除稀有种,去除常见种减小效应最弱。秋季去除偶见种引起的Richness指数、Shannon多样性指数和Simpson指数减小效应最强,其次为去除稀有种,而去除常见种引起减小效应并不明显。
比较拉萨河中下游不同季节原生动物去除稀有种、偶见种和常见种对物种多样性的减小效应,当去除偶见种时,各样点物种多样性减小效应表现最强,表明在原生动物物种类群组成中偶见种是物种多样性格局的重要贡献者。
2.5 原生动物群落相似性分析
基于Bray-Curtis距离的主坐标分析(Principal Coordinate Analysis,PCoA)和相似性分析(Analysis Of Similarities,ANOSIM)结果如图4a和图4b,拉萨河中下游原生动物群落在不同季节(R=0.18,P=0.001)和不同流域(R=0.42,P=0.001)均具有显著性差异(P<0.05),且流域间差异(R=0.42)比季节间差异(R=0.18)更大。为探究拉萨河中下游原生动物群落时空格局的形成机制,对原生动物群落相似性与环境因子及地理距离的相关性进行分析,结果如图4c和图4d,拉萨河中下游原生动物群落相似性随环境因子和地理距离的差异增大而具有显著衰减趋势(P<0.01),这表明拉萨河中下游原生动物群落差异性是由环境因子差异和地理距离差异共同作用的结果。
2.6 群落结构与环境因子的关系
利用Mantel检验分析拉萨河中下游原生动物群落结构差异与环境因子差异之间的相关性,如图5。在所测定的13个环境因子中,DO、EC、Salt、TDS、WT和TP是驱动拉萨河中下游原生动物群落组成变化的最显著环境因子(P<0.01)。
3 讨论
3.1 原生动物群落结构特征
生物多样性反映生物群落组成的复杂性和稳定性,是衡量水生态系统环境状态的重要指标(Pool et al,2014;陈泽豪等,2021)。从空间尺度上,拉萨河中下游原生动物物种多样性分析结果显示,中游段多样性指数高于下游段,表明中游原生动物群落组成较下游复杂,群落稳定性较下游好,这与方艳红等(2014)对大渡河河口春秋季原生动物的群落结构研究结果基本一致。这可能是由于中游地区主要为农牧区,生态环境较为原始,水环境受人为干扰影响较少,水质较好;而下游地区主要为拉萨市城镇区,人口密度大,受人类活动干扰较为严重,且河流易受工农业废水与生活污水等污染,对水生态系统造成一定影响,导致水质较差,多样性指数低。研究结果表明,原生动物物种多样性在人类活动干扰下,表现出较强的空间异质性。如魏洪祥等(2018)对卧龙湖浮游生物群落影响因子的研究发现,卧龙湖湿地生态系统的稳定性受不同程度人类开发活动和自然环境因素的影响,采样面积逐年下降,生物种群急剧减少,致使不同区段采集鉴定的物种种类组成上呈现明显的空间异质性,物种多样性指数上体现空间分布差异。
从时间尺度上,拉萨河中下游原生动物群落Richness指数、Margalef指数春季和夏季高于秋季,可能是由于春季(12.9℃)和夏季(12.2℃)的水温较秋季(5.4℃)更适合浮游植物生长与繁殖,浮游植物在适宜的温度范围内,生长速度加快,可为原生动物生命活动提供更充足的营养物质、食物来源及栖息场所,从而使原生动物物种数目变多、种类变丰富、多样性指数升高(金琼贝等,1991;方艳红等,2012)。这与代培等(2019)在探究太湖五里湖湖滨带浮游动物群落结构特征中原生动物生物多样性随水温的季节变化规律基本一致。原生动物作为水生态系统食物链中的初级消费者,同时受初级生产者和次级消费者种内种间竞争和捕食的影响导致其多样性指数降低(Zhu et al,2012;龚森森等,2021)。原生动物群落Shannon-Wiener指数(H')季节变化趋势均为春季和秋季高于夏季,从水生态系统食物链角度来看,可能是由于夏季水温逐渐升高,促进以原生动物为食的其他大型浮游生物和鱼类等生物的生长代谢,从而加快水体中营养物质的消耗速度,加剧物种内部竞争与捕食关系(陈旭淼和徐奎栋,2014)。这与安睿(2016)对三江平原典型湿地浮游生物群落特征研究结果类似。本研究结果表明,原生动物在应对人类不同程度干扰与水环境变化等情况时,反应灵敏迅速,其群落组成与物种多样性能够在一定程度上反映水环境适应性变化与水环境质量状况。
3.2 物种组成对物种多样性的相对贡献
物种多样性格局的形成不仅依赖于群落的构建过程,还依赖于群落中物种类群组成(Marquet et al,2004;王世雄等,2016)。群落中不同物种类群组成是决定物种多样性格局的重要因素,研究不同物种类群组成对物种多样性相对贡献是探究物种多样性格局形成过程的一个重要途径(Lennon et al,2004)。本研究首次通过去除原生动物稀有种(或偶见种、常见种)形成一个“新”的群落,然后比较“新”群落物种多样性与原群落物种多样性间减小效应来间接度量稀有种、偶见种和常见种对群落物种多样性格局的相对贡献。本研究中,当去除偶见种时,原生动物物种多样性减小效应最强,表明偶见种是原生动物物种多样性格局的重要贡献者。这与王世雄等(2018)运用此分析方法对黄土高原辽东栎群落物种多样性贡献的多尺度分析结果存在一定的差异,这可能是由于2个研究研究定义稀有种、偶见种和常见种的标准不同,导致群落物种类群组成的比例差异,因此造成不同物种类群组成对物种多样性格局的贡献不一致。也有可能是因为不同季节物种类群组成在应对环境差异变化时,具有不同的功能性状,如扩散能力或者环境耐受力,它们对物种多样性的主导作用受其自身比例及其他物种比例等因素的影响(彭少麟等,2003)。如稀有种本身对各种因素的干扰表现更为敏感,生存环境较为局限,因此在种间种内竞争过程中比例逐渐减少,去除稀有种引起的整个群落物种多样性的变化较小。
3.3 环境因子对群落结构的影响
原生动物群落结构特征不仅与食物供给、竞争及捕食—被捕食的关系等生物因子影响有关,还与pH、水温、溶解氧等非生物因子密切相关(龚森森等,2021)。使用Mantel检验方法来确定原生动物群落结构差异与环境变量差异之间的潜在相关性,结果表明,拉萨河中下游流域原生动物群落结构均与水温、总溶解性固体、盐度、电导率、总磷、溶解氧呈显著相关性。水温是影响拉萨河中下游流域原生动物群落生长和数量变化的一个重要因素,水温会影响水体的理化性质(如DO、Salt、pH),直接或间接地影响原生动物的食物来源、生长代谢和其他生命活动(Wang et al,2019)。在温度适宜的条件下,丰富的浮游植物为原生动物提供大量饵料与栖息场所,使原生动物多样性大大提高。此外,溶解氧和总磷也是影响该流域原生动物群落结构的重要因素,这与溶解氧和总磷均是生命体进行正常生命活动的必要物质密不可分(魏洪祥等,2018)。浮游生物对溶解氧依赖程度随温度升高而增加,当水温升高时,水中的溶解氧会直接限制生物的丰度,水中含氧量随温度而有昼夜和季节的变化,所以,不同原生动物对氧的需求也有一定的适应范围(沈韫芬,1990)。水体盐度可改变原生动物的细胞渗透压和水体各种离子,进而影响原生动物的生命活动。Davidson等(2010)对南海原生生物群落的组成和分布展开调查,发现原生动物群落与水温、盐度及电导率具有显著相关性,与本研究调查结果一致。总体而言,非生物因子是决定某种原生动物能否生活在某一特定生境中的决定性条件,而生物因子则是确定某一种群相对丰度的首要条件。本研究与前人对原生动物的群落结构特征研究结果类似,表明造成其群落物种丰度、组成、结构等方面的差异是原生动物不同类群生活史特征与生境条件协同作用的结果(赵瑞等,2019)。
参考文献
安瑞志,张鹏,达珍,等,2021. 西藏麦地卡湿地不同水文期原生动物优势种生态位及其种间联结性[J].林业科学,57(2):126-138.
安瑞志,潘成梅,刘洋,等,2020. 巴松措(湖)夏季浮游植物优势种群分布格局与共存机制研究[J].高原科学研究,4(2):27-36.
安宝晟,姚檀栋,郭燕红,等,2021. 拉萨河流域典型区域保护、修复、治理技术示范体系[J].科学通报,66(22):2775-2784.
安睿,2016. 三江平原典型湿地浮游生物群落特征及其与环境因子的关系[D].哈尔滨:东北林业大学.
巴桑,杨欣兰,黄香,等,2017. 拉萨河中上游春季原生动物群落特征[J].西北师范大学学报(自然科学版),53(5):70-76,83.
陈旭淼,徐奎栋,2014. 深海原生动物多样性研究进展[J].海洋科学,38(10):119-126.
陈歆,靳甜甜,苏辉东,等,2019. 拉萨河河流健康评价指标体系构建及应用[J].生态学报,39(3):799-809.
陈丽,王东波,君珊,等,2019. 拉萨河流域大型底栖动物群落结构及其与环境因子的关系[J].生态学报,39(3):757-769.
陈泽豪,杨文,王颖,等,2021. 白洋淀大型底栖动物群落结构及其与环境因子关系[J].生态学杂志,40(7):2175-2185.
代培,刘凯,周彦锋,等,2019. 太湖五里湖湖滨带浮游动物群落结构特征[J].水生态学杂志,40(1):55-63.
方艳红,黄道明,王文君,等,2012. 大渡河河口秋季浮游动物的群落结构特征[J].水生态学杂志,33(3):34-40.
方艳红,王崇,王文君,等,2014. 大渡河河口春秋季原生动物的群落结构[J].水生态学杂志,35(4):29-34.
国家环境保护总局,2002. 水和废水监测分析方法[M].北京:中国环境科学出版社.
龚森森,吴嘉伟,柴毅,等,2021. 长湖浮游动物群落结构特征及其季节变化[J].水产科学,40(3):329-338.
何柳,2019. 拉萨河流域水文地球化学特征及其风化指示[D].南昌:东华理工大学.
君珊,王东波,周健华,等,2019. 拉萨河流域浮游植物群落结构特征及与环境因子的关系[J].生态学报,39(3):787-798.
刘久潭,李颖智,高宗军,等,2020. 拉萨河流域中下游地区水化学及地表水-地下水转化关系研究[J].山东科技大学学报(自然科学版),39(5):10-20.
刘旻霞,李全弟,蒋晓轩,等,2020. 甘南亚高寒草甸稀有种对物种多样性和物种多度分布格局的贡献[J].生物多样性,28(2):107-116.
彭定志,徐宗学,巩同梁,2008. 雅鲁藏布江拉萨河流域水文模型应用研究[J].北京师范大学学报(自然科学版),(1):92-95.
彭少麟,殷祚云,任海,等,2003.多物种集合的种-多度关系模型研究进展[J].生态学报,(8):1590-1605.
索雨泽,刘洋,刘宇康,等,2021. 雅鲁藏布江中上游浮游植物污染指示种及水质评价[J].环境生态学,3(3):26-32.
沈韫芬,1990. 微型生物监测新技术[M].北京:中国建筑工业出版社.
沈韫芬,1999.原生动物学[M].北京:科学出版社.
王思齐,朱章明,2022. 中国蔷薇属植物物种丰富度分布格局及其与环境因子的关系[J].生态学报,42(1):209-219.
王世雄,赵亮,李娜,等,2016. 稀有种和常见种对植物群落物种丰富度格局的相对贡献[J].生物多样性,24(6):658-664.
王世雄,王孝安,郭华,等,2018. 稀有种和常见种对黄土高原辽东栎群落物种多样性贡献的多尺度分析[J].生态学报,38(22):8060-8069.
魏洪祥,石俊艳,王晓光,等,2018. 卧龙湖浮游生物群落的演替、影响因子及其水质评价[J].生态科学,37(1):55-61.
杨欣兰,巴桑,黄香,等,2019. 拉萨河中上游夏秋季纤毛虫群落时空变动及其与环境的关系[J].生态学报,39(9):3121-3132.
章宗涉,黄祥飞,1991.淡水浮游生物研究方法[M].北京:科学出版社.
赵瑞,赵学敏,苟婷,等,2019.广西贝江大型底栖动物群落结构时空分布特征[J].中国环境监测,35(4):40-49.
中国科学院青藏高原综合科学考察队,1981.西藏地热[M].北京:科学出版社.
Lennon J J, Koleff P, Greenwood J J D, et al,2004. Contribution of rarity and commonness to patterns of species richness[J]. Ecology Letters, 7(2) :81-87.
Marquet P A, Fernández M, Navarrete S A, et al,2004. Diversity emerging toward a deconstruction of biodiversity patterns[M]//Lomolino M,Heaneyl.Frontiers of Biogeography :New dinections in the Geography of Nature,Sunderland MA:Sinauer Associates:191-209.
Moura L C S,dos Santos S M, de Souza C A, et al,2021. Phytoplankton richness and abundance in response to seasonality and spatiality in a tropical reservoir[J].Acta Limnologica Breasiliensia,33(1):DOI:10.1590/s2179-975x11419.
Pool T K, Grenouille G, Villéger S, 2014.Species contribute differently to the taxonomic,functional,and phylogenetic alpha and beta diversity of freshwater fish communities[J]. Diversity and Distributions, 20(11) : 1235-1244.
Zhang Y J, Duo L, Pang Y Z, et al, 2018.Modern pollen assemblages and their relationships to vegetation and climate in the Lhasa Valley, Tibetan Plateau, China[J]. Quaternary International, 467:210-221.
Zhu W T, Zhang A K, Qin C X, et al, 2021.Seasonal and spatial variation of protist communities from reef water and open ocean water in patchy coral reef areas of a semi-enclosed bay[J]. Marine Environmental Research Volume,169:105407.
(责任编辑 郑金秀)
Species Distribution and Factors Driving the Protozoan Community
in the Middle and Lower Lhasa River
DA Zhen, ZHANG Peng, AN Rui‐zhi, PAN Cheng‐mei, QIAO Nan‐qian, BA Sang
( Laboratory of Wetland and Catchments Ecology in Tibetan Plateau, Faculty of Natural Sciences,
Tibet University, Lhasa 850000, P.R. China)
Abstract:Lhasa River is one of the five largest tributaries of the Yarlung Zangbo River and plays important roles as an ecological corridor, in regional climate regulation, as a water source and in maintaining biodiversity in Tibet. The current study explored species distribution patterns and factors influencing the protozoan community in the middle and lower reaches of Lhasa River, aiming to provide basic data for conserving biodiversity and water resources. In May (spring), August (summer) of 2015 and November (autumn) of 2016, qualitative and quantitative protozoan samples were collected at 17 sampling sites in the middle and lower reaches of the Lhasa River. Species composition and distribution patterns of the protozoan community and their correlation with physicochemical factors of the river water were then analyzed. Results show that: (1) A total of 197 protozoan species were identified during the three seasons, including 80 rare species, 95 occasional species, 21 common species, and 1 constant species, which accounted for 40%, 48%, 11% and 0.5% of the total species, respectively. (2) Occasional species were important contributors to the species diversity of the protozoan community in Lhasa River. (3) The Richness and Margalef indices were higher in spring and summer than in autumn, while the Shannon-Wiener and Simpson indices were higher in spring and autumn than in summer. (4) The results of principal coordinate analysis and similarity analysis based on Bray-Curtis distance showed that the protozoan community structure in the middle and lower reaches of Lhasa River varied significantly with season and river segment, with larger variations between river segments than between seasons. (5) The Mantel test was used to analyze the correlations between protozoan community parameters and environmental factors. Water temperature (WT), total dissolved solids (TDS), salinity (Salt), electrical conductivity (EC), total phosphorus (TP) and dissolved oxygen (DO) were the primary environmental factors affecting protozoan community structure.
Key words: protozoan; species diversity; environmental factors; Lhasa River
猜你喜欢
水生态学杂志(2023年4期)2023-04-29 00:44:03
西藏农业科技(2019年3期)2019-11-04 00:35:16
教育教学论坛(2019年7期)2019-03-18 11:50:12
安徽农学通报(2018年19期)2018-12-25 10:57:36
布达拉(2018年4期)2018-05-14 08:53:30
水利水电工程设计(2017年1期)2017-05-17 05:20:01
绿色科技(2016年21期)2016-12-27 10:53:26
现代农业科技(2016年20期)2016-12-20 09:17:04
科技创新导报(2016年21期)2016-12-17 12:53:50
西藏大学学报(自然科学版)(2016年1期)2016-11-15 05:23:28