
两种生境下野慈姑繁殖差异及其机制
2018-06-23 03:08罗文杰金晓芳汪正祥
生态学报 2018年10期
罗文杰, 金晓芳, 汪正祥, 3, 4, 戴 璨, 3, 4,*
1 湖北大学资源环境学院,武汉 430062 2 南昌工程学院生态与环境科学研究所,南昌 330099 3 湖北省生物资源绿色转化协同创新中心,武汉 430062 4 区域开发与环境响应湖北省重点实验室,武汉 430062
植物与物理环境的互作是生态学关注的一大焦点。一方面,环境对植物的生长和繁殖会带来极大的直接影响[1- 2]。植物生长不可或缺的光照、温度、水分、营养在不同的地理尺度上必然存在变化,各个因素的异同往往会造成植物生长速率、发育水平、繁殖方式或生活史某种程度的改变。例如在光照充足的环境下玄参科的沟酸浆属植物(Mimulus)会较遮阴环境下推迟开花[3],类似的开花物候推迟也出现在高寒草甸植物群落对光照长度的响应中[4]。湿生扁蕾(Gentianopsispaludosa)在草甸生境获得的光照资源多于灌丛生境,导致其在蕾期的花朵数量显著偏高[5]。如果某种环境压力是持续不变的,从长远来看,还会导致植物种群的进化,即产生各种性状的适应性改变[6]。例如美洲风铃草(Campanulastrumamericanum)在林荫环境下多为两年生的植物,而光照环境下多为一年生[7]。斑点老鹳草(Geraniummaculatum)在异质性环境下种群的繁殖系统也存在着差异,干燥环境下主要是性二态植株,而湿润环境下主要是性单态植株[8]。
另一方面,物理环境的差异会导致生物群落的改变,继而使得与植物相关的各种互利共生、寄生、竞争、捕食等种间关系发生变化。其中,传粉者对有花植物的有性繁殖起着至关重要的作用,如果传粉者的组成和行为在不同的环境下发生改变,势必会给繁殖带来影响。Herrera对宽叶薰衣草(Lavandulalatifolia)的6年调查表明:不同生境下该植物出现了分别以蜂类和蝶类为主要传粉者的居群,由于不同昆虫的本身属性和行为差异,蜂类访花频繁但提高了同株异花授粉(geitonogamy),蝶类访花较少但因飞行距离远从而促进了异株传粉[9]。
过去的研究往往强调不同的环境因子和植物相应的繁殖表现,即整个过程的起点和终点,而未细致拆分每一个可能发生变化的阶段,也未区分生物因子和非生物因子或独立或协同的作用[7]。例如,调查发现相思树(Acacialongifolia)在湿润气候下果实败育数量要显著少于干旱气候的[1];南北坡向对暗紫贝母(Fritillariaunibracteata)的单果种子数存在显著影响[10];川滇蔷薇(Rosesoulieana)的果实重量和种子活力在山区河谷的不同流域间差异显著[11]等等。以上研究极易导致某种物理环境被贴上或“恶劣”或“优异”的标签,然而这些植物在座果和结实上的变化还可能是受到群落或传粉的间接影响,如果不进行细致研究,很可能导致错误的归因和建议。
也有少量研究同时关注了物理环境和传粉者对植物的影响。例如,美洲风铃草(Campanulastrumamericanum)在林隙和林下两种生境里的开花物候、昆虫访花行为以及花粉限制程度存在显著差异,导致了前者的果实数量更高,进一步的分析得出其繁殖差异并不是由某一个因素独立决定,而是多个直接和间接影响同向叠加的结果[7],类似的原因也导致了桃叶风铃草(Campanulapersicifolia)的个体繁殖差异[12]。也有研究发现环境对植物的直接和间接作用方向相反,如不同海拔的两个草甸毛茛(Ranunculusacris)居群里,低海拔植株的开花和胚珠数量均高于高海拔的个体,但是低海拔的居群存在明显的花粉限制,导致其繁殖不足[13]。以上实例表明,环境介导的传粉作用不可忽视,这一环节的缺失会影响理解不同生境下植物差异性繁殖的根本原因。
本文通过控制对比实验,研究两种不同生境,特别是光照环境对于植物繁殖适合度的影响,强调同时关注环境的直接作用和传粉者的间接作用,以期分析环境导致植物繁殖差异的机制。以多年生草本植株野慈姑(SagittariatrifoliaL.)作为研究材料拟解决以下问题:(1)不同的光照环境是否影响野慈姑的开花生理?(2)光照环境的差异是否影响野慈姑传粉昆虫的组成和访花行为?(3)不同光照环境下野慈姑繁殖产出如何改变?(4)环境对植物的直接与间接作用的方向是否一致?相对大小如何?
1 材料与方法
1.1 实验材料
野慈姑(SagittariatrifoliaL.)隶属泽泻科慈姑属,是一种生长在池塘、沼泽、沟渠、水田等浅水区域的多年生单子叶草本植物[14],适应能力较强,几乎全国各地均有分布[15],且自然生长于各种光照环境下。它是一种自交亲和、兼有有性繁殖(开花结实)和无性繁殖(地下球茎)的植物。花期5—10月左右(湖北地区的盛花期7—8月),总状花序,每轮一般三朵花,自下而上顺次开放,花序基部为雌花,开放持续1—2 d,顶部为雄花,开放持续4—10 d。野慈姑为典型的雌花先熟、雌雄异花同株的植物,且单花开花时长仅有一天,传粉者在6:00—14:00间表现活跃[16]。野慈姑的雄雌花的花部结构基本相同,都有三瓣白色花瓣,具花蜜,雌花的中部为心皮,呈球形、绿色,雌花在授粉三周后长为成熟的球形瘦果;雄花的中部为花药,多雄蕊、呈黄色。野慈姑是虫媒传粉的植物,食蚜蝇、小型蜂类等多种昆虫可以为其传粉[17]。
1.2 实验设计
1.2.1 质区选取方法
实验于2015年在中国科学院武汉植物园开展,在野外选取两个不同环境的实验区:一个区域上方有高大的池杉和樟树,周边大部分区域为天然乔木灌木群落,为典型的密闭林下环境(以下简称林荫区);另一个区域选在一定高度的露天台地,周围没有任何高大植株或建筑物的遮蔽,在晴朗的天气太阳可整日连续照射,为典型的全日照环境(以下简称日照区)。两个实验区均有88盆野慈姑的盆栽植株。所有植株来自于2013年野外采集的球茎克隆分株[18],且两个区域的基因型组成基本相同,以确保环境是野慈姑后续开花传粉和繁殖差异的单一来源。
1.2.2 光照强度测量
因为光照因子是质区选择的主要考虑,为了量化两个区域的光照强度差异,在林荫区和日照区分别放置一个相同型号的光量子计(Apogee MQ~200,美国犹他州),于2015年7月24日开始对光合有效辐射(PAR,以下简称光强,μmol m-2s-1)进行连续30 d的测量。为了全面反映每个区域的平均光照强度,光量子计每天移至区域内一个新的随机位置。记录从6:00持续至19:30,每30 min自动计算并存储该时间段内的平均光照强度,故每天林荫区和日照区分别有28个数据。
1.2.3 野慈姑开花统计
于2015年7月24日至30日,每天8:00—10:30对当日两个区域内开放的野慈姑花朵进行统计,包括每个区内野慈姑的雄花数量、雌花数量、总开花数量和开花植株数量。同时对当日开放的雌花用吊牌标记,便于收获和统计果实。
1.2.4 昆虫访花观察
在上述开花统计时段内,选择晴朗或多云天气,于每日6:00—14:00间观察访花昆虫对实验区域内野慈姑花朵的访问。一组实验以花朵为观察对象,记录固定花朵被昆虫访问的情况。对林荫区和日照区分别选取位置集中的共计约40朵野慈姑花(雌雄花各半)作为观察对象,进行连续15分钟的观察,记录传粉者访问雌花和雄花的总次数,传粉者在花朵上停留时间超过3秒记录为成功访花一次。在6:00—14:00的时间段内每个小时每个区域完成4个15分钟的观察记录(共计32个),最后计算昆虫访花频率(昆虫访问的总次数÷观测的花朵数,单位:次/(朵×15 min))。
另一组实验以单个访花昆虫的一个访问回合为单位(从昆虫进入到飞离区域记为一个完整的访问回合,Foraging bout),记录昆虫类别和详细的访问过程。同时,对每个区域内出现的访花昆虫进行拍照和取样,用于物种鉴定。观测的指标包括每个访问回合访花昆虫的种类(蜂类、蝇类、蝶类、蚜蝇类和其他五大类),每回合访花持续时间,每回合访问的花序数量、雄花数、雌花数和总花数,同时计算每回合的单花停留时间(访花持续时间÷总花数)。每个区域分别观察记录约50个访花回合。
1.2.5 繁殖产出
野慈姑的繁殖产出为在开花统计时段内所有开花植株的座果率、单果种子数量和种子平均面积。在两个区域内野慈姑雌花开放约20 d后,将所有成熟果实采摘,单个存放于牛皮纸袋中(6 cm×10 cm),烘干待用(50℃,12 h)。同时,分别统计林荫区和日照区每个开花植株的果实数量和座果率(果实数量÷雌花数量)。对每个植株随机挑选3个果实进行种子计数。具体方法为将每一个果实的所有种子人工平铺于一张A3白板纸上,尽量保证种子之间没有重合,用固定位置和焦距的单反照相机(尼康D7000)对其拍照,然后用Image J 软件对照片进行分析处理,得出单果种子数量与种子的平均面积。
1.3 数据统计与分析
用SAS数据处理软件利用方差分析(ANOVA, proc glm, proc mixed)对两个区域的光合有效辐射、开花情况、昆虫访花频率和昆虫每回合的访花情况进行了比较。在光强的分析中,把“区域”作为一个固定因素,“日期”作为一个随机影响因素,“时间”(记为从1到28的连续变量)及其二次项和它们分别与区域的交互效应都同时列入固定的变异来源(且结果均显著)。在开花情况的统计中,区域为固定因素,日期为随机因素。比较两个区域的昆虫访花频率时,把花性别、区域和时间(同上)作为固定因素,天气(晴朗和多云两类)和观察者作为随机因素(交互作用不显著)。访花昆虫的组成在两个区域的区别采用卡方分析(Chi-Square, proc freq)。昆虫每回合访花的分析中,响应变量有访花持续时间、单花停留时间、访问雄花数、访问雌花数、访问总花数和访问花序数,每个模型中,区域、昆虫种类(由于其他类昆虫组成十分多样、不具有代表性,故只计入4种类型)和每个回合对应的时间(同上)都同时作为固定因素,日期作为随机因素(交互作用不显著),并采用Tukey进行事后两两比较。针对花朵和花序数量的4个变量,还在模型中分别加入了总雄花数、总雌花数、总花数和总开花花序数作为协变量,以分析昆虫的每个访花回合是否受整个区域开花情况的影响。繁殖产出方面,两个区域座果率的比较采用响应变量为二元的广义线性混合模型(proc glimmix, dist=binary);种子数量和面积的比较采用了方差分析。两个模型中固定因素是“区域”,随机因素是“个体”。为了满足方差分析的正态性和方差同质性的前提,对种子面积进行了平方根转换。
2 结果
2.1 两种环境的光强与野慈姑的开花情况
日照区与林荫区的光合有效辐射有显著差别,日照区的日平均光照强度为(751.6 ± 21.4) μmol m-2s-1,而林荫区的为(248.3 ± 11.7) μmol m-2s-1,日照区显著偏高(F1,1890=13.67,P=0.0002)。两个区域内野慈姑的开花情况统计发现:日照区每日开放的雌花数、雄花数、开花总数量和开花的植株数都显著大于林荫区(表1),表明光照充足的生长环境对植物的有性繁殖存在正向增益。
2.2 两种环境访花昆虫的访花频率
日照区昆虫的访花频率要高于林荫区(图1),该差异接近显著(F1,122=3.53,P=0.06),说明光照强的环境在一定程度上促进了昆虫对野慈姑单个花朵单位时间的访问。同时,昆虫访花的时间段也对访花频率有显著的影响(F1,122=6.58,P=0.01),表现为从上午到下午的递增趋势。访花频率在雌雄花之间不存在性别差异(F1,122=0.07,P=0.79)

表1 两种生境下野慈姑的开花情况
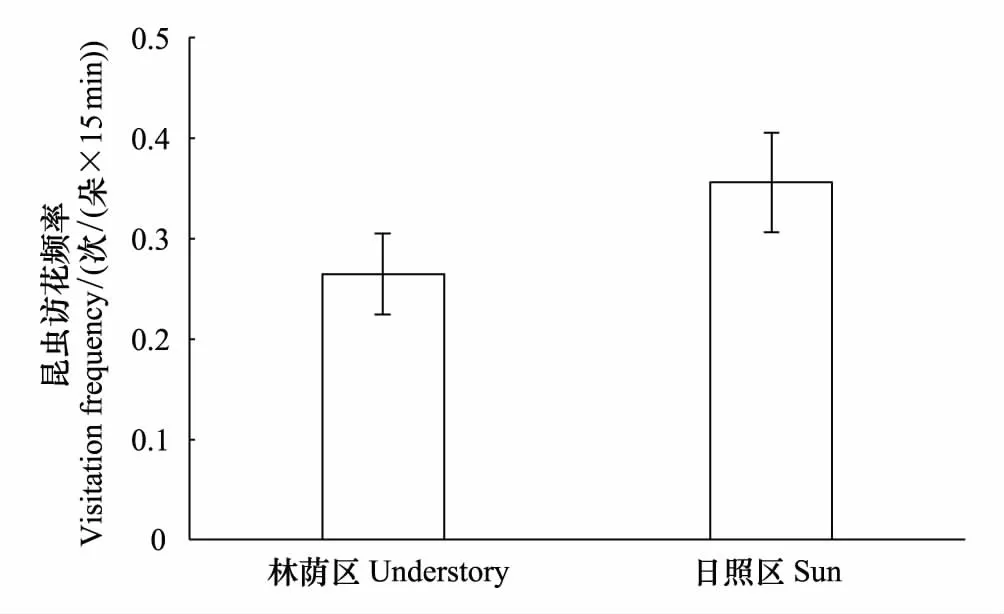
图1 两种生境下野慈姑的昆虫访花频率 Fig.1 The pollinator visitation frequency on Sagittaria trifolia under two habitats
2.3 两种环境的访花昆虫种类
野慈姑的访花昆虫可大致分为五类:蚜蝇类、蝇类、蜂类、蝶类以及其他类(表2),其他类的昆虫从物种构成上非常多样,但每种来访的次数均较少。林荫区记录共计49个次,其中有蚜蝇类23个次,以黑带(细扁)食蚜蝇为主,兼有少量其他种(拉丁名见表2,下同),常见的还有蝇类(丝光绿蝇)11个次,蜂类(意大利蜂)5个次,和蝶类(灰蝶科)5个次;其他类昆虫有记录5个次(日本弓背蚁、二星蝽属、小长蝽属、蟹蛛科等)。日照区记录共计51个次,其中蜂类13个次,蚜蝇类(黄斑粗腿管食蚜蝇、棕腿斑眼蚜蝇、黑色斑眼蚜蝇、黄道蚜蝇、裸芒宽盾蚜蝇)9个次,蝇类(丝光绿蝇、口鼻蝇属、纹蝇属)7个次和蝶类(曲纹稻弄蝶、点玄灰蝶、酢浆灰蝶、粉蝶科)4个次;其他类包括黑异喙蜾赢、墨胸胡蜂、七星瓢虫、叶舌蜂属、隧蜂科、指角蝇科等18个次。除开其他类,四种不同访花者类型的占比在两个区域有明显区别(χ2=9.3,P=0.026; 图2),林荫区的传粉昆虫以蚜蝇为主,而日照区的为蜂类。总体而言,野慈姑的花朵能够吸引多种昆虫访花,并不依靠某单一类群,属于泛化的访花系统。

表2 野慈姑访花昆虫种类名录
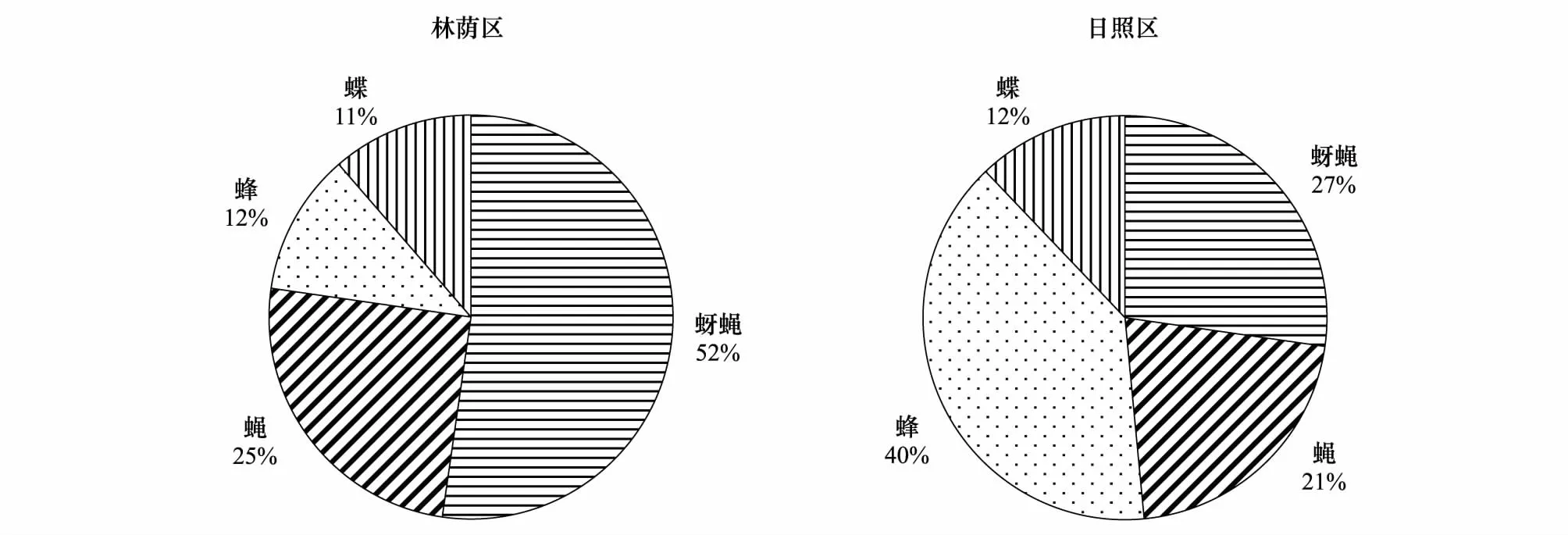
图2 两种生境下野慈姑的访花昆虫组成Fig.2 Pollinator compositions of Sagittaria trifolia under two habitats
2.4 两种环境下访花昆虫每回合访花情况
日照区的昆虫每个回合访问的花序数量高于林荫区、且访问的总花数和雄花数也显著高于林荫区,但是访问的雌花数量相当(表3)。值得一提的是,整个区域的开花情况对各个访问量均没有线性影响(开放的总花序数:F1,64=2.23,P=0.14;总花数:F1,64=0.24,P=0.63;总雄花数:F1,64=0.07,P=0.80;总雌花数:F1,64=0.05,P=0.83)。这表明昆虫访花行为上的区别在很大程度上是直接受环境影响,而不是由两个区域在开花基础数量上的差异所导致。虽然每个访花回合的持续时间在两个区域之间差异不明显,但是林荫区的单花停留时间有高于日照区的趋势(表3)。
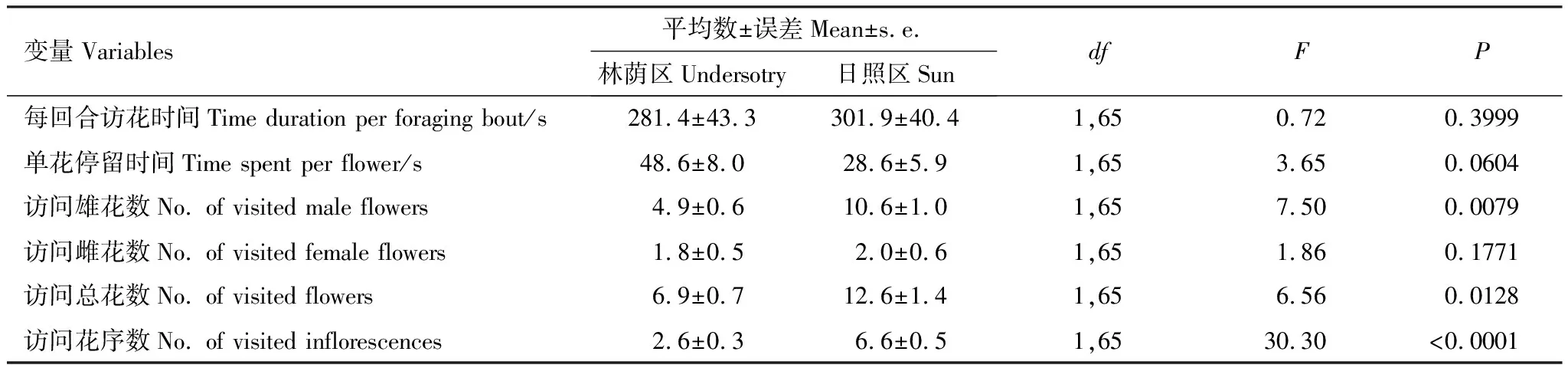
表3 两种生境下野慈姑传粉昆虫的每回合访花特征
从昆虫类型来看,蝇类在每个访花回合的持续时间上显著长于蜂类(t= 2.73,P=0.04);且蝇类访问的雌花数显著高于其他3个类型(F3,65=5.50,P=0.002)。以上结果说明蝇类访花习性明显与其他的访花昆虫不同,表现出偏雌花的喜好和长时间的停留(图3)。
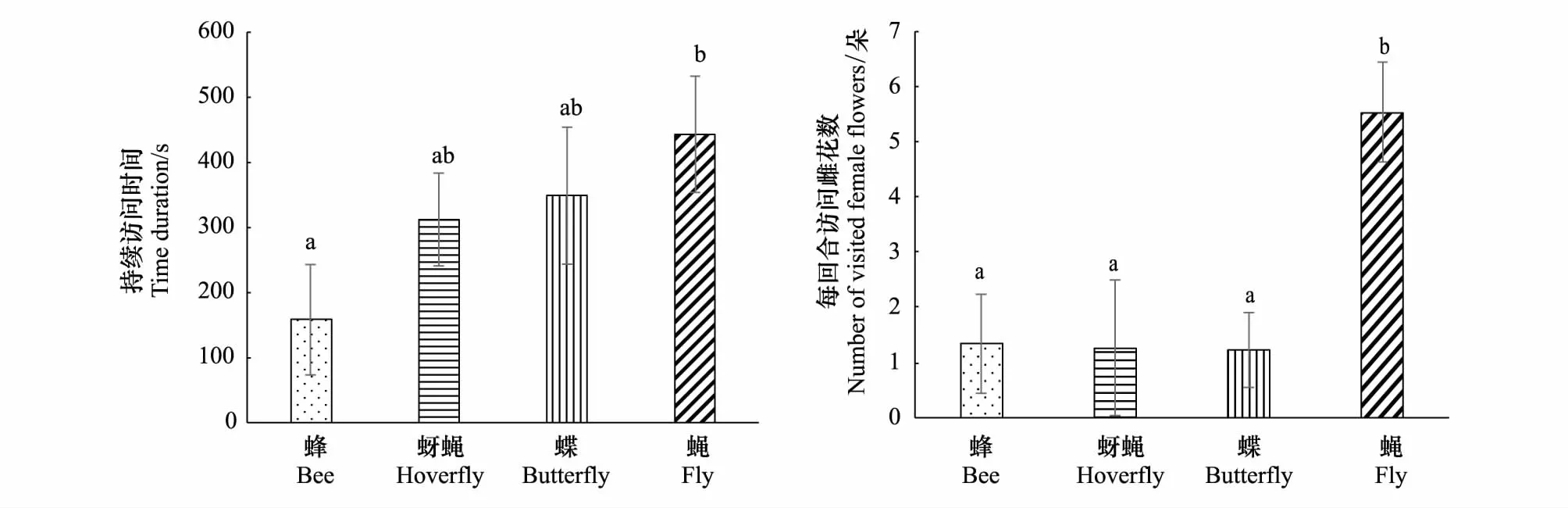
图3 野慈姑不同类型的传粉昆虫每访花回合的访问特征Fig.3 Pollinators′ visitation characters per foraging bout on Sagittaria trifolia for different pollinator categories
2.5 两种环境下野慈姑的繁殖产出
在实验时间段内,日照区结实总量为322个,而林荫区为99个。日照区的座果率显著高于林荫区(F1,460=17.1,P<0.0001)。单果水平上,林荫区的种子数量与日照区的基本一致(F1,164=0.16,P=0.69),暗示两个区域的雌花在胚珠数量上没有显著的不同,即光照环境未影响植株在胚珠上的繁殖投入。同时,林荫区的种子平均面积与日照区的也无显著差异(F1,164=2.16,P=0.14),表明两个区域的种子质量区别不明显(图4)。

图4 两种生境下野慈姑的繁殖产出Fig.4 The reproductive output of Sagittaria trifolia under two habitats***表示P<0.001,ns表示两者之间区别不显著
3 讨论
3.1 两个生境的光照差异及其对野慈姑的直接影响
光是植物生长必须的能量来源,本实验在选取野慈姑的两个不同生境时最主要的考量因子是光照,一为没有任何遮盖的日照区,另一为林荫区,实验证实两个区域的光合有效辐射存在显著差异,且日照区的日平均光强是林荫区的3倍左右。除光照外,由于两个实验区域的直线距离仅约800 m,其天气、降水等气象因素应该相差无几。盆栽的实验设计也决定了两个区域中土壤和肥力的一致性。虽然气温理应相当,但微环境的温度通常会随着光强的增大而提升[7]。故本研究的两个生境应该是在光强和温度上均有差异,表现为日照区的光强和温度都高于林荫区,均更利于植物的生长和有机物的积累[19]。
从开花阶段的表现来看,日照区野慈姑的每日平均开放雄花数、雌花数、总花数和开花总花序数均为林荫区的2—3倍。由于两个区域野慈姑的基因型构成相同(通过球茎克隆分株种植得来),其在开花数量上的差异体现出环境的主导作用,表明日照区野慈姑的资源总量和有性繁殖投入远远大于林荫区。值得强调的是,此处开花数量在两个区域之间的差异是物理环境引起的直接响应,还未涉及到传粉和结实阶段的变化。由于本研究只关注了花水平的数量差异,对于更小层次的胚珠或花粉的变化我们不得而知,不过从单果平均种子量和种子面积在两个区域的无差异性推测,林荫区应该与日照区相当,间接证明了生境差异并未导致胚珠数量或质量上的显著下降。这方面其他植物也有类似的报道,例如心叶凹唇姜(Boesenbergialongiflora)的单花胚珠数在三个不同的生境里并不存在显著差异[20],可能与植物开花固有的发育机理相关。
3.2 环境对野慈姑的间接影响——传粉
在两个生境中,野慈姑访花昆虫的组成、昆虫类型的相对重要性及其行为有明显的变化。日照区的访花昆虫较林荫区更为多样,这可能是受日照区周边植物群落的影响,该区域生长了很多具蜜腺的有花植物,如葫芦科、苋科和菊科植物(作者观察),吸引了不少种类和数量的传粉昆虫。相对而言,林荫区的“其他类”昆虫较少,可能是林荫生境下的群落组成多为阴生植物,如禾本科、三白草科和部分蕨类植物等(作者观察),而这样的群落本身对传粉昆虫需求不大,故群落整体的吸引力不强。需要指出的是,蝶类在访问野慈姑时均只在花冠停留用较长的口器采蜜,其携带花粉的能力较弱(作者观察),所以两个生境中的传粉昆虫应只包括蜂、蝇、蚜蝇三类。
林荫区野慈姑最主要的传粉昆虫是食蚜蝇,而日照区为蜜蜂,这也许是上述不同植物群落的效应,还可能跟昆虫本身的习性相关,如意大利蜜蜂(本研究的传粉者)的某些品种对高温高热环境的抗逆性很强[21]。不同生境里传粉昆虫的重要性差异基于野慈姑的泛化传粉系统(generalist),在同类研究中也有一些特化传粉的植物(specialist)可以同时控制光照和传粉昆虫类型[7],故其变量相对单一、解释也更为明确。本实验中蜂类的访花在停留时间上显著偏短,表现为“短平快”的访花习性,有其他研究指出其相较于蚜蝇类昆虫有更高效的花粉传递[21];且日照区的蜂类占比较多,这可能对该区域野慈姑的结实存在一定积极影响。此外,蝇类的访问行为较为特殊,它们倾向于访问更多的雌花且停留更长的时间,然而,长时间在单一性别的花上停留会导致花粉得不到高效的传递[22],也可能损害花朵的某些结构[23],对结实造成负面影响。鉴于蝇类在两个区域出现的几率相当(25%和21%),不应是区域间繁殖差异的主要原因。
由环境介导的野慈姑的传粉差异还表现在两个区域内传粉昆虫的访花频率和每回合访问花朵数量上的大相径庭。日照区的昆虫访花频率高于林荫区,单位时间内访花次数的增加可给花朵带来足量授粉从而促进结实。这也许是环境影响的昆虫行为和花展示特征共同造成,因为日照区的开花数量更大,对昆虫的吸引力更强,从而提升了单花的访花频率,这一结果与很多研究吻合[7, 12]。在每个回合的访问中,传粉昆虫在不同生境访问的雌花数量均为2个左右,而日照区的昆虫访问两倍于林荫区的花序数和雄花数,暗示昆虫在携带花粉数量和基因多样性上的倍增。由此推测,即便同是对雌花的单次访问,日照区的昆虫可能会传递更多高质量的花粉,进而提升了单次访问的效益。结果中已经指出,昆虫在访问回合中访花数量的差异不依赖于区域内的开花数量,即不论区域内开花多少,日照环境比林荫环境的昆虫在每个访花回合中倾向于访问更多的雄花和花序,因此,该影响应该是环境对昆虫访花行为独立作用的结果。
4 结论
总体而言,日照区的野慈姑由于其生境内光照的提升,开花数量更多,传粉昆虫对每朵花在单位时间内的访问次数更多,每次访问携带花粉的数量或多样性更高,最终导致座果率更高。环境介导的植株繁殖表现能够在同一方向上不断积累增加,对植物的适合度产生更大的增益,最终使得两个生境下基因型组成相同的野慈姑在繁殖总量上达到了3倍以上的差距。虽然日照区的野慈姑在繁殖过程中的每一步都领先于林荫区,但从作用强度上看,环境因子对花朵数量的直接影响占主导地位;而其间接影响,由环境介导的传粉昆虫的组成和行为的作用,对结实差异的贡献量相对较小。光照条件更好的环境下植物的繁殖产出往往更高[7, 24],这几乎是众所周知的现象;然而,本文从细节出发,量化每个繁殖阶段的表现,特别是传粉昆虫在不同生境下的响应,从机制上阐述了繁殖差异产生的原因。同时,本文也指出此类研究中同时关注植物、物理环境和生物因子三者之间互作的重要性,只有这样,才能从根本上理解环境的异质性对于植物生长和繁殖的影响。
致谢:感谢湖北大学梁熙健、廖明林同学对实验的帮助,苗璐、李乐娜对写作的帮助。
参考文献(References):
[1] Fernandes P, Antunes C, Correia O, Máguas C. Do climatic and habitat conditions affect the reproductive success of an invasive tree species? An assessment of the phenology ofAcacialongifoliain Portugal. Plant Ecology, 2015, 216(2): 343- 355.
[2] Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, van der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167- 234.
[3] Galloway L F, Burgess K S. Artificial selection on flowering time: influence on reproductive phenology across natural light environments. Journal of Ecology, 2012, 100(4): 852- 861.
[4] Keller F, Körner C. The role of photoperiodism in alpine plant development. Arctic, Antarctic, and Alpine Research, 2003, 35(3): 361- 368.
[5] 侯勤正, 叶广继, 马小兵, 苏雪, 张世虎, 孙坤. 青藏高原不同生境下湿生扁蕾(Gentianopsispaludosa)个体大小依赖的繁殖分配. 生态学报, 2016, 36(9): 2686- 2694.
[6] Munguía-Rosas M A, Parra-Tabla V, Ollerton J, Cervera J C. Environmental control of reproductive phenology and the effect of pollen supplementation on resource allocation in the cleistogamous weed,Ruellianudiflora(Acanthaceae). Annals of Botany, 2012, 109(2): 343- 350.
[7] Kilkenny F F, Galloway L F. Reproductive success in varying light environments: direct and indirect effects of light on plants and pollinators. Oecologia, 2008, 155(2): 247- 255.
[8] van Etten M L, Chang S M. Effects of environmental heterogeneity on the distribution of sexes within and among populations in a gynodioecious species,Geraniummaculatum. New Phytologist, 2009, 183(3): 649- 660.
[9] Herrera C M. Components of pollinator “quality”: comparative analysis of a diverse insect assemblage. Oikos, 1987, 50(1): 79- 90.
[10] 陈文年, 王辉, 肖小君, 陈发军, 张志勇, 齐泽民, 黄作喜. 坡向对暗紫贝母生长和繁殖特征的影响. 生态学报, 2016, 36(24): 8174- 8182.
[11] 周志琼, 包维楷. 横断山区干旱河谷川滇蔷薇生长与繁殖特征的地理空间差异. 生态学报, 2016, 36(8): 2192- 2201.
[12] Hansen V I, Totland Ø. Pollinator visitation, pollen limitation, and selection on flower size through female function in contrasting habitats within a population ofCampanulapersicifolia. Canadian Journal of Botany, 2006, 84(3): 412- 420.
[13] Totland Ø. Environment‐dependent pollen limitation and selection on floral traits in an alpine species. Ecology, 2001, 82(8): 2233- 2244.
[14] 陈家宽. 中国慈姑属的系统与进化植物学研究. 武汉: 武汉大学出版社, 1989: 31- 38.
[15] Chen J M, Liu F, Wang Q F, Motley T J. Phylogeography of a marsh herbSagittariatrifolia(Alismataceae) in China inferred from cpDNA atpB-rbcL intergenic spacers. Molecular Phylogenetics and Evolution, 2008, 48(1): 168- 175.
[16] 覃道凤, 李婷, 戴璨. 野慈姑花粉限制评估的影响因素. 应用生态学报, 2015, 26(12): 3865- 3870.
[17] Huang S Q, Tang L L, Sun J F, Lu Y. Pollinator response to female and male floral display in a monoecious species and its implications for the evolution of floral dimorphism. New Phytologist, 2006, 171(2): 417- 424.
[18] 李婷, 覃道凤, 戴璨. 利用SSR荧光标记对野慈姑异交率的估测. 植物科学学报, 2015, 33(4): 554- 563.
[19] Raven J A, Griffiths H. Photosynthesis in reproductive structures: costs and benefits. Journal of Experimental Botany, 2015, 66(7): 1699- 1705.
[20] 杨淑霞, 高江云. 心叶凹唇姜的开花格局和雄性先熟机制. 植物生态学报, 2009, 33(3): 449- 459.
[21] Abou-Shaara H F, Al-Ghamdi A A, Mohamed A A. Tolerance of two honey bee races to various temperature and relative humidity gradients. Environmental and Experimental Biology, 2012, 10(4): 133- 138.
[22] 肖宜安, 曾建军, 李晓红, 胡文海, 何平. 濒危植物长柄双花木自然种群结实的花粉和资源限制. 生态学报, 2006, 26(2): 496- 502.
[23] Wesselingh R A. Pollen limitation meets resource allocation: towards a comprehensive methodology. New Phytologist, 2007, 174(1): 26- 37.
[24] Atlan A, Hornoy B, Delerue F, Gonzalez M, Pierre J, Tarayre M. Phenotypic plasticity in reproductive traits of the perennial shrubUlexeuropaeusin response to shading: a multi-year monitoring of cultivated clones. PLoS One, 2015, 10(9): e0137500.
猜你喜欢
中国科学探险(2022年8期)2022-12-31
小学生优秀作文(高年级)(2022年4期)2022-04-25
疯狂英语·新悦读(2022年12期)2022-03-08
养生月刊(2021年12期)2021-12-09
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
诗潮(2017年12期)2018-01-08
作文成功之路(高考冲刺)(2017年9期)2017-09-23
新农业(2016年19期)2016-08-16
中成药(2016年8期)2016-05-17
语文教学与研究(2014年6期)2014-03-12
