
贺兰山马麝夏季营养采食策略
2018-06-26 07:56刘尊显滕丽微孙玉姣王志勇王继飞马忠其刘振生
野生动物学报 2018年2期
赵 唱 刘尊显 滕丽微,3 高 惠 孙玉姣王志勇 王继飞 马忠其 刘振生,3*
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.黑龙江三江国家级自然保护区管理局,抚远,156500;3.国家林业局野生动物保护学重点实验室,哈尔滨,150040;4.宁夏贺兰山国家级自然保护区管理局,银川,750021;5.银川市绿化一处,银川,750001)
“营养”一词,最早被提出用来衡量生物体与自然环境,各生物体之间相互作用的物质[1]。食物为有机体提供能量,而能量的传递随着食物链从被捕食者传递到捕食者中[2],这对生物体的进化来说至关重要,对食草动物更是如此。营养生态学最基本任务就是来确定该动物在某种时空条件下达到最适时对不同营养成分的需求,即确定该动物的营养选择策略[3]。动物随着环境改变,在不同季节对营养的需求不同,因植物本身随着季节的变化,自身的营养成分也会发生变化,因此动物会在众多的食物中进行选择,以便更好地适应环境,更好生存[4]。
在营养生态学目前的研究结果中,普遍认为动物对食物采食有以下策略:蛋白质摄入最大化[5-6]、能量摄取最大化[7]、采食营养物质均衡化[8]以及回避有毒物质[9]。传统的研究通常是针对单个营养成分因子对动物采食策略进行分析,或仅仅关注蛋白质、能量,亦或关注营养成分中的某种元素进行线性的分析。而在有些情况下,食物各营养成分间的关系并不仅存在于单一线性关系中。
综上所述,本文在研究贺兰山马麝(Moschus sifanicus)种群食物组成的基础上,测量马麝所采食夏季大宗食物中的粗蛋白含量、总能以及植物次生代谢产物单宁含量,通过建立非线性模型来衡量贺兰山地区马麝种群水平对各水平下食物中蛋白质、能量和单宁的相互响应关系,进而得到贺兰山地区马麝夏季采食的营养策略,同时通过建模结果预测马麝夏季对食物中蛋白质和单宁的选择偏好。
1 研究地概况
贺兰山位于内蒙古自然保护区和宁夏回族自治区的交界处(N38°21'~ 39°22',E105°49'~ 106°42'),海拔高度一般为2000~3000 m。具有典型的大陆性季风气候特点。年平均气温-0.7℃,年平均降水日数为94 d,平均降水量420 mm,最低降水量和最高降水量分别为187.6 mm和627.5 mm,年日照时数高达3100 h,全年无霜期仅为124 d。已知的贺兰山野生维管植物共87科357属788种2亚种,28个变种。包括种子植物77科346属2亚种28变种和蕨类植物10科11属18 种[10]。
2 材料方法
2.1 野外样品收集及切片制备
2014~2016年,结合2013年全年布设的红外相机拍摄到的马麝出现位置,在贺兰山马麝活动比较频繁的12条沟段设计样线,所设定的样线覆盖了马麝活动的所有区域。采集野外马麝新鲜粪便,以10~20粒为一份装在信封里,并记录粪便采集时间、地点、生境类型、GPS定位等数据,从所收集的不同粪堆中随机抽取6~10 g粪便混合一起,分别组成独立样本。我们将3 a的夏季粪便样本数据组合在一起,共采集制作独立粪便样本70个,每个样本制作3张切片进行观察,最终共计观察粪便亚显微视野210个。同时采集沟系内的马麝活动区域内的所有啃食的植物样本,分别取每种植物的花、茎、叶、杆等部位,每种植物做相同标记采集2份,将一份放于标本夹中,以便请教专家或查阅书籍进行物种鉴定;另一份装入牛皮纸中密封保存,带回实验室中进行称重及制片处理。
将采集的植物与用于食性分析的粪便样本在60℃烘箱里烘72 h至恒重,用粉碎机粉碎,在40~100目筛中过筛,倒入次氯酸钠并间隔3~5 h制作临时装片观察细胞形态是否清晰,待细胞形态清晰后清洗,滴1~2滴番红花红染色。用镊子夹取碎片置于载玻片上,滴1滴甘油,盖上盖玻片。最后用中性树胶封片并贴上标签待观察。每种植物和粪便样本均制作3张装片贴标签待检。
2.2 植物中单宁含量测定
取风干的植物样本,用液氮将其研磨成粉末状,精准称量0.05 g,加入70%的甲醇溶液5 mL,室温静置24 h,离心10 min,取上层清液0.5 mL,加入4%的香草醛甲醇溶液3 mL,加入浓盐酸1.5 mL,铝箔遮光,摇匀试管。而后进水浴锅加热20 min。在510 nm波长下以试剂空白作对照测量吸光度。以单宁质量浓度(依次为0、0.4、0.8、1.2、1.6、2.0,单位μg/mL)为横坐标,吸光度为纵坐标作图,其中标准样品为儿茶素。
2.3 植物中蛋白质含量测定
采用凯氏定氮法,即在催化剂作用下,用硫酸使得含氮物转化成硫酸铵,之后加入强碱进行蒸馏,使得氨气溢出,用硼酸吸收后,在进行酸性滴定,得出氮含量。
运用公式:
样品中粗蛋白含量(N×6.25)%=(V3-V0)×0.0100×0.014 g×6.25×V1/V2×100/W
式中:W—样品重,g;
N—样本量;
V1—消化液稀释前量,mL;
V2—稀释液蒸馏量,mL;
V3—滴定样品馏出液的0.0100 mol HCL的消耗量,mL;
V0—世纪空白消耗的HCL用量。
2.4 总能的测定
运用奔特WZR-1A氧弹热量计进行能量的测定。首先准确称量1 g样品,放入坩埚中,用氧弹热量计测量热容量,向计算机输入相关参数,试验结束计算机自动计算出热容量,重复试验4~5次,如结果符合条件,则通过计算机计算出平均热容量及极差,得到植物样品的热量值。
2.5 电镜下镜检及定量
统计求得每种植物角质碎片的出现频率F。采用频率转换法,依公式:
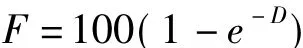
转换为每个视野中每种植物可辨认表皮角质碎片的平均密度D,D又可转换为相对密度RD。方法为:
RD=(每种植物可辨认的表皮角质碎片的密度)/(各种植物可辨认角质碎片的密度之和)100%。
RD即食物中各种植物的干重组成比例。可作为马麝对不同植物采食频率的估计值,据此列出贺兰山马麝夏季食谱及大宗食物。
2.6 数据处理与分析
利用Excel 2013建表,运用SPSS 20.0进行植物聚类分析及差异性显著分析,同时运用Design-Expert 8.0.6软件进行中心合成设计(Central Composite Design,CCD)建立优化响应曲面模型设计包含二阶因子轴向点以及中心点等3组设计点。所有结果中,定义显著性水平P<0.05。
3 结果
3.1 马麝夏季主要食物组分
食性分析结果表明,马麝主要采食12种植物,包括苔藓植物(Bryophytes)、山杏(Prunus sibirica)、密齿柳(Salix characta)、蒙古扁桃(Prunus mongolica)、脚苔草(Carex pediformis)、狭叶锦鸡儿(Caraga-na stenophylla)、高山地榆(Sanguisorba alpina)、紫丝膜菌(Cortinarius purascens)、秦氏黄芪(Astragalus chingianus)、紫丁香(Syringa oblata)、小叶金露梅(Potentilla parvifolia)、内蒙野丁香(Leptodermis ordosica),柱状图表明,在所有植物组分中,该12种植物占比达70%以上,是马麝夏季的主要食物(图1)。
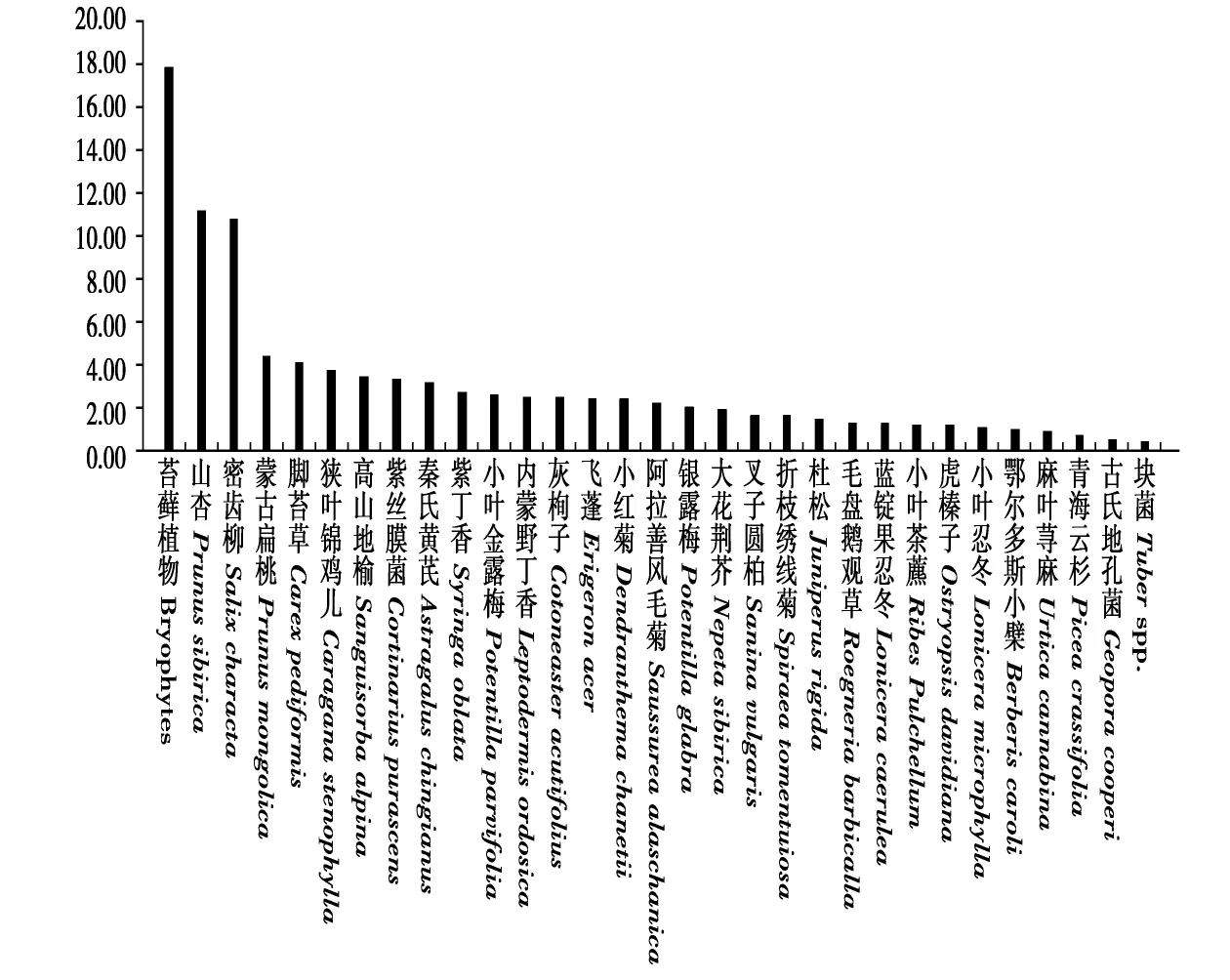
图1 马麝夏季食物组成(%)Fig.1 Diet composition of Alpine musk deer in summer
对贺兰山马麝夏季采食植物组成按RD值以最小距离法进行聚类分析,结果如下。
从聚类分析图中可以得出:苔藓植物单独一类,密齿柳和山杏聚为一类,两类与其他距离较近的种类合成了一类,苔藓植物、山杏、密齿柳共同组成了高山麝夏季的最主要食物。
将马麝采食的所有植物按科别进行归类得出,蔷薇科(Rosaceae)所占比例最高,为28.32%,其他各科按照大小依次为苔藓植物>杨柳科(Salicaceae)>菊科(Compositae)>豆科(Leguminosae)>莎草科(Cyperaceae)>茜草科(Rubiaceae)>丝膜菌科(Cortinariaceae)>柏科(Cupressaceae)>木犀科(Oleaceae)>忍冬科(Caprifoliaceae)>唇形科(Labiatae)>禾本科(Gramineae)>虎耳草科(Saxifragaceae)>桦木科(Betulaceae)>小檗科(Berberidaceae)>块菌科(Tuberaceae)>荨麻科(Urticaceae)>松科(Pinaceae)。取食的植物类别中,灌木总比例为38.79%,草本占22.49%,苔藓植物占18.15%,乔木占16.17%,菌类比例最少,占4.40%(图3)。
3.2 马麝夏季食物中主要成分含量
3.2.1 粗蛋白含量
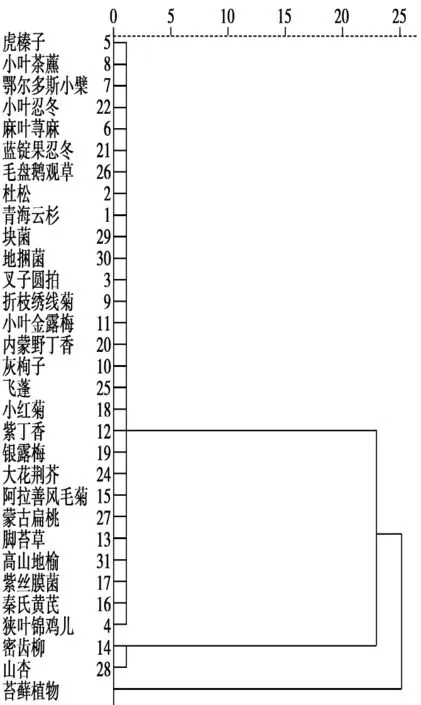
图2 马麝夏季食物组成系统聚类树状图Fig.2 The cluster dendrogram of diet composition of Alpine musk deer in summer
在夏季,粗蛋白含量最高的依然是古氏地孔菌(Geopora cooperi),其含量为35.91%,其次粗蛋白含量大于10%的植物由大到小分别为块菌(Tuber spp.)、紫丝膜菌、小叶金露梅、麻叶荨麻(Urtica cannabina)、银露梅(Potentilla glabra)、虎榛子(Ostryopsis davidiana)、高山地榆、毛盘鹅观草(Roegneria barbicalla)、狭叶锦鸡儿、秦氏黄芪、鄂尔多斯小檗(Berberis caroli),粗蛋白含量分别为:29.26%、25.66%、17.27%、14.91%、14.79%、13.65%、12.42%、11.75%、11.59%、10.67%、10.41%。含量最低的是苔藓植物,其粗蛋白含量为3.54%。从植物类别上划分,冬、夏两季节马麝所食植物中粗蛋白的含量:菌类>木本植物和草本植物>苔藓(图4)。
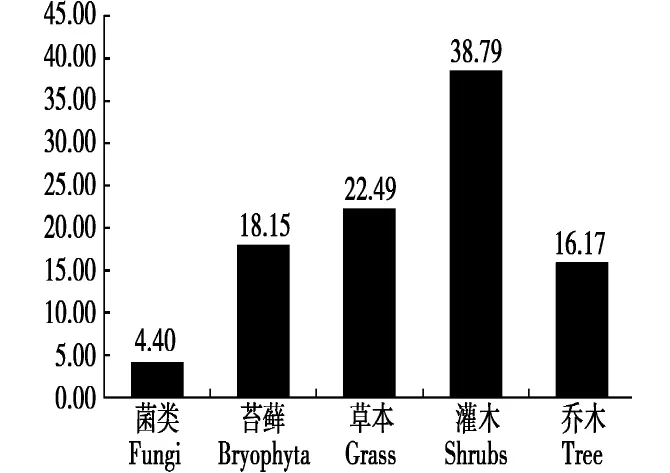
图3 马麝夏季食物不同类别的组成比例(%)Fig.3 Diet composition difference of Alpine musk deer in summer
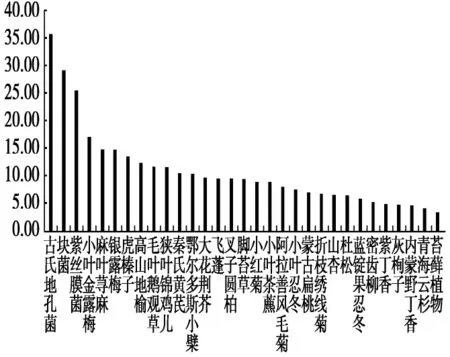
图4 贺兰山马麝夏季食物的粗蛋白含量(%)Fig.4 The crude protein content of food of Alpine musk deer in summer
3.2.2 粗脂肪含量
在夏季,粗脂肪含量最高的是高山地榆,其含量为59.27 g/kg,其次粗脂肪含量大于30%的植物为山杏、秦氏黄芪、蒙古扁桃、银露梅、密齿柳、脚苔草、内蒙野丁香、灰栒子(Cotoneaster acutifolius),含量分别为 56.62 g/kg、54.44 g/kg、41.76 g/kg、41.62 g/kg、39.17 g/kg、36.22 g/kg、32.54 g/kg、31.34 g/kg。含量最低为苔藓植物,其粗脂肪含量为1.07 g/kg。整体来看,木本植物粗脂肪含量最高,其次是菌类,草本植物位列第三,苔藓植物最低(图5)。
3.2.3 单宁含量
在夏季,单宁含量最大的植物为密齿柳,其含量为147.05 g/kg,其次单宁含量排在前十位的植物分别为:虎榛子、蓝锭果忍冬(Lonicera caerulea)、飞蓬(Erigeron acer)、蒙古扁桃、山杏、小红菊(Dendranthema chanetii)、银露梅、阿拉善风毛菊(Saussurea alaschanica)、小叶忍冬(Lonicera microphylla),单宁含量分别为:83.33 g/kg、69.75 g/kg、60.68 g/kg、50.39 g/kg、49.87 g/kg、45.65 g/kg、42.16 g/kg、39.29 g/kg、38.74 g/kg,含量最低的是苔藓植物,其含量为4.79 g/kg。据植物类别分析,木本植物的单宁含量相对较高,草本植物其次,菌类植物第三,而苔藓植物的单宁含量最低(图6)。
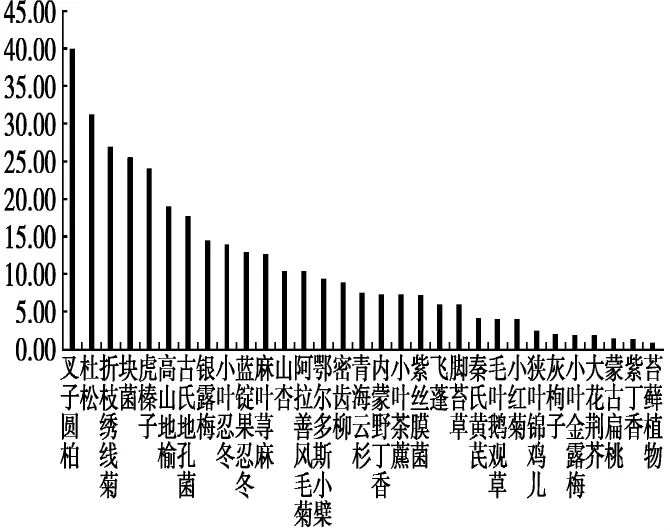
图5 贺兰山马麝夏季食物的粗脂肪含量(g/kg)Fig.5 The ether extract content of food of Alpine musk deer in summer
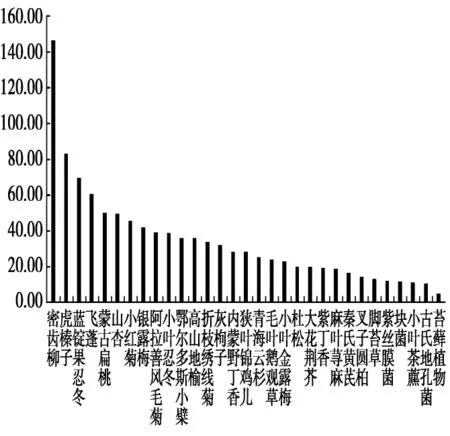
图6 贺兰山马麝夏季食物的单宁含量(g/kg)Fig.6 The tannins content of food of Alpine musk deer in summer
3.3 马麝夏季营养采食策略
在贺兰山马麝夏季粗蛋白、能量、单宁与马麝各食物组分的响应曲面模型结果显示,在粗蛋白与能量曲面中,马麝对于食物采集在能量变化上有着双曲线的变化趋势,一方面,在能量为17070.18~18967 J/g的范围内,马麝更愿意选择高能量低蛋白的植物作为食物,另一方面,在能量范围为11370~17070.18 J/g的范围内,马麝趋向于选择低能量、低蛋白的植物为食物,这说明马麝采食策略趋向于特定的能量与蛋白质比例的食物,但不论采食的食物能量高与低,马麝都趋向于选择低蛋白比例的食物(图7a)。
在粗蛋白和单宁曲面中,马麝趋向于采食高单宁含量的植物,选择粗蛋白含量低的植物,虽然粗蛋白在单宁影响下与食物组分成负相关,但是整体来说,对高含量单宁植物的采食,是该季节马麝对食物采食的策略特点之一,即使将食物组分最高而单宁值含量较低的苔藓植物去掉,马麝依然表现出选择单宁含量高的植物趋势(图7b)。
在能量和单宁曲面中,马麝表现出对能量的一定范围内的选择,一方面,当植物能量在11370~17070.18 J/g的范围内,马麝趋向于取食单宁含量低且能量低的植物,而另一方面,当植物中能量在17070.18~18967 J/g的范围内,马麝趋向于取食能量更高单宁含量更低的植物,这同样说明马麝采食策略趋向于特定的能量与单宁比例食物,但不论采食食物的能量高与低,马麝都更趋向于选择采食单宁含量低的食物(图7c)。对于贺兰山马麝的夏季食物采食策略,蛋白粗、能量、单宁与食物组分的数量关系可表示为三元二次公式(图7d)。
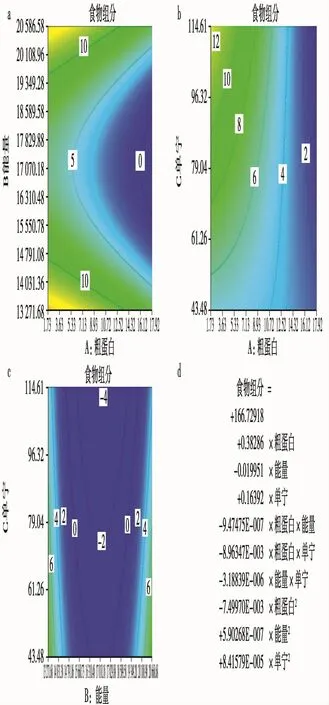
图7 贺兰山马麝夏季食物中粗蛋白、能量和单宁响应曲面模型Fig.7 Response surface model of crude protein,energy and tannins of food of Alpine musk deer in summera.粗蛋白和能量曲面;b.粗蛋白和单宁曲面;c.能量和单宁曲面;d.模型公式a.Surface for crude protein and energy;b.Surface for crude protein and tannins;c.Surface for energy and tannins;d.Model formula
4 讨论
4.1 误差来源及改进策略
粪便亚显微分析方法是基于粪便中可以辨认的植物碎片来确定动物的食物组成[11],其主要目的是检验亚显微组织学能否定性、定量的确定粪便中的食物组成。使用此方法研究有蹄类食草动物的食性时最理想的状态是:(1)每种植物样本的角质化碎片具有同等的辨认率;(2)目标动物对各种植物具有相同的消化率;(3)每份亚显微切片样本中每种植物的每克碎片数和粉碎、筛选后的碎片大小分布均等[12]。显然,在实际操作中,不可能保证上述3种情形达到理想状态,即出现误差。传统方法中使用两组电子显微镜分别对比植物装片和粪便装片并计数的方法会出现随机偶然误差,人为带入某种植物,对实验结果造成一定误差。
粪便亚显微结构分析法分析有蹄类食草动物的食性,核心难度是对粪便亚显微装片以及植物装片中的亚显微结构进行精准对位识别。本研究在完成各装片制作后首先对植物亚显微装片进行试看,并且采用多人多次看同一张切片的方式减少因个人原因造成的计数误差。在制作亚显微装片时,为了防止碎片在制作亚显微切片时出现卷曲和重叠,通常利用眼科镊进行轻微搅拌,选用甘油作为介质,但会加剧碎片更加破碎化,这需要在识别各植物亚显微装片时,找到同种植物所有亚显微装片形态,再对粪便中的碎片进行精准识别。本研究中我们利用番茄红素对植物细胞壁着色,使得细胞形态清晰可见,并且在显微镜上加装照相机对每种植物与粪便亚显微切片进行拍照,利用Photoshop 5.0图形处理软件对切片照片进行局部放大,多重比较,减小误差。我们将马麝夏季主要食物的样本装片与部分易混淆植物的装片列举如下(图8)。

图8 贺兰山马麝夏季主要食物的样本装片Fig.8 Samples of main food of Alpine musk deer in summer
在实验室电镜检验过程中,发现粪便亚显微装片中木本植物与草本植物的细胞形态有许多相似之处,在同等倍数显微镜下分别拍照后,在Photoshop 5.0中同时打开两个相似种植物照片,并添加网格,通过计算每个细胞所占网格数目的大小来量化区分细胞。例如,我们在对蔷薇科的小叶金露梅与禾本科的毛盘鹅观草进行对比时,由于碎片的相似度极高,便通过添加网格计数来计算二者的细胞大小计算比率,而后运用Spss 22.0软件进行差异性分析检验。在Wald-Wolfowitz检验和Mann-Whitney U检验下发现差异性小于0.01属差异显著(图9),而传统鉴别方法中,将植物装片与粪便装片分别在电镜下检验,不能同时观察二者的装片,因此会对数据的精确度产生一定影响。
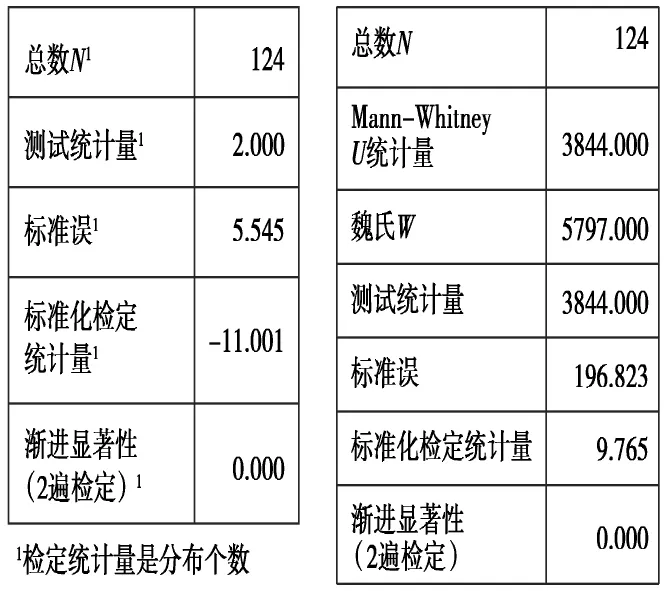
图9 两种植物细胞大小的Wald-Wolfowitz检验和Mann-Whitney U检验结果Fig.9 Nonparametric test of two plant cell size results
4.2 马麝夏季主要食物组分
马麝夏季在贺兰山取食的植物有19科31种(属),其中,食苔藓植物位于马麝食物组分的第一位,这说明,在食物丰富的夏季,马麝对栖息地内植物中的鲜嫩植物需求量极大。从显微镜镜检看,高海拔地区的木本植物和草本植物占了75%以上,与此同时夏季的贺兰山人为活动较为频繁,可能是马麝为躲避人为干扰而移向高海拔地区活动所做出的反应。贺兰山属温带大陆性气候,夏季湿润多雨,且在马麝活动的高海拔地区具备了充足的空间和光照,草本植物和灌木生长茂盛,马麝在此时期可大量获得更多的幼枝嫩叶,因此杨柳科、苔藓植物、豆科有着明显的采食优势。
4.3 马麝夏季的采食策略
食草动物在野外实际采食过程中,经常会遇到不均衡的营养而迫使其必须增加某种食物的摄取量[13-15],从而达到营养互补均衡。Smith的研究表明,当某些营养成分达不到动物的生理需求时,动物为了其健康稳定的生长,就会通过自身的转化作用来合成身体生长所需要的营养物质,而Smith通过长时间的实验得出,蛋白质和能量是草食动物生存最主要的两大营养成分,主导着动物自身的营养状况[16]。而植物中的单宁含量是有蹄类食草动物趋向于回避的一个因素,大量采食单宁高的食物会和食物中的蛋白质结合生成不溶于水的化合物,从而影响对蛋白质的消化[17-20]。同时,对于植物次生代谢产物也要考虑到食物营养成分中,Jones等的研究表明,密西西比白尾鹿(Odocoileus virginianus)中,单宁含量每增加1%,种群食物的体外干物质减少量1.9%,并且蛋白质消化率减少2.5%[21]。从袁喜才、McArt等国内外学者的研究也可以看出,国内外北方地区食草动物采食的植物中含有双子叶木本植物,其单宁含量非常丰富[22-25]。由此可见,单宁对贺兰山马麝种群的营养状况不容忽视。因此,我们在对马麝的食物营养策略研究建模时,将单宁含量加入到蛋白质和能量的模型中,会更好地反映出马麝的采食策略及食物中的营养状态。
蛋白质对于动物来说是最基本最重要的营养元素之一,它影响着动物的生长、发育、繁殖。而能量是动物体生命活动的基础,从新陈代谢到能量交换,整个生命体的基本活动中无时无刻不在发生着能量的生成与代谢过程,马麝从出生、生长,到细胞更新、代谢,营养物质的消化与吸收,都需要能量[26]。而作为植物中的次生代谢产物之一,单宁含量是影响马麝食物适口性重要的影响因素。Schwartz、Cooper等研究都表明,单宁类植物中的次生代谢产物都会降低食草动物的采食频率[27-29],苔藓类植物的单宁含量最低,分布最广,在食物丰富的夏季,对于马麝而言,大量的采食苔藓植物不论从可获得性还是适口性都是不错的选择。在对粗脂肪的测定中我们发现,马麝依然对脂肪的需求显著,而通过建模比较发现,马麝夏季随食物组分的增加对粗蛋白、能量需求减少,这说明马麝在矿质元素,水分需求显著的前提下,可以通过摄取粗脂肪更高含量的植物来补充自身能量的需求。同时,程建国等研究结果说明,在圈养马麝时投入高蛋白的食物会影响雄性马麝泌香期激素分泌减少,从而导致泌香减少[30],而贺兰山马麝每年泌香旺盛期正值6~8月的夏季,低蛋白和低能量的摄取可以保证野生马麝泌香的顺利进行,这也是野生马麝夏季采食策略之一。而雌性马麝每年6月都会进入哺乳期照顾幼崽,对可获得性的食物有着更高的需求。综上,我们可以得出,贺兰山马麝夏季对食物的选择是由多因素共同作用而非单一因素决定的结果,同时也符合绝大多数有蹄类食草动物的夏季采食策略。
[1] Westoby M.An analysis of diet selection by large generalist herbivores[J].The American Naturalist,1974,108(961):290 -304.
[2] Schmitz O J.Herbivory from individuals to ecosystems[J].Annual Review of Ecology,Evolution,and Systematics,2008,39:133 -152.
[3] Simpson S J,Sibly R M,Lee K P,et al.Optimal foraging when regulating intake of multiple nutrients [J].Animal Behaviour,2004,68(6):1299-1311.
[4] Barboza P S,Hartbauer D W,Hauer W E,et al.Polygynous mating impairs body condition and homeostasis in male reindeer(Rangifer tarandus tarandus) [J].Journal of Comparative Physiology B,2004,174(4):309-317.
[5] Mattson Jr W J.Herbivory in relation to plant Nitrogen content[J].Annual Review of Ecology and Systematics,1980,11(1):119 -161.
[6] White T C R.The inadequate environment:nitrogen and the abundance of animals[M].Berlin:Springer-Verlag,1993.
[7] Schoener T W.Theory of feeding strategies[J].Annual Review of Ecology and Systematics,1971,2(1):369 -404.
[8] Raubenheimer D,Simpson S J,Mayntz D.Nutrition,ecology and nutritional ecology:toward an integrated framework[J].Functional Ecology,2009,23(1):4-16.
[9] Dearing M D,Foley W J,McLean S.The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates[J].Annual Review of Ecology,Evolution,and Systematics,2005,36:169 -189.
[10] 杜茜,闫兴富.贺兰山植被类型多样性及空间分布特征[J].安徽农业科学,2010,38(7):3666-3667.
[11] Baumgartner L L,Martin A C.Plant histology as an aid in squirrel food-habit studies [J]. The Journal of Wildlife Management,1939,3(3):266-268.
[12] Holechek J L,Vavra M,Pieper R D.Botanical composition determination of range herbivore diets:a review [J].Journal of Range Management,1982,35(3):309 -315.
[13] Simpson S J,Sword G A,Lorch P D,et al.Cannibal crickets on a forced march for protein and salt[J].Proceedings of the National Academy of Sciences of the United States of America,2006,103(11):4152-4156.
[14] Simpson S J,Raubenheimer D,Charleston M A,et al.Modelling nutritional interactions:from individuals to communities[J].Trends in Ecology& Evolution,2010,25(1):53-60.
[15] Schulkin J.Sodium hunger:the search for a salty taste [J].Quarterly Review of Biology,2005(17):1249-1250.
[16] Smith J M.Optimization theory in evolution [J].Annual Review of Ecology and Systematics,1978,9(1):31 -56.
[17] Cleve K V,Oliver L,Schlentner R,et al.Productivity and nutrient cycling in taiga forest ecosystems[J].Canadian Journal of Forest Research,1983,13(5):747-766.
[18] Rohrs-Richey J K,Mulder C P H.Effects of local changes in active layer and soil climate on seasonal foliar nitrogen concentrations of three boreal forest shrubs[J].Canadian Journal of Forest Research,2007,37(2):383-394.
[19] Aphalo P J,Lahti M,Lehto T,et al.Responses of silver birch saplings to low soil temperature [J].Silva Fennica,2006,40(3):429-442.
[20] Robbins C T,Hanley T A,Hagerman A E,et al.Role of tannins in defending plants against ruminants:reduction in protein availability[J].Ecology,1987,68(1):98-107.
[21] Jones P D,Rude B,Muir J P,et al.Condensed tannins'effect on white-tailed deer forage digestibility in Mississippi[J].Journal of Wildlife Management,2010,74(4):707 -713.
[22] 袁喜才,刘晓明,王骏,等.海南坡鹿食性的研究[J].东北林业大学学报,1990,18(1):66-71.
[23] 崔多英,刘振生,王小明,等.贺兰山马鹿冬季食性分析[J].动物学研究,2007,28(4):383-388.
[24] Ayres M P,Clausen T P,MacLean S F,et al.Diversity of structure and antiherbivore activity in condensed tannins [J].Ecology,1997,78(6):1696-1712.
[25] McArt S H,Spalinger D E,Collins W B,et al.Summer dietary nitrogen availability as a potential bottom-up constraint on moose in south-central Alaska[J].Ecology,2009,90(5):1400 -1411.
[26] Bryant J P,Reichardt P B,Clausen T P.Chemically mediated interactions between woody plants and browsing mammals[J].Journal of Range Management,1992,45(1):18 -24.
[27] Schwartz C C,Nagy J G,Regelin W L.Juniper oil yield,terpenoid concentration and antimicrobial effects on deer[J].The Journal of Wildlife Management,1980,44(1):107 -113.
[28] Cooper S M,Owen-Smith N.Condensed tannins deter feeding by browsing ruminants in a South African savanna [J].Oecologia,1985,67(1):142-146.
[29] Palo R T.Chemical defense in birch:inhibition of digestibility in ruminants by phenolic extracts[J].Oecologia,1985,68(1):10-14.
[30] 程建国,罗燕,蔡永华,等.中药对麝香产量的影响研究[J].安徽农业科学,2009,37(26):12579-12581.
猜你喜欢
酿酒科技(2022年5期)2022-05-24
天津农林科技(2022年2期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
疯狂英语·新悦读(2021年10期)2021-11-23
文萃报·周二版(2021年5期)2021-03-02
中国酿造(2020年12期)2021-01-10
中国果菜(2019年11期)2019-12-14
金秋(2019年24期)2019-06-17
中老年健康(2016年1期)2016-03-07
军事体育学报(2014年4期)2014-02-27
