
日本沼虾高磷脂饲料中适宜能量蛋白比的研究
2018-07-04 09:39:26蔡春芳丁惠明叶元土崔广同任胜杰邱小亮
中国饲料 2018年10期
王 志,蔡春芳*,丁惠明,叶元土,崔广同,任胜杰,邱小亮
(1.苏州大学基础医学与生物科学学院,江苏省水产动物与营养重点实验室,江苏苏州 215123;2.苏州市阳澄湖国家现代农业示范区发展有限公司,江苏苏州 215123)
在三大类能量营养物质中,蛋白质相对比较昂贵,功能强大,因此是饲料质量和价格最重的评价依据。然而蛋白质作为供能物质,其功能可被脂肪和糖类所代替,因此饲料中充足的脂肪和糖可起到节约蛋白质的作用(Cui等,2017;江星等,2013)。然而,任何水产动物对营养需求的考量都必须基于各种营养素之间的平衡与互作(Silva等,1991),饲料能量营养素的不平衡可能会导致脂肪在甲壳动物肝胰腺中蓄积(张南南等,2016;González-Fél等,2002),甚 至 会 抑制水产动物的正常生长(Zhao等,2015;Xu等,2013),限制饲料中蛋白质的消化和利用(陈乃松等,2012),造成机体的氧化损伤等(Du等,2008)。因此配制饲料时还要注意其能量蛋白比。
本课题组在前期关于日本沼虾(Macrobrachium nipponense)脂类营养需求研究中发现,磷脂可改善日本沼虾对脂类的利用性(磷脂影响日本沼虾对饲料脂肪的需要量,动物营养学报,待刊)。本研究将进一步探讨高磷脂水平下日本沼虾饲料中适宜的能量蛋白比,以期为日本沼虾饲料配方的设计和完善提供科学依据。
1 材料和方法
1.1 试验饲料 本试验以鱼粉、植物性蛋白质饲料和干酪素为蛋白源(鱼粉为定量添加,满足饲料的适口性),将鱼油、豆油、磷脂油比1∶1∶4混合为脂肪源,以糊精为糖源。设置三个蛋白(36%、40%、44%)和两个可消化能水平(11和12 MJ/kg),配制成6种试验饲料。饲料配方见表1。饲料的制作过程参照本实验室标准操作流程,简述之:通过粉碎机将固态饲料原料粉碎并过80目筛,均匀混合后加入脂肪源重新混合均匀,并添加一定量的水混匀,在双螺杆挤条机中制成直径为1.0 mm的颗粒饲料,风干后置于自封袋密封,-20℃保存备用。
1.2 养殖管理 试验在苏州市相城区阳澄湖现代农业产业园有限公司的研究生工作站进行。试验用虾苗购自江苏省苏州市甪直镇本地虾苗繁育场,幼虾初始体质量为(0.28±0.06)g,健康无病,规格整齐。试验虾在养殖水池暂养15 d,禁食24 h 后,分养于24个100 cm×100 cm×100 cm的塑料缸内。缸内盛水约300 L,并放置有生物性和非生物性的附着基供日本沼虾攀附。每缸放养200尾。每种饲料投喂4缸试验虾,随机分配投喂。每天上午7:30和下午18:30各投饲1次,采用饱食投喂,同时及时捞出死虾并称重。试验用水为纱网过滤、沉淀24 h后的湖水。每天用虹吸管排污1次,隔天换水1/3。试验期间水温24~28℃,水体pH为7.5~8.1,氨氮低于0.05 mg/L,溶氧保持在6.2~7.1 mg/L。养殖周期为8周。饲养结束后,禁食24 h,采样。
1.3 分析与测定
1.3.1 生长性能、饲料效率 将每缸所有日本沼虾捞出后分别称总重,记录尾数,用于计算增重率(WG)、成活率(SR)、特定增长率(SGR)及饲料系数(FCR)。每缸取6尾虾,剥离其完整肝胰腺称重,用于计算其肝体比值(HSI)。
1.3.2 体成分和饲料概略养分测定 称重后的虾用于水分、粗蛋白质、粗脂肪等体成分分析,其中水分含量测定采用低温冷冻干燥法(LJB 18型冷冻干燥机,北京四环科学仪器厂有限公司);粗蛋白质含量测定采用凯氏定氮法(GB 5009.5-2010。所用消化仪:LNK 87型,江苏省宜兴市科教仪器研究所;蒸馏仪:KN 520型,济南阿尔瓦仪器有限公司);粗脂肪含量测定采用索氏抽提法(GB/T 14772-2008);灰分含量的测定采用GB 5009.4-2010方法测定,样品先于200℃ 碳化至不冒烟,再于550℃ 陶瓷纤维马弗炉(8-10 TP型,上海慧泰仪器制造有限公司)中灼烧至恒重。饲料总能采用全自动氧弹量热仪(PARR 6300型,上海昌吉地质仪器有限公司)测定;总糖的测定参考文献(李如亮等,1998)中的方法,其余成分的测定同体成分测定方法。饲料概略养分见表1。
1.3.3 计算与统计分析
相关的计算公式如下:
成活率/%=Nt/N0×100;
饲料系数 =m1/(mt-m0);
特定生长率 /%/d=(lnmt- lnm0)/t×100
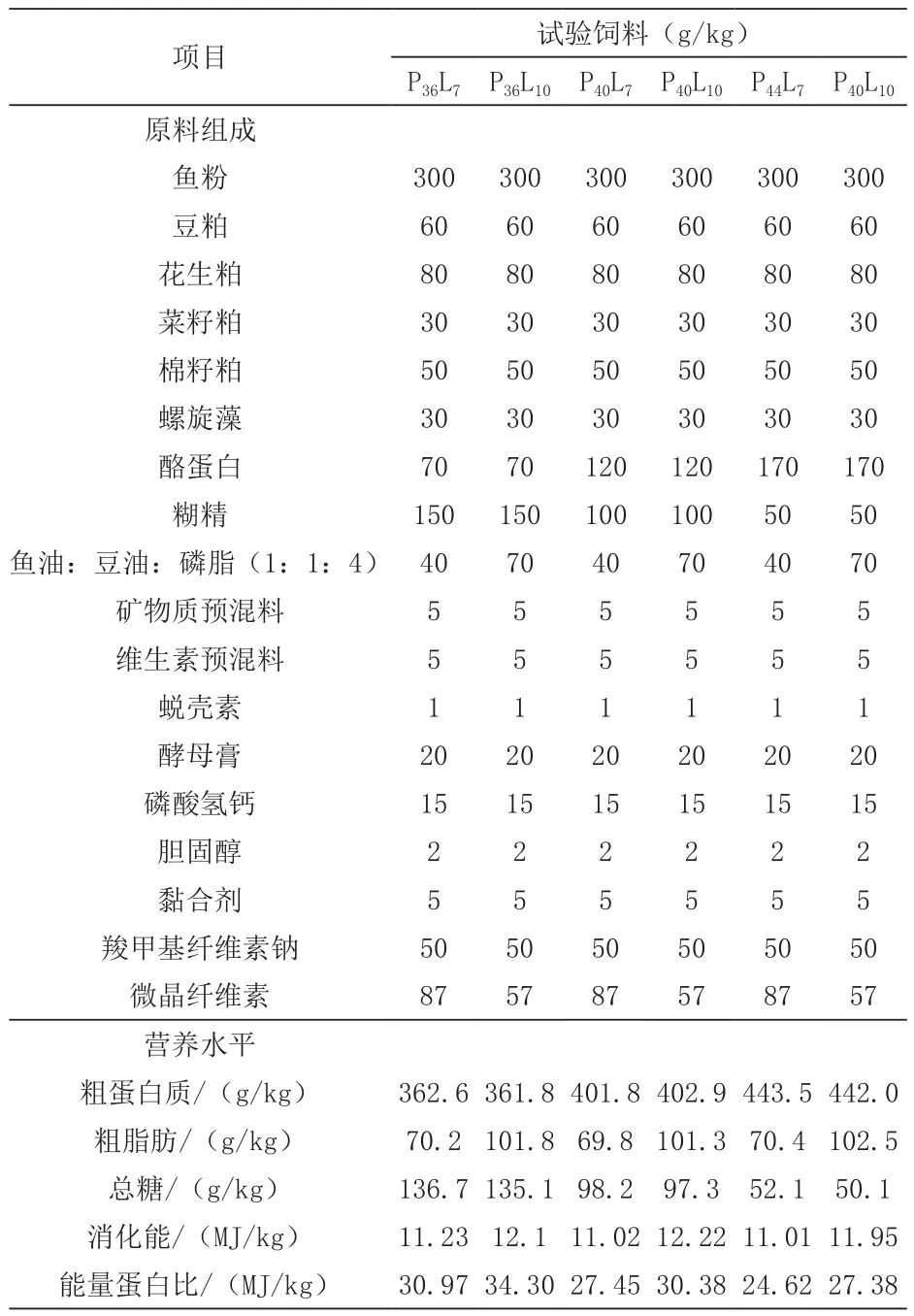
表1 试验饲料配方及营养水平(风干基础)
增重率 /%=(mt-m0)/m0×100 ;
蛋白质效率=(mt-m0)/m2;
肝体指数/%=m3/mt×100。
式中,Nt为试验结束存活虾数量(尾);N0为试验开始虾数量(尾);mt为试验结束体质量(g,含死虾质量);m0为试验开始体质量,g;m1为摄入饲料量(干物质质量),g;m2为蛋白质摄入量,g;t为试验时间,d;m3为肝胰脏质量,g。
试验数据以“平均值±标准差”表示,所有数据采用SPSS 22.0进行单因子方差分析(one-way ANOVA)和双因子方差分析(two-way ANOVA),同时进行Duncan氏多重比较,P<0.05为差异显著,P<0.01为差异极显著。此外,就不同能量水平下SGR、FCR、PER与E/P间的相关性采用回归分析。
2 结果
由表2可知,各组虾的 WG、SGR、PER和FCR差异显著(P<0.05),虾体水分、粗蛋白质、粗脂肪与粗灰分含量差异不显著(P>0.05)。当摄食的饲料的蛋白含量为36%,E/P比为34.30 MJ/kg时,日本沼虾的WG和SGR最高,FCR最低。双因子方差分析结果表明:随着饲料蛋白水平的增加,日本沼虾的SR、WG、SGR和FCR均无显著性差异(P>0.05),PER呈现显著下降趋势(P<0.05)。随着可消化能水平的提高,日本沼虾的WG、SGR、PER显著提高(P<0.01)。
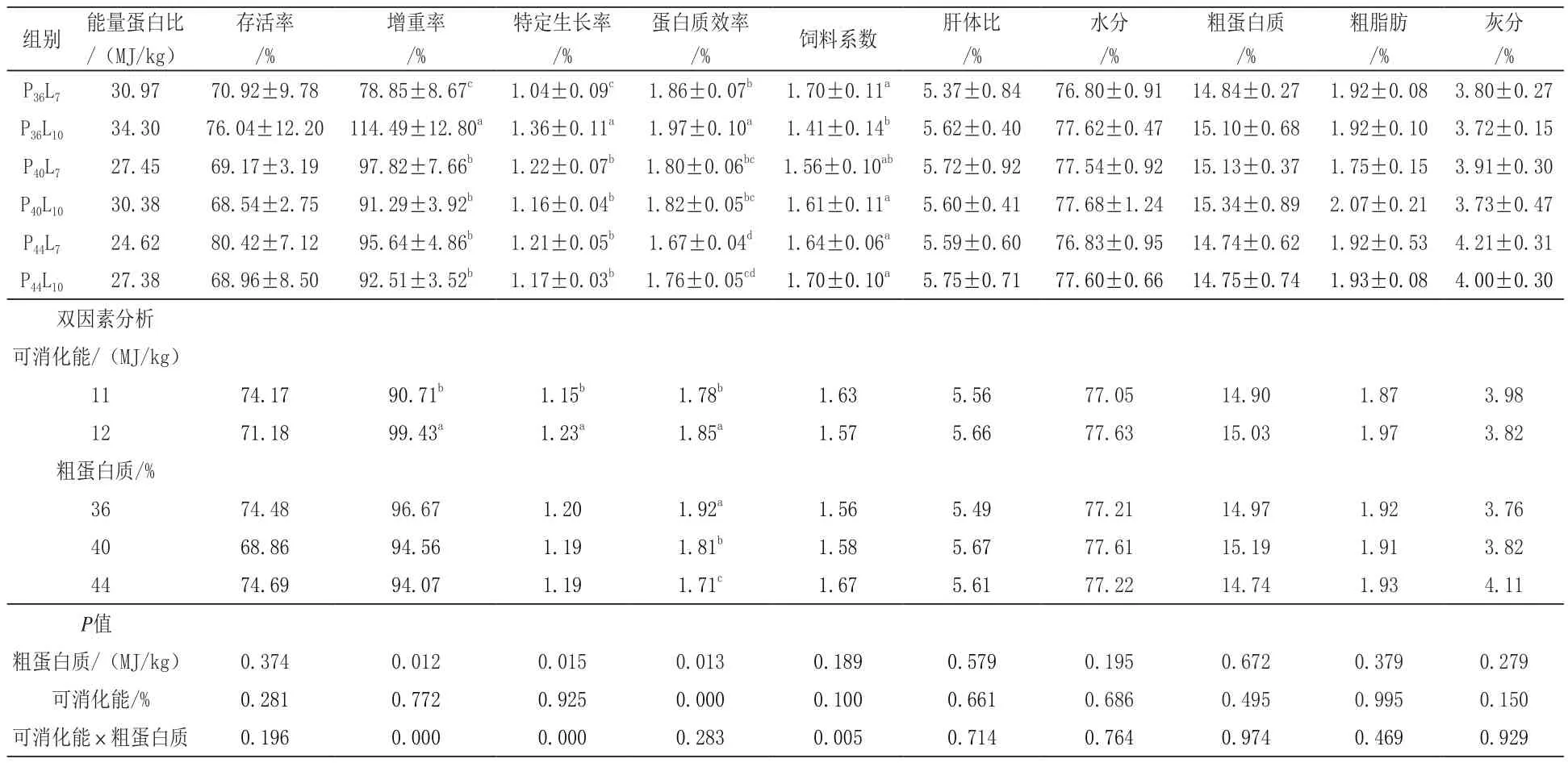
表2 不同E/P的饲料对日本沼虾生长性能和体组成的影响
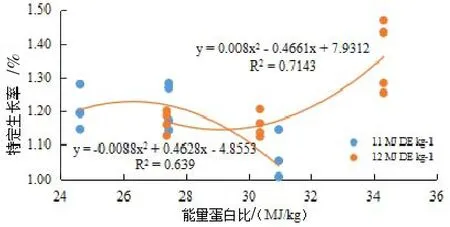
图1 能量蛋白比与特定生长率的关系
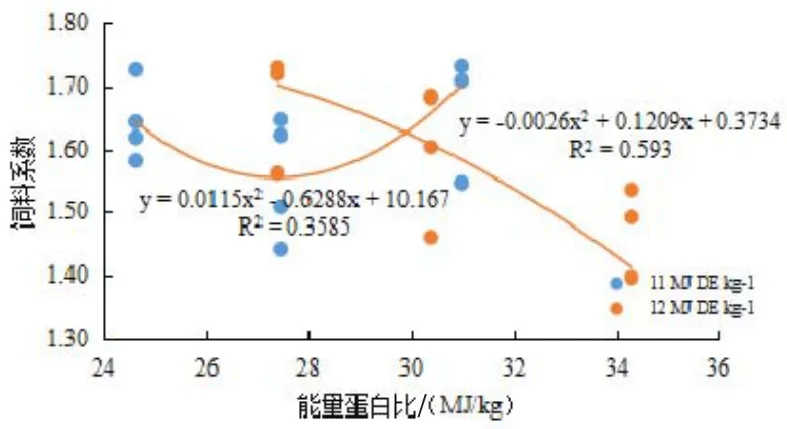
图2 能量蛋白比与饲料系数的关系
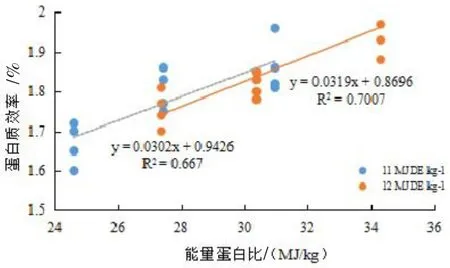
图3 能量蛋白比与蛋白质效率的关系
回归分析结果表明,可消化能水平为11 MJ/kg时,日本沼虾SGR随E/P升高(即蛋白含量下降)呈先升高后下降的态势,获得最高SGR的 E/P值为26.27(图1);可消化能水平为12 MJ/kg时,日本沼虾SGR随E/P的升高而升高,FCR随E/P的升高而下降;(见图1,图2)。在可消化能一定的情况下,PER都随着E/P的升高而升高,可消化能为11 MJ/kg时,每增加1 MJ/kg能量可提高PER 0.0302;可消化能为12 MJ/kg时,每增加1 MJ/kg能量可提高PER 0.0319(见图3)。
3 讨论
本试验结果表明随着可消化能水平升高,PER上升。摄入的蛋白质除了用于构成机体组成成分等功能外,还可用于作为能量消耗(Hu等,2008),当饲料中脂肪和糖含量丰富时,动物会利用脂肪和糖类氧化供能,降低蛋白质作为能量消耗的比例,使摄入的蛋白能够更多地用于构成机体,起到节约蛋白质的作用(Catacutan等,2002;Lee等,2002)。本研究中饲料能量的差异很大程度上是由于脂肪含量的不同(表1),因此PER的提高主要是因为脂肪对蛋白质的节约作用。研究结果也提示高磷脂时饲料中脂肪水平达10%时仍能被日本沼虾很好的利用,与我们前期研究一致。
试验结果还表明,蛋白质水平从36%升高到44%,PER下降,SGR最高值也在饲料蛋白为36%时。李贵锋等在探究不同蛋白质(26%、30%、33% 和 36%)和能量水平(13.5和 14.5 MJ/kg)对建鲤幼鱼(Cyprinus carpiovar Jian)生长性能的影响时也发现,在饲料能量较高时(14.5 MJ/kg),33%的蛋白质水平就能够获得最佳的生长性能和FCR(李贵锋等,2012)。Ai等报道用高蛋白饲料喂养花鲈(Lateolabrax japonicus)幼鱼并不能获得最高的SGR和最低的FCR(Ai等,2004)。这些研究结果一致表明过分追求高蛋白并不利于养殖动物的生长。就日本沼虾生长而言,本研究结果表明饲料中蛋白36%、可消化能为12 MJ/kg相对比较合适。
有趣的是,不同能量水平下,日本沼虾对于E/P变化的响应不同。在可消化能水平为11 MJ/kg时,日本沼虾SGR随着E/P升高(即蛋白水平下降)呈先升高后下降的态势,获得最高SGR的 E/P值为26.27 MJ/kg(图1)。这一现象与在克氏原螯虾(Procambarus clarkii)(于宁等,2014)、尖齿胡鲶(Clarias gariepinus)(Tibbetts等,2015)和黑线鳕(Melanogrammus aeglefinus)(Ali等,2015)上的研究结果相似,表明日本沼虾在可消化能较低(11 MJ/kg)时,蛋白质水平容易成为生长的制约因素。而当可消化能提高到12 MJ/kg时,日本沼虾的SGR随E/P的升高而升高,FCR随E/P的升高而下降(见图1,图2)。本实验结果提示,日本沼虾饲料中适宜的E/P值为34.30 MJ/kg或更高,高于虞冰如等(1990)在日本沼虾上研究结果(33.48 MJ/kg),这可能与饲料中较高的磷脂水平改善了脂类的利用有关(Sánchez等,2014)。
E/P与PER回归分析结果表明,可消化能为11 MJ/kg时,每增加1 MJ/kg能量可提高PER 0.0302;可消化能为12 MJ/kg时,每增加1 MJ/kg能量可提高PER 0.0319。可见能量对蛋白质的节约作用并没有随饲料能量水平的升高而下降,提示日本沼虾饲料中能量水平可进一步提高,以节约更多的蛋白质。
[1]Cui Y,Ma Q,Limbu S M,et al. Effects of dietary protein to energy ratios on growth,body composition and digestive enzyme activities in Chinese mitten-handed crab,Eriocheir sinensis[J].Aquaculture Research,2017,48(5):2243~ 2252.
[2]江星.中华绒螯蟹饲料中适宜糖源,蛋白能量比及其原料消化利用率的研究[D].华东师范大学,2013.
[3]Silva SSD,Gunasekera RM,Shim KF. Interactions of varying dietary protein and lipid levels in young red tilapia:evidence of protein sparing[J]. Aquaculture,1991,95(3~ 4):305~ 318.
[4]张南南. 基于适宜饲料蛋白和能量水平下的日本沼虾(Macrobrachium nipponense)适宜蛋白源和脂肪源研究[D].华东师范大学,2016.
[5]González-Félix M L,Iii D M G,Lawrence A L,et al. Effect of Various Dietary Lipid Levels on Quantitative Essential Fatty Acid Requirements of Juvenile Pacific White Shrimp Litopenaeus vannamei[J]. Journal of the World Aquaculture Society,2002,33(3):330~340.
[6]Zhao J,Wen X,Li S,et al. Effects of dietary lipid levels on growth,feed utilization,body composition and antioxidants of juvenile mud crab Scylla paramamosain,(Estampador)[J].Aquaculture,2015,435(435):200 ~ 206.
[7]Xu W N,Liu W B,Shen M,et al. Effect of different dietary protein and lipid levels on growth performance,body composition of juvenile red swamp crayfish(Procambarus clarkii)[J]. Aquaculture International,2013,21(3):687 ~ 697.
[8]陈乃松,肖温温,梁勤朗,等.饲料中脂肪与蛋白质比对大口黑鲈生长,体组成和非特异性免疫的影响[J]. 水产学报,2012,36(8):1270~1280.
[9]Du Z Y,Clouet P,Huang L M,et al. Utilization of different dietary lipid sources at high level in herbivorous grass carp(Ctenopharyngodon idella):mechanism related to hepatic fatty acid oxidation[J]. Aquaculture Nutrition,2008,14(1):77~ 92.
[10]李如亮.生物化学实验[M].武汉:武汉大学出版杜,1998.
[11]Y. HU,Tan B,Mai K,et al. Growth and body composition of juvenile white shrimp,Litopenaeus vannamei,fed different ratios of dietary protein to energy[J]. Aquaculture Nutrition. 2008,14(6):499~506.
[12]Catacutan M R. Growth and body composition of juvenile mud crab,Scylla serrata,fed different dietary protein and lipid levels and protein to energy ratios[J]. Aquaculture,2002,208(1):113 ~ 123.
[13]Lee S M,Jeon I G,Lee J Y. Effects of digestible protein and lipid levels in practical diets on growth,protein utilization and body composition of juvenile rockfish(Sebastes schlegeli)[J].Aquaculture,2002,211(1~ 4):227~ 239.
[14]李贵锋,蒋广震,刘文斌,等.不同蛋白质和能量水平对建鲤幼鱼生长性能、体组成和消化酶活性的影响[J].上海海洋大学学报,2012,21(2):225~ 232.
[15]Ai Q,Mai K,Li H,et al. Effects of dietary protein to energy ratios on growth and body composition of juvenile Japanese seabass,Lateolabrax japonicus.[J]. Aquaculture,2004,230(1):507 ~ 516.[16]于宁,朱站英,冯文和,等. 克氏原螯虾饲料最适能量蛋白质比 [J].动物营养学报,2014,26(4):1111~ 1119.
[17]Tibbetts SM,Lall SP,Milley JE. Effects of dietary protein and lipid levels and DP DE-1 ratio on growth,feed utilization and hepatosomatic index of juvenile haddock,Melanogrammus aeglefinus L[J]. Aquaculture Nutrition,2015,11(1):67~ 75.
[18]Ali M Z,Jauncey K. Approaches to optimizing dietary protein to energy ratio for African catfish Clarias gariepinus(Burchell,1822)[J].Aquaculture Nutrition,2015,11(2):95 ~ 101.
[19] 虞冰如,沈竑. 日本沼虾饲料最适蛋白质、脂肪含量及能量蛋白比的研究[J].水产学报,1990,14(4):321~327.
[20] Sánchez D R,Fox J M,Iii D G,et al. Dietary effect of fish oil and soybean lecithin on growth and survival of juvenile Litopenaeus vannamei in the presence or absence of phytoplankton in an indoor system[J].Aquaculture Research,2014,45(8):1367~ 1379.
猜你喜欢
英语世界(2022年9期)2022-10-18 01:11:24
当代水产(2022年7期)2022-09-20 06:23:00
当代水产(2022年6期)2022-06-29 01:12:02
保健医苑(2022年4期)2022-05-05 06:11:14
当代水产(2021年8期)2021-11-04 08:49:10
肝博士(2021年1期)2021-03-29 02:32:10
小哥白尼(趣味科学)(2020年1期)2020-06-16 03:24:44
中成药(2019年12期)2020-01-04 02:02:24
中成药(2018年7期)2018-08-04 06:04:18
海洋与渔业(2017年11期)2018-01-26 01:44:58
