
广东韩江潮州段浮游植物群落结构特征与水质评价
2023-04-29 21:57:01林小植胡苑玲王瑞旋李冬梅林鸿生查广才温茹淑吴晓琼
水生态学杂志 2023年4期
林小植 胡苑玲 王瑞旋 李冬梅 林鸿生 查广才 温茹淑 吴晓琼

摘要:为探究广东韩江潮州市饮用水源保护江段浮游植物群落结构特征及水质富营养化状况,于2019年2月至2020年1月在潮州段设立4个采样点,进行浮游植物调查和水质监测。结果表明,韩江潮州共鉴定出浮游植物8门103属290种,其中绿藻门42属99种,硅藻门21属87种,蓝藻门24属70种。全年有9个月的种类组成以硅藻门为绝对优势种,绿藻门仅在6、9、10月种类数多于硅藻门,表明该江段浮游植物组成为硅藻-绿藻型。浮游植物丰度月变化在0.83×106~1.99×106个/L,月均丰度为0.90×106 个/L,全年以硅藻门丰度最高,占47.01%。优势种11种,其中硅藻门5种,隐藻门4种,蓝藻门和裸藻门各1种,颗粒沟链藻(Melosira granulata)是绝对优势种。Shannon-Wiener多样性指数(H)为3.14~5.08,平均值4.40;Pielou均匀度指数(J)为0.69~0.99,平均值0.87;Margalef丰富度指数(D)为2.60~6.85,平均值5.08,表明水质为轻度污染。Cluster聚类和NMDS排序分析显示,物种构成可分为2个类群,类群I包括2019年2-7月,类群II包括2019年8-12月和2020年1月。Pearson相关性和冗余分析表明,影响浮游植物优势种的水环境因子主要是透明度和亚硝态氮,影响颗粒沟链藻分布的是盐度和水温,影响隐藻门分布的主要是透明度、pH和水温。参照地表水环境质量标准,韩江潮州段为Ⅱ类水,多样性指数显示为轻度污染,浮游植物群落结构和优势物种显示水体为偏中营养型。
关键词:浮游生物;群落结构;水环境;季节动态;韩江潮州段
中图分类号:Q145,X824 文献标志码:A 文章编号:1674-3075(2023)04-0052-09
由于人类经济社会活动的影响,水环境生态系统和水资源污染日益严重,许多河流和湖泊的水质富营养化正在快速恶化中(Douterelo et al, 2004;况琪军等,2005)。20世纪初,韩博平等(2003)对广东省各流域19座水库进行研究,发现大部分水库水质状况为中度污染,3座污染严重,水质已超富营养化。浮游植物是水生生态系统中最重要的初级生产者,其生长速度快,对水生态系统敏感,能够对环境干扰作出反应,被认为是水体营养状态的良好指标,也是反映水体污染状况最好的指示生物(Paerl et al, 2010;Jiang et al,2014);其群落特征及动态变化是多个环境因子的时空综合作用结果,与水环境中各种理化因子密切相关,是评价水质状况及其变化趋势的重要指标,可用于评价河流、湖泊富营养化(Kamenir et al,2004;况琪军等,2005;Suikkanen et al,2007)。已有研究表明,通过浮游生物群落结构特征结合水环境因子动态情况进行多样性、Pearson相关性和RDA分析,多角度对水质进行评价,能够更好地反映水质综合状况(吴慧双等,2020)。
韩江(115°13′~117°09′E,23°17′~26°05′N)是广东省第二大河流,也是粤东1 400万居民的饮用水源地,曾被评为“全国十大最美家乡河”之一,具有重要的饮用水、灌溉、航运等综合功能。上游由梅江和汀江汇合而成,中游为梅州的三河坝至潮州水利枢纽段,下游流经潮州分为东溪、西溪、北溪,最终注入南海。近年来,由于经济快速发展,排入江河的污染物增多,水质污染事件时有发生(张李源清和王玲, 2010),有时水质为Ⅲ级(广东省水利厅,2021)。有关韩江富营养状况和浮游植物的研究甚少,仅见零星调查报告(陈霓彤,2014)。2019年2月至2020年1月,对韩江潮州段进行浮游植物组成和周年变化调查,并检测水质指标,分析其种群结构、优势物种、污染指示物种、多样性指数等变化,旨在为水生态学评价和水环境监测提供依据,为建立韩江饮用水源健康安全环境管理机制提供参考。
1 材料与方法
1.1 采样地点
本次调查共设置4个采样点,分别是韩江西岸的竹竿山水厂(S1)和青龙庙(S4);东岸的桥东水厂(S2)和韩江大桥(S3),该江段为潮州市一级饮用水源保护江段(图1)。每月采样1次,共12次。
1.2 采样方法
浮游生物定性、定量采样方法参照《湖泊富营养化调查规范》进行(金相灿和屠清瑛,1990)。因采样点的分布和距离相对较小(<5 km),均位于广东省潮州水利枢纽上游的类湖泊环境中,统计数据取该江段4个采样点的平均值。
定量采样用2 L有机玻璃采水器取水6 L,各采样点设3个重复,使用25号浮游生物网(64 μm)过滤,以0.1 mL浮游生物计数框计数(章宗涉和黄祥飞,1991)。定性采样用25号浮游生物网,参照《湖泊富营养化调查规范》,在Nikon E100光学显微镜进行拍照并分类鉴定(胡鸿钧和魏印心,2006)。
1.3 水环境指标测定
用哈希HQ40d双通道多参数分析仪在采样点测定水体溶解氧、盐度、水温、电导率和pH;用塞氏盘测定透明度,用LGY-II型智能流速仪测定流速。
取表层水用于大肠杆菌、NH3-N、NO2-N、高锰酸盐指数的测定,采用哈纳HI83203微电脑多参数离子浓度测定仪测定水中NH3-N和NO2-N,高锰酸盐指数、大肠菌群检测参照《水和废水监测分析方法》(国家环保总局, 2002)。
1.4 数据分析
在Excel统计物种数、丰度、出现频度,DPS 9.50计算浮游植物多样性指数,SPSS 19.0分析浮游植物与水环境因子的Pearson相关关系,采用Canoco5.0对全年浮游植物优势种与水环境因子的相关关系进行冗余分析。相对丰富度(Q)、优势度(Y)指数、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)和Margalef丰富度指数(D)计算公式(吴朝和张庆国,2009;王丽卿等,2011;Siham et al, 2019)如下:
Q = (ni/N)×100% ①
Y = (ni/N)×fi ②
H = -[i=1snin] log2[nin] ③
J = H/log2S ④
D = (S - 1)/log2N ⑤
式中:ni为每月第i种浮游植物的个体数,N为全年浮游植物总个体数,fi为第i种浮游植物在12次采样中的出现的频率;n为每月浮游植物总个体数;S为每月浮游植物总种数。
浮游生物多样性指数的污染状态水质评价标准(郑丙辉等,2007)见表1。
1.5 季节变化聚类
以12个月浮游植物的种类和数量组成原始数据矩阵,用Bray-Curtis相似性系数为基础构建月份之间的相似性矩阵。采用等级聚类(UPGMA)的分类方法和非度量多维标度排序法(NMDS)分析潮州段浮游植物群落的季节变化特征。本文四季划分为春季(2-4月)、夏季(5-7月)、秋季(8-10月)、冬季(11-1月),建立聚类分析图和NMDS平面图。任意两样本间的距离代表其相似程度;聚类越近,相似程度越高。以协强系数(Stress)检验NMDS分析结果的优劣(Clarke & Warwick,2001;林小植等,2016)。
通过相似性分析(ANOSIM)检验不同矩阵间的差异显著性,组内所有位点的平均秩相似性与组间所有位点间的平均秩相似性差异用R表示,域值为[-1, 1] (林小植等, 2016) 。应用SIMPER (Similarity Percentages)方法分析每种浮游植物群落组内相似性和组间相异性的平均贡献率,将贡献率≥5%的浮游植物定义为主要特征种;对全年优势物种浮游植物进行聚类和排序分析。所有群落结构分析用PRIMER 5.0软件包(Plymouth Marine Laboratory, UK)完成。
2 结果
2.1 浮游植物种类组成
2019年2月至2020年1月的采样调查,韩江潮州段共鉴定出浮游植物8门103属290种。其中绿藻门最多,达42属99种,硅藻门21属87种,蓝藻门24属70种,裸藻门16种,甲藻门7种,黄藻门6种,隐藻门4种,金藻门1种,种类组成及其占比见图2。
从时间分布上看(图3),浮游植物物种数出现最多在2019年5月,有127种,最少的在2019年6月,仅44种。
各月的物种构成中,硅藻门、绿藻门、蓝藻门的物种在所有物种中占比较大。硅藻门的物种数在2019年2、3、4、5、7、8、11、12月和2020年1月中占比均最大,绿藻门在6、9、10月物种数大于硅藻门(图4)。因此认为韩江潮州段浮游植物的种类组成特征为硅藻-绿藻型。
2019年6月韩江潮州段仅由硅藻门、蓝藻门、绿藻门构成,其余各月均由3~7门浮游植物构成;金藻门只有分歧锥囊藻(Dinobryon drivergens)1种,仅在2020年1月采到(图3)。
2.2 优势种组成和丰度
全年Y≥0.02的优势种共有11种,其中硅藻门5种,颗粒沟链藻为绝对优势种;隐藻门4种,绿藻门、蓝藻门各1种(表2)。
2.3 浮游植物丰度构成和季节变化
韩江潮州段浮游植物月丰度在0.83×106~1.99×106 个/L,月均丰度0.90×106个/L。全年硅藻门丰度为5.06×106个/L,占全部类群的47.01%,全年的绿藻门、隐藻门、蓝藻门占比也较大,分别占20.97%、16.87%、11.38%(图5)。
浮游植物全年丰度最高值出现在2019年10月,为1.99×106个/L,月均丰度0.83×106 个/L;丰度最低值在2019年2月,为0.20×106 个/L(图6)。硅藻门全年都有出现,且数量占比大,隐藻门则在秋末冬初出现,金藻门和黄藻门在定量采样中未采集到。根据湖泊富营养类型评价的细胞密度标准(况琪军等,2005),潮州市江段水质处于贫营养与贫中营养型。
2.4 浮游植物多样性指数
韩江潮州段浮游植物Shannon-Wiener多样性指数(H)均值为4.40(3.14~5.08),2019年3月最低,为3.14;Pielou均匀度指数(J)均值为0.87(0.69~0.99);Margalef丰富度指数(D)均值为5.08(2.60~6.85)(图7)。
2.5 理化指标
根据地表水环境质量标准(GB3838-2002),韩江潮州段的高锰酸盐指数、溶解氧、氨氮、粪大肠杆菌各月平均值均在Ⅱ类水范围内。水体环境理化指标见表3。
2.6 浮游植物与水环境因子的相关性及冗余分析
浮游植物丰度与水环境因子的Pearson相关性分析显示,隐藻门丰度与流速(FR)呈显著正相关(r=0.768,P=0.011),与透明度(SD)呈极显著正相关(r=0.872,P=0.002)。
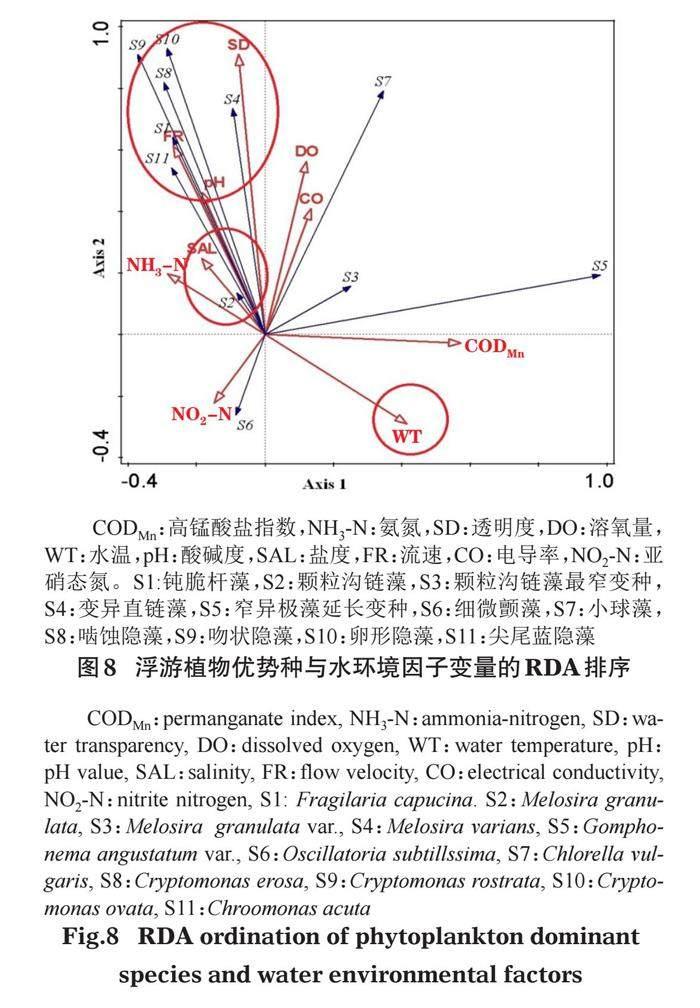
以浮游植物优势种(Y≥0.02)的生物量与各月份的相关水环境因子数据进行冗余(RDA)分析(图8)。结果表明,RDA第1、2轴的特征值分别为0.4632、0.2777,共解释了韩江潮州段浮游植物总变异数的98.4%,第1轴和第2轴浮游植物优势种与水环境因子的典型相关都达到了0.999,表明具有很强的相关性,透明度(SD)和亚硝态氮(NO2-N)对浮游植物优势种影响最大,且呈显著相关(P<0.05),大部分优势种与透明度呈正相关关系,与亚硝态氮呈现负相关关系。以隐藻门为主(S8、S9、S10、S11)的优势种与FR、pH、SD呈正相关关系,与水温(WT)呈负相关关系。硅藻门中颗粒沟链藻(S2)与盐度(SAL)呈正相关关系,与WT呈负相关关系。
2.7 聚类分析及群落特征
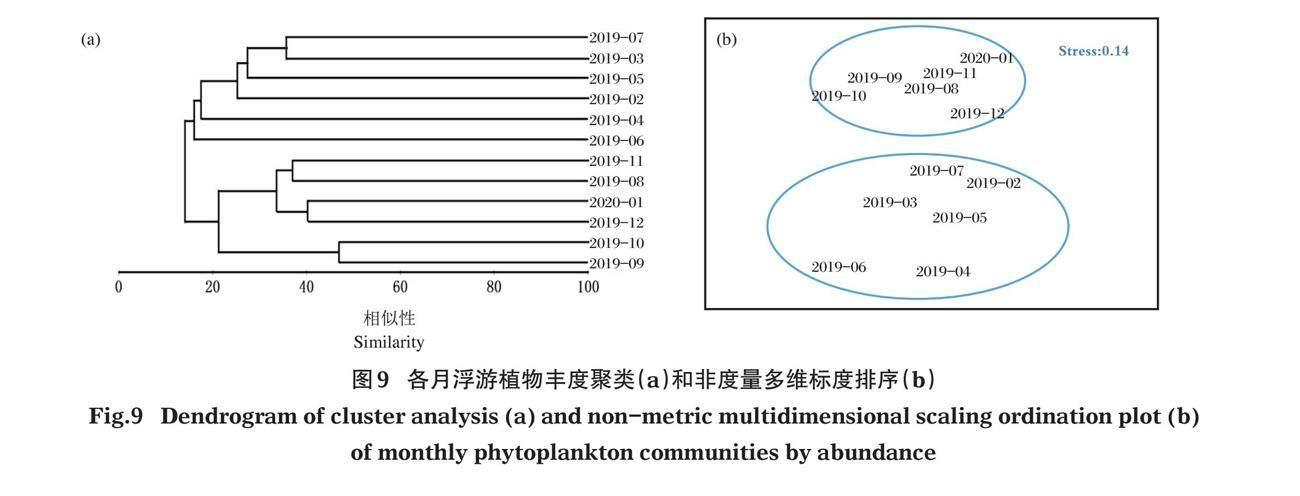
等级聚类分析表明,可将12个调查月分成2个类群,类群I包括2019年2-7月;类群II包括2019年8-12月和2020年1月(图9-a)。类群I中对群落内贡献率≥5%的种类有克洛脆杆藻(Fragilaria crotonensis)、钝脆杆藻、尖针杆藻(Synedra acus)、游丝藻(Planctonema lauterbornii)、颗粒沟链藻、变异直链藻;类群II中对群落内贡献率≥5%的种类有颗粒沟链藻、颗粒沟链藻最窄变种、变异直链藻、啮蚀隐藻、吻状隐藻、尖尾蓝隐藻。类群I与II的差异系数为86.95%,对类群间差异贡献率≥5%的种类有颗粒沟链藻、窄异极藻延长变种、吻状隐藻。
NMDS排序的协强系数为0.14,表明NMDS排序的二维点阵图结果可以接受。聚类分析的2个组群在NMDS图上能够清晰地表现在2个不同的区域中,标识为类群I和类群II(图9-b)。ANOSM差异显著性检验结果显示,r=0.887 (<1), P=0.002 (<0.01),表明2个类群之间差异极显著。
将2019年8-12月和2020年1月与2019年2-7月的多样性指数(H、D)进行独立样本T检验,发现2个类群的H和D之间均存在极显著差异(P=0.007和P=0.002),与聚类分析结果一致。
对11种全年优势物种进行Cluster聚类(图10-a)和NMDS排序分析(图10-b),NMDS排序的协强系数为0.08,表明二维点阵图可以很好地解释浮游植物物种间相似关系。11种浮游植物可分为4个类群,ANOSM显著性检验结果显示,r=0.907 (<1),P=0.001 (<0.01),表明2个群聚存在极显著差异。类群A包括卵形隐藻、吻状隐藻、啮蚀隐藻、尖尾蓝隐藻,其群落特点是在秋冬季出现。类群B包括细微颤藻,在4-5月出现。类群C包括钝脆杆藻、变异直链藻、颗粒沟链藻、颗粒沟链藻最窄变种、小球藻,其群落特点是四季常见种。类群D包括窄异极藻延长变种,在10月中出现。
3 讨论
3.1 浮游植物的群落结构特征与季节演替
浮游植物是重要的生产者,其群落构成和特征是水生态系统的综合表现(Jiang et al,2014)。韩江潮州段12个月的浮游植物构成可分为2个类群,客观反映了该江段浮游植物组成的季节性波动特征。韩江地处广东东部,表层水温19.8~30.1℃,适合大多数耐高温藻类生长,特别是硅藻门、绿藻门和蓝藻门。在类群I和类群II中,对群落贡献率≥5%的共有物种是颗粒沟链藻和变异直链藻,可见在浮游植物群落结构中,同一水体不同时间的类群也存在群落组成的边界模糊性;况琪军等(2005)研究中也有类似阐述,即不同类型水体的指示物种均有营养过渡性藻类存在,有的藻类适应性很强,能够生存在不同营养水平的水体中(金相灿和屠清瑛,1990;况琪军等, 2005) 。
浮游植物群落结构和丰度受到季节环境因子、水动力学和营养盐的共同作用(韩博平等,2003;Jiang et al,2014 )。从潮州段浮游植物丰度看,丰度最小的是2月,正处于冬春季节的交替期;与陈霓彤(2014 )对韩江下游梅溪河段的研究结果一致。2月雨量少,水温最低,水的流动性较小,其浮游植物构成主要是能够适应低温生活的硅藻、绿藻和裸藻,其中硅藻数量最多,占59.61%,裸藻和绿藻各占10.10%和30.29%。这种浮游植物丰度构成大致与同处亚热带地区的广东省新丰江水库相类似,其浮游植物丰度最低也出现在2月(赵孟绪等,2005);而珠江广州段的周年调研发现,其丰度最低出现在6月,由于该江段为重度富营养化水体,其浮游植物丰度构成中蓝藻、绿藻的占比较大(张俊逸等, 2011) 。本研究中,浮游植物丰度变化在3-5月出现一个小高峰,9-12月出现最高峰,其原因主要与韩江潮州段的水文环境因子和季节有关。冬春交替季节(2月)的水温逐渐上升,浮游植物初级生产力会加速消耗冬季积累下来的营养盐,加之降水量少、农业灌溉、生活用水和自然蒸腾作用,使得浮游植物丰度经过冬季的峰值之后逐渐降低到较低水平;6月是夏季汛期降雨量最大的月份,水流量大,流速较快,大量颗粒物沉降到底部,营养水平相对较低,水温较高,不利于浮游植物的漂浮生活,因此浮游植物的丰度和初级生产力在此阶段也较低;秋冬季水流相对平稳,对流垂直混合使得表层营养盐得到补充,阳光充足,水温适宜,因而浮游植物的生长得到了较好的环境支持,初级生产力增长最大。
3.2 浮游植物群落结构与环境因子的关系
透明度是评价水质的重要指标,是表征水体中悬浮物多少的重要依据,也可以反映水的浑浊程度。当透明度降低时,会影响光在水下的穿透和传播,导致浮游植物的光合作用受阻,不利于藻类生长;在淮河流域西淝河、里下河大纵湖也发现浮游植物与透明度存在显著相关关系(邓建明等,2010;王顺天等,2020;张珊等, 2020) 。
亚硝态氮是氨氮氧化或硝态氮还原过程的中间产物,环境污染加剧导致亚硝态氮含量上升(王九龙等,2012) ;氮浓度变化会直接影响浮游植物生长,不适合耐低氮的硅藻生长,但有助于耐高氮的藻类成为优势种(Elser et al, 2009) ,与卢云黎等(2021) 报道的汉江集家嘴受到亚硝态氮的影响一致。硅藻门全年数量分布广,在秋末冬初时丰度很高,21℃为其适宜生存温度,硅藻门的颗粒沟链藻是绝对优势种,直链藻对水环境变化非常敏感,容易受到水温影响,而且其有一定的适盐性,相比其他藻类,更能适应高盐度,在富营养化水体及污染水体中极易形成优势种(Wang et al, 2009;陈威等, 2018) 。
隐藻门的繁殖主要集中在秋冬季,由于其特殊的光敏感度和营养型,在全年温度较低的时期为优势藻种(陈霓彤,2014) ;而高pH值的碱性环境有助于藻类进行光合作用并促进其增殖(徐畅,2020) 。本研究中,隐藻门的丰度与透明度呈正相关,与水温呈显著负相关,符合隐藻门的生长特点。
2019年2月至2020年1月,潮州段浮游植物种类组成的变化与韩江地区的气候有关,受海洋性东南季风的影响,4-9月为每年的汛期,阴天多,降水量占全年降雨量的70%以上。浮游植物的群落结构与光照有关,阴天会影响光合作用(林少君等, 2005) ,物种构成较单一,造成2019年6月的物种数较少。
3.3 不同指标的韩江潮州段水质评价
理化指标单因子评价法是我国水质监测公报的综合水质评价方法,参照地表水环境质量标准(GB3838-2002),选择单项最差指标所属类别来确定水域综合水质类别(戴润泉等, 2004;尹海龙和徐祖信,2008;丛铭等, 2021)。结合表4,本次监测表明韩江潮州段水质属于II类,水体为碱性。
多样性指数通常是研究浮游植物群落结构特征和动态规律的测度工具,同时也是判断水体富营养状况的常用指标。H、J、D指数值越大,其群落结构越复杂,群落所包含的物种多度和丰度信息量也越大,稳定性越强,水质越好;但多样性指数减小时,群落结构的物种多度和丰度会趋于简单,稳定性变弱,水质变差(金相灿和屠清瑛,1990;况琪军等, 2005) 。根据多样性指数评价标准(表1),韩江潮州段水质总体处于轻度污染状态,对比广东省部分河湖的多样性指数(表5),潮州段水质污染程度比小东江下游、连江、孔江、陆丰海域、珠海海域、珠江广州段低,总体水质较好。
湖泊的富营养化与浮游植物群落密切相关,其结构特征及优势种变化趋势是分析和预警水体富营养化和蓝藻水华的依据之一(金相灿和屠清瑛,1990)。研究表明,浮游藻类的生物量及种类组成,对于评价河流湖泊富营养化状况是有效的(江源等,2013;Jiang et al, 2014)。韩江潮州段浮游植物种类组成为硅藻-绿藻型(物种33.90%、29.79%,丰度20.24%、46.69%),根据不同藻类组成类型的指示作用显示,韩江潮州江段为寡-中营养型至中营养型,从蓝藻的种类和数量判断(物种23.97%、丰度11.52%),该江段为偏中营养型。
藻类作为指示物种,其种类和数量可以反映水体营养状况,特别是对于优势物种的存在,其指示等级可划分可为超富营养型、富营养型、中营养型和寡营养型(况琪军等,2005;张丽娜和于洪贤,2010);通过对浮游植物的优势种进行等级划分(表2),大多数优势种的指示等级为β-ms型和α~β-ms型,表明水体营养型为中营养型至中-中富营养型。
参考文献
陈霓彤,2014. 韩江下游梅溪河段浮游藻类动态监测及分析[J]. 城镇供水, (1):32-34.
陈威,魏南,金小伟,等,2018. 松花江哈尔滨段藻类植物分布及其与环境因子的关系[J]. 中国环境监测, 34(4):102-110.
丛铭,阳辉,张晓静,等,2021. 单因子法与可变模糊法在水质评价中的应用[J]. 南水北调与水利科技, 19(4):720-728.
戴润泉,臧小平,邱光胜,2004. 三峡水库蓄水前库区水质状况研究[J]. 长江流域资源与环境,(2):124-127.
邓建明,蔡永久,陈宇炜,等,2010. 洪湖浮游植物群落结构及其与环境因子的关系[J]. 湖泊科学, 22(1):70-78.
广东省水利厅,2021. 2008-2020水资源公报[N/OL]. http://slt.gd.gov.cn/tjsj8778/index.html.
国家环保总局,2002. 水和废水监测分析方法[M]. 4版. 北京:中国环境科学出版社.
韩博平,李铁,林旭钿,2003. 广东省大中型水库富营养化现状与防治对策研究[M]. 北京:科学出版社.
何森华,丘泓欣,邓婉璐,等,2021. 广东孔江国家湿地公园水体中的浮游植物群落结构特征[J]. 湿地科学, 19(2):218-225.
胡鸿钧,魏印心,2006. 中国淡水藻类——系统、分类及生态[M]. 北京:科学出版社.
黄报远,李秋华,陈桐生,2009. 广东连江梯级电站开发后春季河流浮游生物群落结构特征[J]. 安徽农业科学, 37(7):3137-3140.
江源,彭秋志,廖剑宇,等,2013. 浮游藻类与河流生境关系研究进展与展望[J]. 资源科学, 35(3):461-472.
金相灿,屠清瑛,1990. 湖泊富营养化调查规范[M]. 北京:中国环境科学出版社.
况琪军,马沛明,胡征宇,等,2005. 湖泊富营养化的藻类生物学评价与治理研究进展[J]. 安全与环境学报, 5(2):87-91.
林少君,顾继光,魏鹏,等,2005. 广东省公平水库与星湖生态特征的对比分析[J]. 生态学杂志,(7):773-779.
林小植,李冬梅,刘焕章,等,2016. 广东韩江潮州江段鱼类多样性及季节变化[J]. 生物多样性, 24(2):185-194.
卢云黎,王琪,罗春艳,2021. 汉江集家嘴2017-2019年浮游植物变化研究[J]. 水利水电快报, 42(5):43-48.
粟丽,黄梓荣,陈作志,2017. 广东沿岸不同海域浮游植物群落结构特征的比较分析[J]. 海洋环境科学, 36(1):61-65.
王九龙,张明明,王金叶,2012. 青岛农业大学生活饮用水亚硝态氮检测与情况分析[J]. 青岛农业大学学报(自然科学版), 29(2):111-114.
王丽卿,施荣,季高华,等,2011. 淀山湖浮游植物群落特征及其演替规律[J]. 生物多样性, 19(1):48-56.
王顺天,雷俊山,贾海燕,等,2020. 三峡水库浮游植物群落特征及水体富营养化评价[J]. 三峡生态环境监测, 5(1):32-41.
吴朝,张庆国,2009. 淡水浮游生物多样性及数量分析方法研究进展[J]. 安徽农学通报, 15(12):41-42.
吴慧双,胡泓,李正炎,等,2020. 2018年秋季大沽河浮游生物群落特征及其与水环境因子的关系[J]. 海洋湖沼通报,(5):148-154.
徐畅,2020. 微山湖浮游生物群落时空变化特征及其驱动因子分析[D]. 济南:济南大学.
尹海龙,徐祖信,2008. 河流综合水质评价方法比较研究[J]. 长江流域资源与环境, (5):729-733.
张才学,詹冬玲,陈春亮,等, 2010. 小东江茂名段浮游生物及污染状况分析[J]. 广东海洋大学学报, 30(4):22-28.
张俊逸,蒋江峦,刘擎,等,2011. 珠江广州段微生物和浮游植物群落与水质特征研究[J]. 水生态学杂志, 32(2):38-46.
张李源清,王玲,2010. 紫金矿业污水入粤致渔业受损[N]. 中国经济时报, 2010-07-23(1).
张丽娜,于洪贤,2010. 牡丹江海浪河夏季浮游植物群落结构和水质评价[J]. 水产学杂志, 23(1):40-46.
张珊,商乃萱,张金羽,等,2020. 淮河流域西淝河浮游植物群落结构特征[J]. 水生态学杂志, 41(4):81-89.
章宗涉,黄祥飞,1991. 淡水浮游生物研究方法[M]. 北京:科学出版社.
赵孟绪,雷腊梅,韩博平,2005. 亚热带水库浮游植物群落季节变化及其影响因素分析——以汤溪水库为例[J]. 热带亚热带植物学报, (5):386-392.
郑丙辉,田自强,张雷,等, 2007. 太湖西岸湖滨带水生生物分布特征及水质营养状况[J]. 生态学报,(10):4214-4223.
Clarke K R, Warwick R M, 2001. Change in Marine Communities: an An Approach to Statistical Analysis and Interpretation[M]. PRIMER-E Ltd., Plymouth.
Douterelo I, Perona E, Mateo P, 2004. Use of cyanobacteria to assess water quality in running waters[J]. Enviromental Poluution, 127:377-384.
Elser J J, Andersen T, Baron J S, et al, 2009. Shifts in Lake N:P Stoichiometry and Nutrient Limitation Driven by Atmospheric Nitrogen Deposition[J]. Science, 326:835-837.
Jiang Y J, He W, Liu W X, et al, 2014. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu) [J]. Ecological Indicatiors, 40:58-67.
Kamenir Y, Dubinsky Z, Zohary T, 2004. Phytoplankton size structure stability in a meso-eutrophic subtropical lake[J]. Hydrobiologia, 520(1):89-104.
Paerl H W, Rossignol K L, Hall S N, et al, 2010. Phytoplankton Community Indicators of Short-and Long-term Ecological Change in the Anthropogenically and Climatically Impacted Neuse River Estuary, North Carolina, USA[J]. Estuaries and Coasts, 33(2):485-497.
Siham A, Somia H, Abdessamad R M, et al, 2019. Seasonal variation of water quality and phytoplankton dynamics and diversity in the surface water of Boukourdane Lake, Algeria[J]. Arabian Journal of Geosciences, DOI: 10.1007/s12517-018-4164-4
Suikkanen S, Laamanen M, Huttunen M, 2007. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea Estuarine[J]. Coastal and Shelf Science, 71(3/4): 580-592.
Wang C, Li X H, Lai Z N, et al, 2009. Seasonal variations of Aulacoseira granulata population abundance in the Pearl River Estuary[J]. Estuarine, Coastal and Shelf Science, 85(4):585-592.
(责任编辑 万月华)
Phytoplankton Community Structure and Water Quality Assessment
of the Chaozhou Section of Hanjiang River
LIN Xiao‐zhi1, HU Yuan‐ling1, WANG Rui‐xuan1, LI Dong‐mei1, LIN Hong‐sheng1,
ZHA Guang‐cai1, WEN Ru‐shu2, WU Xiao‐qiong1
(1. Hanshan Normal University, Chaozhou 521041, P. R. China;
2. Jiaying University, Meizhou 514015, P. R. China)
Abstract:Hanjiang River is the second largest river in Guangdong Province and serves as the drinking water source for eastern Guangdong. In this study, we characterized the phytoplankton community structure and eutrophication status of the Chaozhou section of Hanjiang River, which is the drinking water source for Chaozhou City. From February 2019 to January 2020, phytoplankton and water quality monitoring was carried out monthly at 4 sampling sites, focusing on phytoplankton community structure, dominant species, diversity indices, pollution indicator species and 11 water environment parameters. Our aim was to provide data for aquatic ecosystem evaluation and water quality monitoring of Hanjiang River, and to provide a reference for developing a management strategy for maintaining a healthy and safe environment of drinking water source. A total of 290 phytoplankton species from 103 genera and 8 phyla were identified, including Chlorophyta (99 species, 42 genera), Bacillariophyta (87 species, 21 genera) and Cyanophyta (70 species, 24 genera). While species richness of the diatom phylum was absolutely dominant for nine months of the year, Chlorophyta had higher species richness in June, September and October. Thus, the phytoplankton community of the river section was of the diatom-green algae type. The monthly abundance of phytoplankton varied from 0.83×106 to 1.99×106 cells/L, with an average abundance of 0.90×106 cells/L. The abundance of diatoms was the highest, accounting for 47.01% of the total. There were 11 dominant species identified during the investigation, consisting of 5 Bacillariophyta species, 4 Cryptophyta species, 1 Cyanophyta species and 1 Euglena species, among which Melosira granulata was the absolutely dominant species. The Shannon-Wiener diversity (H), Pielou evenness (J) and Margalef richness (D) indices were in the range of 3.14-5.08, 0.69-0.99 and 2.60-6.85, with the average values of 4.40, 0.87 and 5.08, respectively, indicating a slightly polluted condition. Cluster analysis and NMDS ordination results show that phytoplankton communities from February to July 2019 was clustered into one group, and communities from August to December 2019 and January 2020 was clustered into another group. Pearson correlation and redundancy analysis show that transparency and nitrite nitrogen were the primary water environmental factors affecting the dominant species of phytoplankton. The factors affecting the distribution of Melosira granulata were salinity and water temperature, and the factors affecting the distribution of Cryptophyta were transparency, pH and water temperature. According to the environmental quality standard of surface water, the Chaozhou section of Hanjiang River met the standard of Class II surface water. The diversity indices also indicated that the water was lightly polluted. Analysis of phytoplankton community structure and dominant species indicated that the water is mesotrophic.
Key words: plankton; community structure; water environment; seasonal dynamics; Chaozhou section of Hanjiang River
猜你喜欢
卷宗(2016年10期)2017-01-21 16:32:59
热带农业科学(2016年11期)2017-01-21 14:59:21
热带农业科学(2016年11期)2017-01-21 14:57:08
环球人文地理·评论版(2016年5期)2017-01-03 03:23:34
科技视界(2016年18期)2016-11-03 23:52:41
中国科技博览(2016年22期)2016-11-01 15:27:53
中国市场(2016年21期)2016-06-06 05:47:19
科技视界(2016年11期)2016-05-23 23:15:43
河北渔业(2015年10期)2015-10-15 11:14:57
河北渔业(2015年7期)2015-07-21 11:06:52