
内蒙古克什克腾旗典型草原区达乌尔黄鼠繁殖起始时间与体重(年龄)的关系
2018-06-26 07:56邹永波锦哈斯宝力道
野生动物学报 2018年2期
唐 宇 邹永波 赵 栋 刘 亮 刘 锦哈斯宝力道 图 雅 郭 聪*
(1.四川大学生命科学学院,成都,610064;2.中国科学院动物研究所农业虫鼠害综合治理研究国家重点实验室,北京,100101;3.内蒙古锡林郭勒盟林业局,锡林浩特,026000)
繁殖是动物生存的重要过程之一,繁殖能力直接影响动物的种群数量和种群动态[1],动物种群密度也影响着其繁殖[2-3],动物繁殖的相关问题一直倍受学者关注[4],对影响动物繁殖因素的探索也是研究的重点。Taylor等[5]研究发现,一些非洲鼠类在雨季开始后才开始繁殖,并推测繁殖起始时间与植被变化所带来的食物改变有关。进一步的研究表明,饲喂植物中提取的6-MBAO可以促进雌性西岸田鼠(Microtus townsendii) 的繁殖[6]。Wauters 等[7]对松鼠(Sciurus vulgaris)的研究则发现,雌性个体只有达到一定体重(300 g)才会发情,且体重越大的个体繁殖成功的概率越高。李春等[8]通过对小熊猫(Ailurus fulgens)粪样的分析,认为雌二醇和孕酮是促使雌性小熊猫繁殖启动的重要因素。环境的变化也会影响动物的繁殖。已有研究发现,光周期的变化可以通过影响褪黑激素等激素的分泌来调控动物季节性繁殖[9]。周延山等[1]分析前人研究,认为温度会影响鼢鼠亚科(Myospalacinae)的繁殖启动。
达乌尔黄鼠(Spermophilus dauricus)为松鼠科(Sciuridae)黄鼠属的一类啮齿动物,广泛分布于我国东北、华北和内蒙古等地的平原和沙漠地区[10]。达乌尔黄鼠是昼行性鼠种,冬季会蛰入洞中冬眠,繁殖期集中在4月,每年繁殖1次,寿命最长可达7 a[11]。雌性达乌尔黄鼠第一年越冬后即可参与繁殖[12],每年仅繁殖1次,孕期28 d左右。4月为达乌尔黄鼠繁殖高峰期,到5月中旬交配期已接近尾声[13]。食性以植物为主[14],对草原草场造成破坏,同时达乌尔黄鼠还是鼠疫的自然疫源宿主[15],是重要的草原害鼠[16],在生态系统中有重要地位[17]。
目前国内对达乌尔黄鼠繁殖起始时间的相关研究较少。为了深入了解达乌尔黄鼠早春的繁殖情况,2006年作者在内蒙古典型草原区开展相关实验,研究了达乌尔黄鼠繁殖起始时间与其身体各指标参数(体重、胴体重、体长等)的关系,增加了对其种群数量和种群动态变化的了解,以期为鼠害和疾病防控提供参考。
1 材料与方法
1.1 研究地点
研究区域位于内蒙古赤峰市克什克腾旗阿其乌拉苏木(N43°26',E116°46',海拔 1250 m),属半干旱草原气候[18],植被类型以典型草原为主。样地内的主要植物有羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)、冷蒿(Artemisia frigida)、寸草(Carex duriuscula)、菊叶委陵菜(Potentilla tanacetifolia)和二裂委陵菜(Potentilla bifurca)等。实验样地为开放的自然样地,样地内达乌尔黄鼠分布较多且活动频繁,此外还有少量其他啮齿类如黑线仓鼠(Cricetulus barabensis)、五趾跳鼠(Allactaga sibirica)、蒙古黑线毛足鼠(Phodopus Sungorus)等栖居。这些啮齿动物的活动吸引了较多捕食者,主要有苍鹰(Accipiter gentilis)、长耳鸮(Asio otus)等猛禽及艾鼬(Mustela eversmanii)、沙狐(Vulpes corsac)等兽类[19]。
1.2 取样方法
2006年5月10~19日期间,以花生米为诱饵,采用夹线法对样地内的鼠类进行夹捕获取实验样本。一共布放18条夹线,相邻两条夹线之间间隔50 m,夹线内相邻2只鼠夹之间间隔10 m,每条夹线放置100只鼠夹,共放置1800夹。每天检查4次收取捕获的鼠类,每2 d将鼠夹挪移位置,连续夹捕10 d。对捕获的每只达乌尔黄鼠进行编号,测量记录样本的体长、体重、性别和雄鼠的繁殖状态(睾丸是否下降),再进行常规解剖,称量胴体重,同时观察雌鼠的繁殖状态,记录胎仔数和胚胎发育等级[19]。
1.3 达乌尔黄鼠妊娠时间的确定
张知彬等[16]提出了常用的5级胚胎分级系统,本研究在此基础上增补了黄体的判别。黄体和各级胚胎所对应的妊娠时间及据此推测的繁殖起始时间见表1。各级繁殖起始时间为对应妊娠时间平均值,表示于多少天之前开始妊娠。
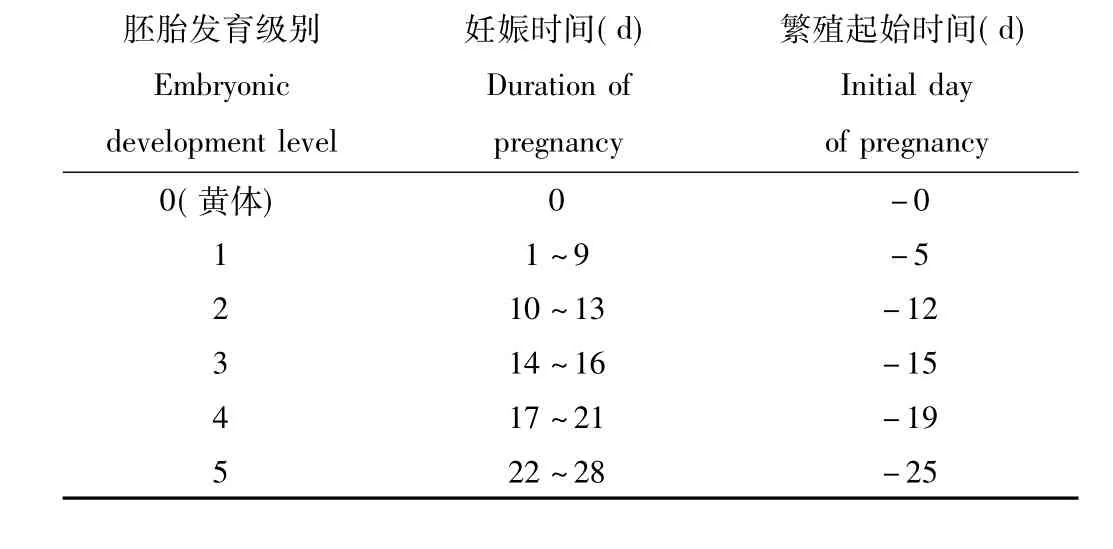
表1 达乌尔黄鼠雌鼠胚胎发育级别与妊娠时间推算Tab.1 The calculate table of embryonic development level and corresponding duration of pregnancy for Spermophilus dauricus
1.4 达乌尔黄鼠雌鼠的体重、胴体重、体长与繁殖起始时间
实验期间共捕获48只雌性达乌尔黄鼠。分别解剖测量记录体重、胴体重、体长数据,观察记录胚胎发育级别,并依据表1所列的对应关系推算每个个体的繁殖起始时间。为了便于统计分析,将黄鼠最早繁殖起始时间作为0点,在此基础上繁殖起始时间越晚,则对应的数值越大,所得数据详见表2。
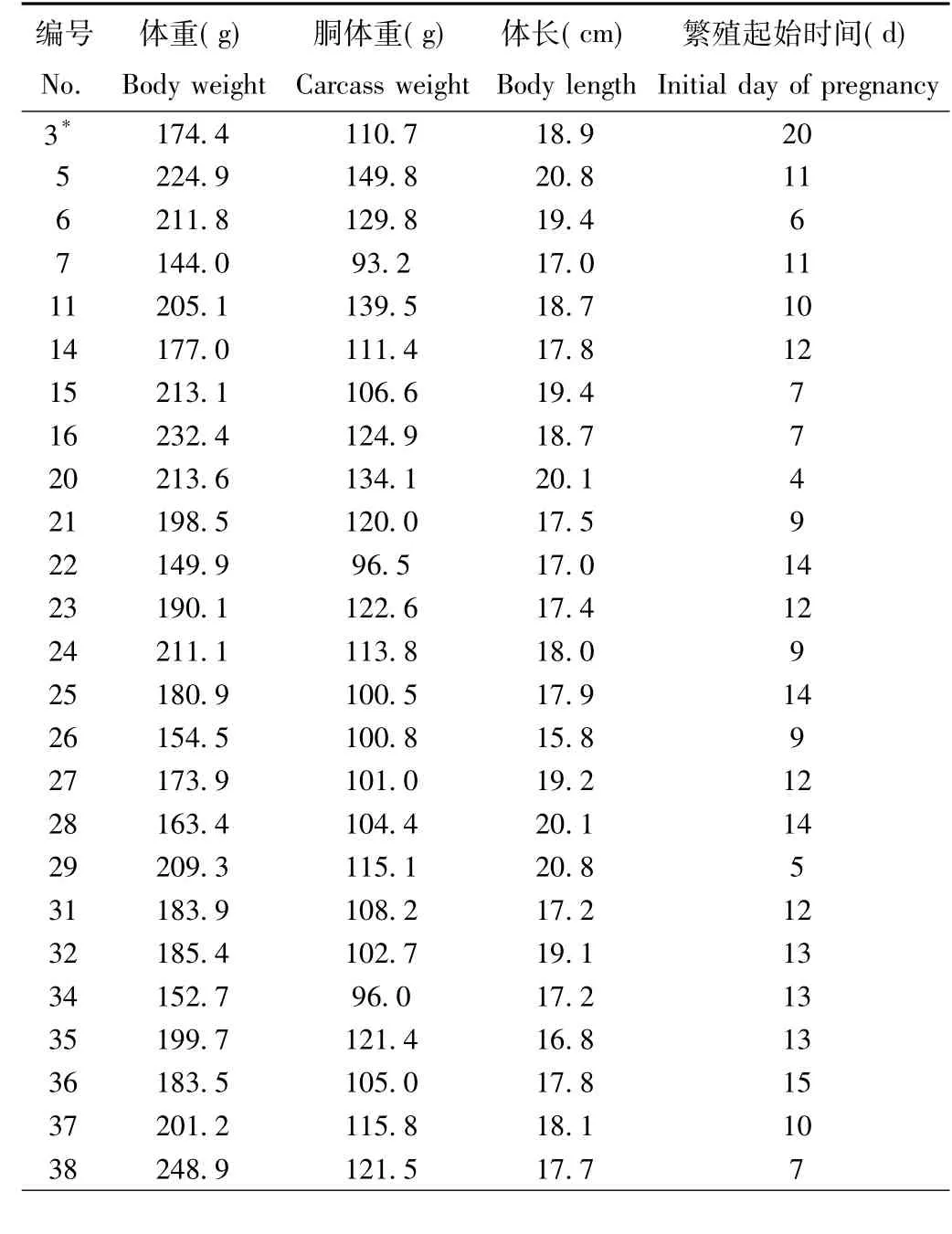
表2 达乌尔黄鼠雌鼠的体重、胴体重、体长与繁殖起始时间Tab.2 The body weight,carcass weight,body length and initial day of pregnancy for female Spermophilus dauricus
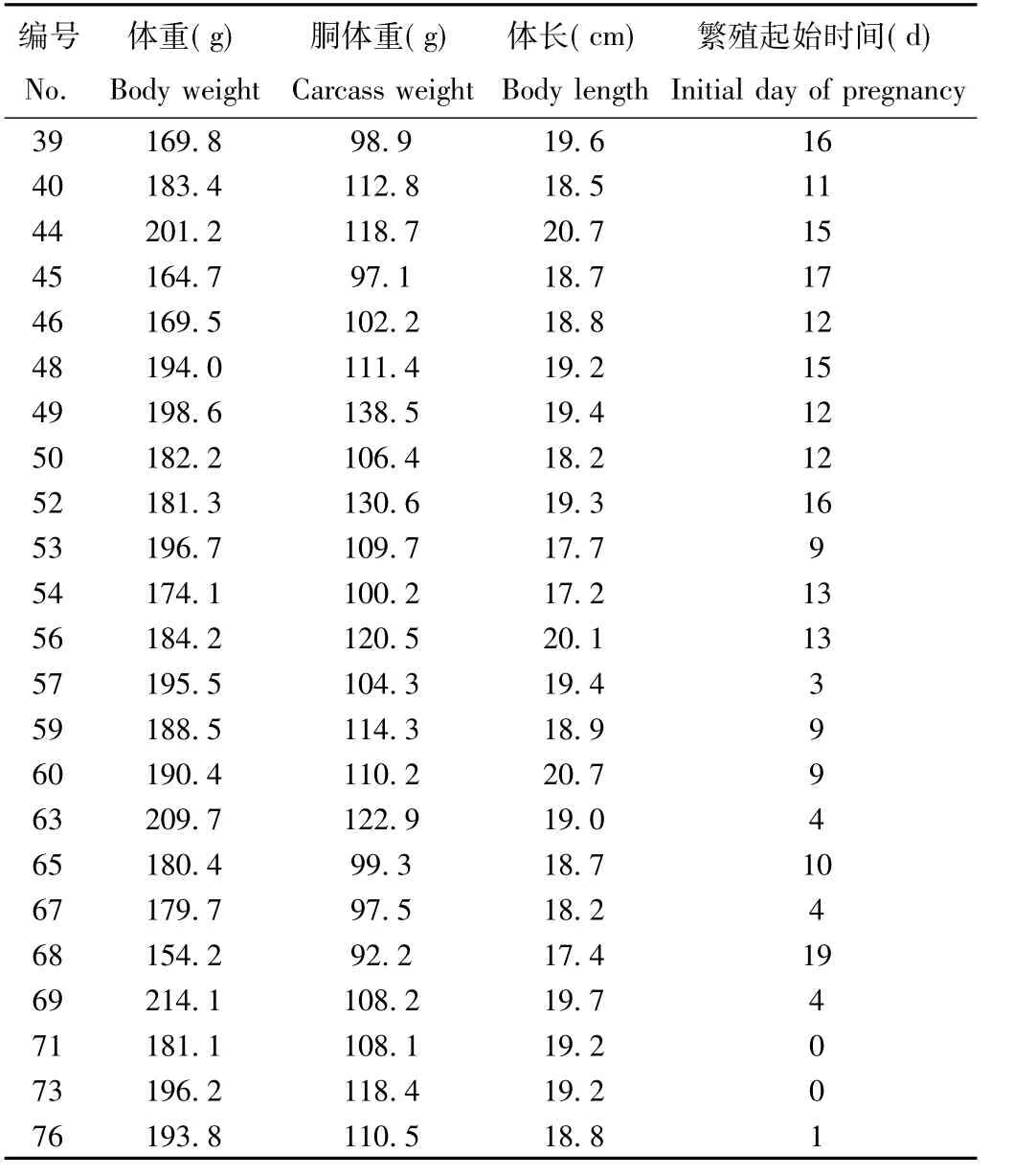
续表2
1.5 数据分析
应用R3.3.2(R Core Team 2015)软件,采用回归进行分析拟合,虚线内表示95%的置信区间。
2 结果与分析
2.1 达乌尔黄鼠繁殖起始时间与体重、胴体重的关系
对表2中体重数据与对应的繁殖起始时间进行分析拟合,拟合结果如图1所示。
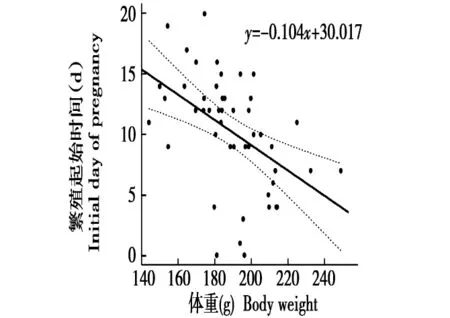
图1 体重与繁殖起始时间的关系Fig.1 The relationship between body weight and initial day of pregnancy
从图1可以看出体重和繁殖起始时间存在线性关系,y=-0.104x+30.017(r= -0.480,P<0.05),同时可以看出,繁殖起始时间与体重趋势存在吻合,即随体重的上升繁殖起始时间也变早。
考虑到妊娠期间,雌鼠会大量进食以保证胎儿营养,体重会相应增加,而繁殖起始时间早的个体,胚胎发育时间更长,胚胎重量也就越重,导致雌鼠的体重相应也较大。故仅用体重与繁殖起始时间做分析可能不甚准确。胴体重表示去掉各内脏器官后的体重,利用胴体重分析可以排除繁殖状况和胃容物等的干扰[20],因此,对繁殖起始时间与对应的胴体重数据做非线性拟合,所得结果如图2所示。
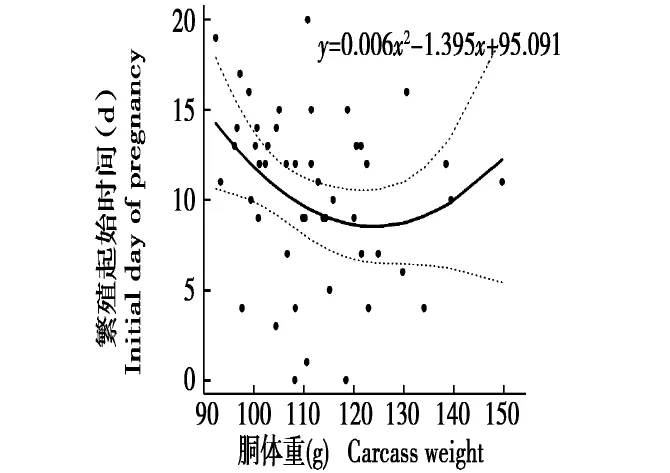
图2 胴体重与繁殖起始时间的关系Fig.2 The relationship between carcass weight and initial day of pregnancy
从图2可以看出,胴体重和繁殖起始时间存在非线性相关关系,y=0.006x2-1.395x +95.091(r=-0.213,P<0.1)。当胴体重小于等于116 g时,随着胴体重的上升繁殖起始时间变早;当胴体重大于116 g时,随着胴体重的上升繁殖起始时间变晚。由此可知,胴体重越接近116 g的雌鼠繁殖时间越早,胴体重偏大或偏小的雌鼠繁殖时间都越晚。
在啮齿动物研究中,常把体重和胴体重作为进行年龄划分的指标[21],张美文等[22]认为,体重和胴体重能反映该啮齿动物的生长发育和繁殖状况。王也等[23]发现,不同年龄组的三趾跳鼠(Dipus sagitta)体重有显著的差异,且体重与年龄呈正相关。因此,体重能准确地反映其年龄,体重越大的个体通常年龄也就越大,因此可以得到,年龄越大,繁殖时间越早。
2.2 达乌尔黄鼠繁殖起始时间和体长的关系
对表1中体长数据与繁殖起始时间进行分析,采用线性回归对实验数据拟合,结果如图3所示。
从图3中可以看出,体长和繁殖起始时间不存在显著的线性关系,y=-0.779x+24.788(r=-0.191,P>0.1)。从图3可以看出,体长越长的个体,繁殖起始时间越早,体长和繁殖起始时间趋势存在吻合,但无显著性。
体长是反映动物身体状况的一个指标,陈立军等[24]研究达乌尔鼠兔(Ochotona dauurica)发现,幼体鼠兔的体长随着年龄增长而增加,但亚成体和成体鼠兔的体长则基本不再增加。对于幼体,体长能较好地反映其年龄,但对亚成体和成体体长则无法作为有效的年龄参考,此外,杨荷芳[25]提出,小型动物的身体肌肉和软组织部分具有很大的伸缩性,在实际测量中会存在很大的人为误差。因此,体长可以作为达乌尔黄鼠年龄划分的参考指标,但不宜作为主要指标。
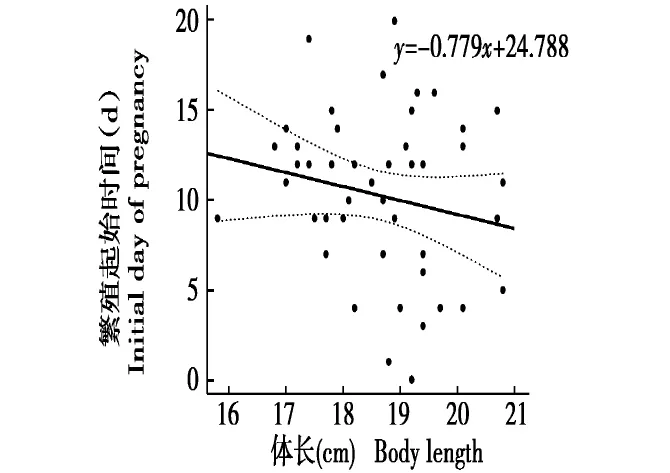
图3 体长与繁殖起始时间的关系Fig.3 The relationship between body length and initial day of pregnancy
3 讨论
达乌尔黄鼠系冬眠鼠,其出蛰时间雌雄具有差异性,费荣中等[13]研究表明达乌尔黄鼠雄鼠出蛰时间早于雌鼠,但不同年龄个体没有顺序规律,达乌尔黄鼠雄性睾丸下降时间集中于4月中旬至5月中旬。本文的结果表明雌鼠繁殖起始时间集中在4月下旬到5月上旬之间,稍晚于雄性繁殖时间,与费荣中等[13]关于达乌尔黄鼠雄性的繁殖时间相符。由于达乌尔黄鼠雄性先出蛰,雌性后出蛰,可见繁殖起始时间的差异主要由雌性黄鼠的生理状态决定。
3.1 繁殖起始时间与营养需求的关系讨论
本文揭示了雌性达乌尔黄鼠体重与早春繁殖起始时间的关系,数据分析的结果表明,雌鼠的体重越大,繁殖起始时间就越早,且排除了繁殖状况的胴体重数据亦是如此。黄鼠为冬眠物种,抵抗冬季的寒冷对能量消耗极大,野外观察发现,刚出蛰的黄鼠往往非常瘦弱,需要大量摄食恢复体能。我们推测,雌鼠繁殖时间的差异可能与其营养状态有关,营养物质积累得多的个体越早达到繁殖的要求。小型哺乳动物的繁殖是非常复杂的过程,繁殖期对能量的需求极高[26],高原鼠兔(Ochotona curzoniae)在繁殖期的能量摄入会比平时增加30%,而中华鼢鼠(Myospalax fontanieri)会增加50%[27]。动物摄入的能量,一部分用于维持自己的生存需要,一部分用于繁殖,这个过程其实就是分配给生长与繁殖的能量之间的权衡。食物、温度、降水和光周期是影响哺乳动物繁殖的主要环境因素[28],选择在环境条件适宜,食物丰富的时间繁殖也是长期自然选择的结果。4月中旬出蛰时温度回升,植物陆续发芽返青,食物逐渐充足,对于雌性达乌尔黄鼠来说,出蛰后需要较长时间的觅食,此时胃中可见大量绿色植物及昆虫碎片[14],表明黄鼠需要储备足够的营养用于妊娠期和随后的哺乳期,体重大的个体先满足繁殖需要的能量条件,可以较早进入妊娠期。
3.2 繁殖起始时间与年龄的关系
胴体重法在小型哺乳动物年龄鉴定方面应用比较广泛。陈立军等[24]利用胴体重法将达乌尔鼠兔分为3个年龄组,各年龄组之间差异均极显著,王也等[23]也用此方法对三趾跳鼠进行年龄划分,划分结果也比较准确。刘加坤等[29]研究了达乌尔黄鼠的年龄结构,以胴体重为量度将达乌尔黄鼠种群分为3个年龄组,说明用胴体重划分年龄比较有参考意义,胴体重越大的个体对应的年龄也越大。本实验发现,胴体重越大的个体繁殖起始时间越早,可认为年龄大的个体繁殖起始时间稍早。邹波等[30]在研究花鼠(Tamias sibiricus)的繁殖特征时,发现花鼠达到性成熟所需时间为10个月。使得年龄Ⅰ组的花鼠繁殖起始时间偏后。达乌尔黄鼠在分类上与花鼠较近,作者认为达乌尔黄鼠同样存在相似现象,详细原因有待进一步研究。
致谢:野外工作得到内蒙古草原动物生态研究站工作人员的协助和支持,在此一并致谢。
[1] 周延山,花立民,纪维红,等.鼢鼠亚科繁殖特性研究进展[J].草业科学,2015,32(6):1010-1016.
[2] 韩群花,郭聪,张美文.密度制约效应对啮齿动物繁殖的影响[J].生态学报,2013,33(19):5981-5989.
[3] 刘蓓蓓,王广鑫,许林,等.阔叶红松林下花鼠的种群动态[J].野生动物学报,2015,36(4):391-394.
[4] 贾义平,靳伟,左之才,等.赤腹松鼠血清电解质的测定及性别与季节差异[J].野生动物学报,2017,38(1):17-21.
[5] Taylor K D,Green M G.The influence of rainfall on diet and reproduction in four African rodent species [J].Journal of Zoology,1976,180(3):367-389.
[6] Korn H,Taitt M J.Initiation of early breeding in a population of Microtus townsendii(Rodentia)with the secondary plant compound 6-MBOA [J].Oecologia,1987,71(4):593 -596.
[7] Wauters L,Dhondt A A.Body weight,longevity and reproductive success in red squirrels(Sciurus vulgaris) [J].Journal of Animal E-cology,1989,58(2):637 -651.
[8] 李春,魏辅文,胡锦矗.雌性小熊猫粪样中雌二醇与孕酮水平的变化与繁殖启动的关系 [J].动物学研究,2005,26(2):147-151.
[9] 黄冬维,储明星.动物季节性繁殖分子调控机理研究进展[J].遗传,2011,33(7):695-706.
[10] 张赫武.达乌尔黄鼠种群年龄研究 [J].动物学杂志,1977(3):33-34.
[11] 范志勤,廖崇惠.温度对达乌尔黄鼠冬眠的影响[J].生态学报,1982,2(4):362,374.
[12] 罗明澍,钟文勤.达乌尔黄鼠种群生态的一些资料[J].动物学杂志,1990,25(2):50-54,60.
[13] 费荣中,李景原,商志宽,等.达乌尔黄鼠的生态研究 [J].动物学报,1975,21(1):18-29.
[14] 罗明澍.达乌尔黄鼠的食性研究 [J].动物学报,1975,21(1):62-70.
[15] Xu Lei,Schmid B V,Liu Jun,et al.The trophic responses of two different rodent-vector-plague systems to climate change[J].Proceedings of the Royal Society of London B:Biological Sciences,2015,282(1800):20141846.
[16] 张知彬,王祖望.农业重要害鼠的生态学及控制对策[M].北京:海洋出版社,1998:242-243.
[17] 张美文,王凯荣,王勇,等.洞庭湖区鼠类群落的物种多样性分析 [J].生态学报,2003,23(11):2260-2270.
[18] 刘振国,李镇清.不同放牧强度下冷蒿种群小尺度空间格局[J].生态学报,2004,24(2):227-234.
[19] 刘亮.典型草原区达乌尔黄鼠的空间利用模式[D].北京:中国科学院动物研究所,2007.
[20] 张洁.北京地区黑线仓鼠年龄鉴定及种群年龄组成的研究[J].兽类学报,1985,5(2):141-150.
[21] 杨再学,周朝霞,潘世昌,等.应用胴体重指标鉴定黄胸鼠的年龄 [J].贵州农业科学,2010,38(3):110-113.
[22] 张美文,郭聪,王勇,等.洞庭平原黄胸鼠种群年龄组的划分[J].兽类学报,1998,18(4):268-276.
[23] 王也,张文杰,任娅茹,等.浑善达克沙地三趾跳鼠体重与年龄划分[J].动物学杂志,2014,49(6):798-803.
[24] 陈立军,刘伟,苏永志,等.典型草原区达乌尔鼠兔年龄划分标准 [J].动物学杂志,2013,48(3):345-350.
[25] 杨荷芳.小型兽类年龄鉴定方法简评 [J].生态学杂志,1990,9(2):54-55.
[26] 刘赫,王德华,王祖望.小型哺乳动物繁殖期的能量收支对策[J].兽类学报,2001,21(4):301-309.
[27] 王祖望,曾缙祥,韩永才,等.高山草甸生态系-小哺乳动物能量动态的研究Ⅰ.高原鼠兔和中华鼢鼠对天然食物的消化率和同化水平的测定 [J].动物学报,1980,26(2):184-195.
[28] Bronson F H.Mammalian reproduction:an ecological perspective[J].Biology of Reproduction,1985,32(1):1 -26.
[29] 刘加坤,王廷正,李金钢,等.达乌尔黄鼠种群年龄结构的研究 [J].兽类学报,1993,13(4):277-282.
[30] 邹波,宁振东,王庭林,等.花鼠种群生态学研究(Ⅱ)-繁殖特征[J].陕西师范大学学报:自然科学版,1997(S1):108-111.
猜你喜欢
能源研究与信息(2022年3期)2022-10-29
中国牛业科学(2020年4期)2020-08-15
做人与处世(2020年10期)2020-06-29
中外文摘(2019年20期)2019-10-24
科技创新与品牌(2018年5期)2018-07-24
新农村(2018年35期)2018-04-02
文史博览·文史(2015年8期)2015-08-18
电影(2015年12期)2015-01-05
食品工业科技(2014年5期)2014-03-11
中国青年(1954年3期)1954-08-29
